- Врожденный и приобретенный иммунитет. Клеточные и гуморальные механизмы

Содержание
- 2. Определение врожденного иммунитета. Свойства врожденного иммунитета. Врожденный иммунитет – наследственно закрепленная система защиты многоклеточных организмов от
- 4. «Врожденное» распознавание: образы «чужого» Клетки врожденного иммунитета распознают своими рецепторами наиболее консервативные структуры бактериальной стенки, белки,
- 7. Toll – рецепторы (Toll R): история открытия В 1985 году при исследовании мутаций у мушки -
- 9. История открытия TLR 4 типа В 1997 году Руслан Меджитов и Чарльз Дженуэй из Йельского Университета
- 10. Руслан Меджитов - биография Первооткрыватель человеческих рецепторов врожденного иммунитета, выходец из Узбекистана Руслан Меджитов стал лауреатом
- 11. Мембранные рецепторы. - TLR (Toll-Like Receptor - Toll-подобный рецептор; т.е. сходный с Toll-рецептором дрозофилы) Мембранные рецепторы.
- 12. Примеры Toll - рецепторов врожденного иммунитета, распознающих «шаблоны чужого»
- 13. ПРОВЕДЕНИЕ СИГНАЛОВ С TOLL-ПОДОБНЫХ РЕЦЕПТОРОВ Все TLR используют одинаковую принципиальную схему передачи активационного сигнала в ядро.
- 14. Сигналлинг с TLR4.
- 15. Цитоплазматические рецепторы: NOD-рецепторы (NOD1 и NOD2) NOD-рецепторы (NOD1 и NOD2) находятся в цитозоле и состоят из
- 16. Цитоплазматические рецепторы: RIG-подобные рецепторы (RLR, RIG-Like Receptors) RIG-подобные рецепторы (RLR, RIG-Like Receptors): RIG-I (Retinoic acid-Inducible Gene-
- 17. Хемоаттракция Аттракция - привлечение. Хемоаттракция – привлечение эффекторных клеток в очаг воспаления химическими факторами – хемоаттратантами.
- 18. Хемотаксис – направленное движение клеток по градиенту хемоаттрактантов
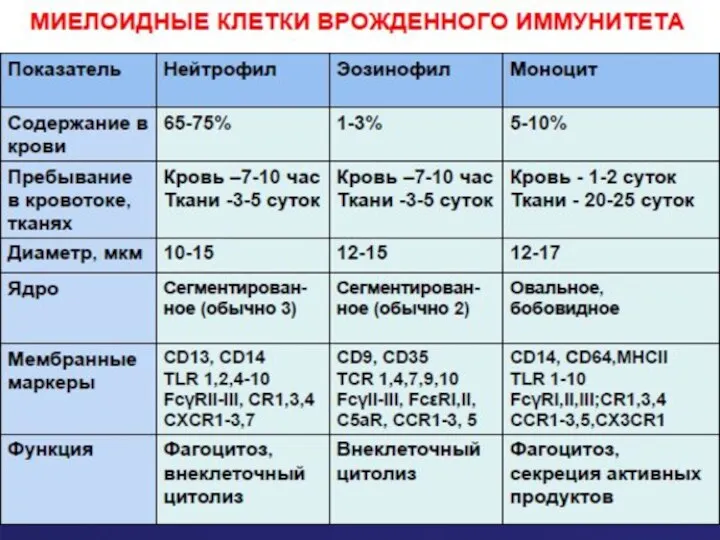
- 19. Нейтрофилы Нейтрофилы составляют значительную часть лейкоцитов периферической крови - 60-70%, или 2,5-7,5х10 в 9 степени клеток
- 21. Нейтрофил человека фагоцитирует Strep. pyogenes Нейтрофильные гранулоциты в большинстве своем выполняют в организме роль «мусорщиков» при
- 23. 1.Свободные радикалы кислорода в организме. 2.Образование активных форм кислорода фагоцитами («кислородный взрыв»: роль НАДФ-Н оксидазы, супероксиддисмутазы,
- 24. Основные процессы, в которых принимают участие нейтрофилы: миграция, поглощение, дегрануляция, внутриклеточный киллинг, деградация, экзоцитоз и апоптоз
- 25. Основные процессы, происходящие в нейтрофилах (НФ) при их активации и фагоцитозе
- 26. Эозинофилы Как и нейтрофил, эозинофил образуется в костном мозгу из клетки – предшественницы (миелоидный росток). В
- 28. Эозинофилы Эозинофилы чаще всего участвуют в борьбе с внеклеточными паразитарными инфекциями и аллергенами. Мембрана Цитоплазма Гранулы
- 29. Базофилы и тучные клетки(ТК) Тучные клетки при активации способны освобождать в окружающее пространство ферменты, цитокины, хемокины,
- 31. Унитарная теория (А. А. Максимов, 1909 г.) - все форменные элементы крови развиваются из единого предшественника
- 32. Макрофаги – потомки моноцитов крови Моноциты, макрофаги, нейтрофилы происходят от клеток –предшественников миелоидного ряда в костном
- 33. Гетерогенность клеток, происходящих от моноцитов. Тканевые макрофаги (МФ) и дендритные клетки (ДК) происходят от моноцитов (МН)
- 34. Макрофаг х 5000 Макрофаги во всех органах и тканях способны к : антигенной презентации (они являются
- 35. Дендритная клетка х 5000 Дендритные клетки- самые важные антигенпрезентирующие клетки (АПК), так как они расположены повсеместно;
- 38. Естественные киллеры (NK) –клетки врожденного иммунитета Большие гранулярные лимфоциты («нулевые клетки», «не Т, не В-клетки»). Т-
- 39. Естественные киллеры (NK) Число NK в крови - 5-15%; в лимфоузлах и селезенке 2.5-5% (от всех
- 40. Естественные киллеры (NK) Естественные киллеры играют ключевую роль в тех случаях, когда необходимо убить инфицированные или
- 41. Рецепторы естественных киллеров: для лизиса клетки-мишени недостаточно одной только отсутствия подавляющего сигнала, необходимо также наличие активирующего
- 42. Цитотоксическое действие естественных киллеров зависит от динамического равновесия между ингибиторными и активирующими рецепторами
- 43. Механизмы цитотоксичности NK-клеток 1. Большинство NK-клеток имеют своей поверхностный белок FasL, который при активации присоединяется к
- 45. Общая характеристика и свойства NK-клеток. Рецепторы для ИЛ-15 и ИЛ-21 ( IL15R и IL21R )
- 46. Гуморальные факторы врожденного иммунитета - дефенсины Дефензины (от англ. Defens защита) - катионные пептиды, активные в
- 47. Гуморальные факторы врожденного иммунитета – лизоцим Лизоцим (мурамидаза) — фермент, обладающий специфической способностью вызывать растворение некоторых
- 48. Гуморальные факторы врожденного иммунитета – белки острой фазы Осторофазовые белки – большая группа белков сыворотки крови
- 50. Пентраксины: СРБ С-реактивный белок (СРБ) был впервые описан в 1930 г. как белок, который в присутствии
- 51. Переход СРБ из пентамерной формы в мономерную приводит активации эндотелия, к продукции цитокинов, к адгезии нейтрофилов,
- 52. Защитные свойства СРБ СРБ имеет много свойств, характерных для иммуноглобулинов: он связывается с бактериальными полисахаридами и
- 53. СРБ, связывая фосфорилхолин клеточных стенок ряда бактерий и одноклеточных грибов, опсонизирует их и активирует систему комплемента
- 55. Гуморальные факторы врожденного иммунитета: цитокины Цитокины — составная часть молекулярных механизмов как врожденного, так и адаптивного
- 56. Цитокины макрофагов и дендритных клеток
- 57. Цитокины макрофагов и дендритных клеток
- 58. Гуморальные факторы врожденного иммунитета - интерфероны 1 типа –противовирусная активность Клетки врожденного иммунитета, пораженные вирусом, синтезируют
- 59. Роль цитокинов дендритных клеток (ДК) в запуске адаптивного иммунного ответа (этот материал будет рассмотрен подробнее далее)
- 60. СИСТЕМА КОМПЛЕМЕНТА –основная система гуморальных факторов врожденного иммунитета Термин «комплемент» ввёл Пауль Эрлих в конце 1890-х
- 61. Классический путь активации СК Последовательность открытия компонентов системы комплемента не соответствует очередности их вступления в реакцию
- 62. Активация комплемента по классическому пути Активацию комплемента по классическому пути запускает иммунный комплекс: комплекс антигена с
- 63. Каскад активации системы комплемента по классическому пути а. Сначала активируется фракция С1: она собирается из трех
- 64. МАС –мембраноатакующий комплекс з. С5-конвертаза расщепляет фракцию С5. и. Появившаяся в результате этого активная фракция С5b
- 65. Поверхность клетки – мишени, атакованная МАС
- 66. Пути активации системы комплемента Классический путь- филогенетически более новый (так как появился только у животных с
- 70. Врожденный и приобретенный иммунитет: основные различия Врожденный 1. Существует в любом организме до начала любой агрессии.
- 71. Врожденный и приобретенный иммунитет: основные различия Врожденный 3. Все клетки врожденного иммунитета активируются и реагируют на
- 72. Врожденный и приобретенный иммунитет: основные различия Врожденный 4. Реакции врожденного иммунитета разворачиваются почти мгновенно (и клетки
- 73. Врожденный и приобретенный иммунитет: основные различия Врожденный 5. Во врожденном иммунитете иммунологическая память отсутствует. В настоящее
- 74. Врожденный и приобретенный иммунитет: основные различия Врожденный 6. Врожденный иммунитет представлен у всех видов организмов; он
- 75. Дженуэй Чарльз А. – младший: постулаты Чарльз А. Дженуэй-младший (1943-2003), автор фундаментальных концепций, снискавших ему славу
- 77. Дженуэй Чарльз А. – младший: постулаты 1. Врожденный иммунитет защищает все виды живых существ ; адаптивный
- 78. Дженуэй Чарльз А. – младший: постулаты 3. Рецепторов, распознающих «чужое» во врожденном иммунитете намного меньше, чем
- 79. Дженуэй Чарльз А. – младший: постулаты 5 . Для запуска эффективного иммунного ответа вместе с антигеном
- 80. Дженуэй Чарльз А. – младший: постулаты 7. Образраспознающие рецепторы клеток врожденного иммунитета узнают образы «чужого» –
- 81. Дженуэй Чарльз А. – младший: постулаты 8. Сигнал 2 (костимуляция) запускает ответ адаптивного иммунитета. 9. Таким
- 82. Вопросы занятия №2 Определение врожденного иммунитета. Свойства врожденного иммунитета. Клетки врожденного иммунитета. Рецепторы клеток врожденного иммунитета.
- 83. Тестовые задания к занятию №2 11. К клеточным факторам врожденного иммунитета не относятся: Т лимфоциты Натуральные
- 84. Тестовые задания к занятию №2 3. Распознавание во врожденном иммунитете: грубое «образное» «по отсутствию своего» тонкое
- 85. Тестовые задания к занятию №2 5. Во врожденном иммунитете: после окончания иммунного ответа остаются клетки памяти
- 86. Тестовые задания к занятию № 2 7.К фагоцитирующим клеткам не относятся: Макрофаги Дендритные клетки Нейтрофилы Моноциты
- 88. Скачать презентацию

Определение врожденного иммунитета.
Свойства врожденного иммунитета.
Врожденный иммунитет – наследственно закрепленная система защиты
Определение врожденного иммунитета.
Свойства врожденного иммунитета.
Врожденный иммунитет – наследственно закрепленная система защиты

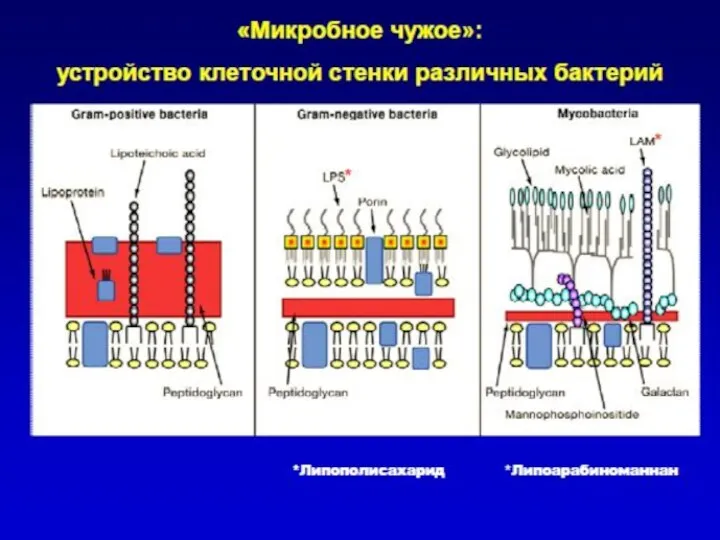
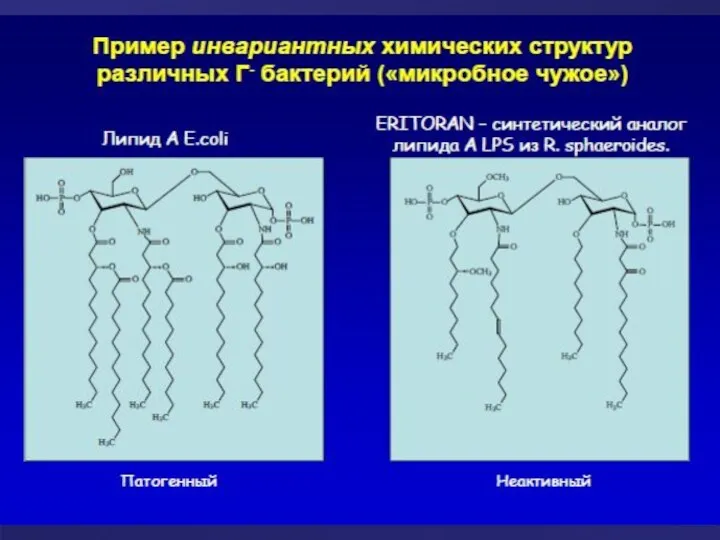
«Врожденное» распознавание: образы «чужого»
Клетки врожденного иммунитета распознают своими рецепторами наиболее консервативные
«Врожденное» распознавание: образы «чужого»
Клетки врожденного иммунитета распознают своими рецепторами наиболее консервативные

Toll – рецепторы (Toll R): история открытия
В 1985 году при исследовании
Toll – рецепторы (Toll R): история открытия
В 1985 году при исследовании

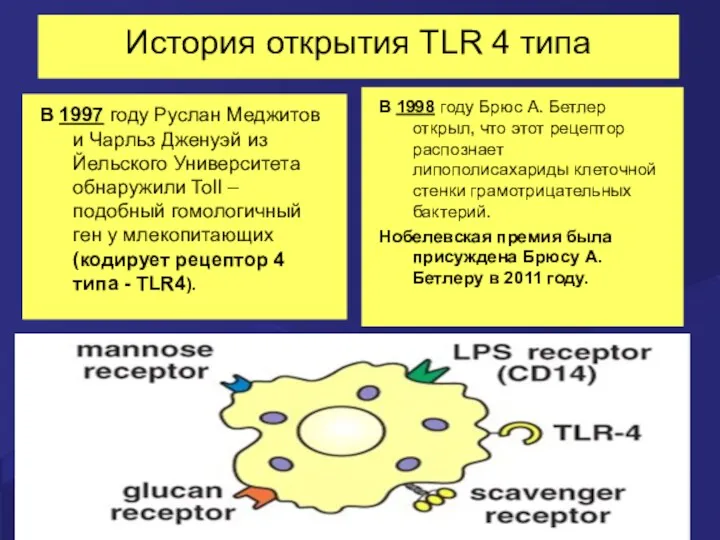
История открытия TLR 4 типа
В 1997 году Руслан Меджитов и Чарльз
История открытия TLR 4 типа
В 1997 году Руслан Меджитов и Чарльз

Руслан Меджитов - биография
Первооткрыватель человеческих рецепторов врожденного иммунитета, выходец из
Руслан Меджитов - биография
Первооткрыватель человеческих рецепторов врожденного иммунитета, выходец из
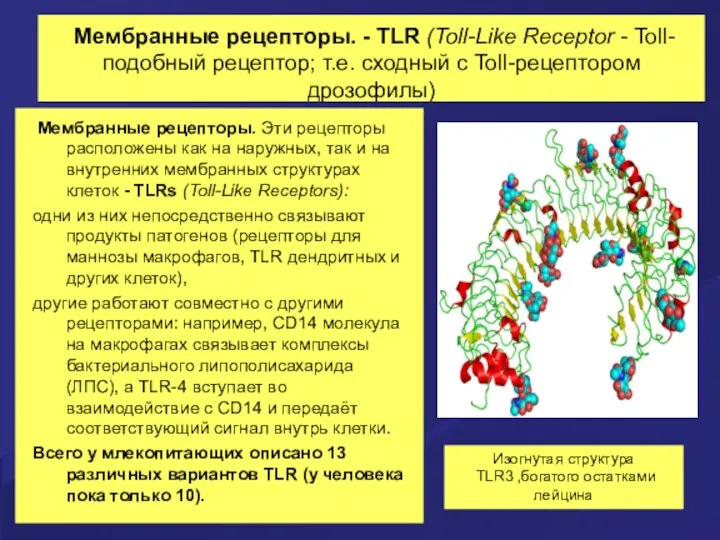
Мембранные рецепторы. - TLR (Toll-Like Receptor - Toll-подобный рецептор; т.е. сходный с Toll-рецептором дрозофилы)
Мембранные
Мембранные рецепторы. - TLR (Toll-Like Receptor - Toll-подобный рецептор; т.е. сходный с Toll-рецептором дрозофилы)
Мембранные
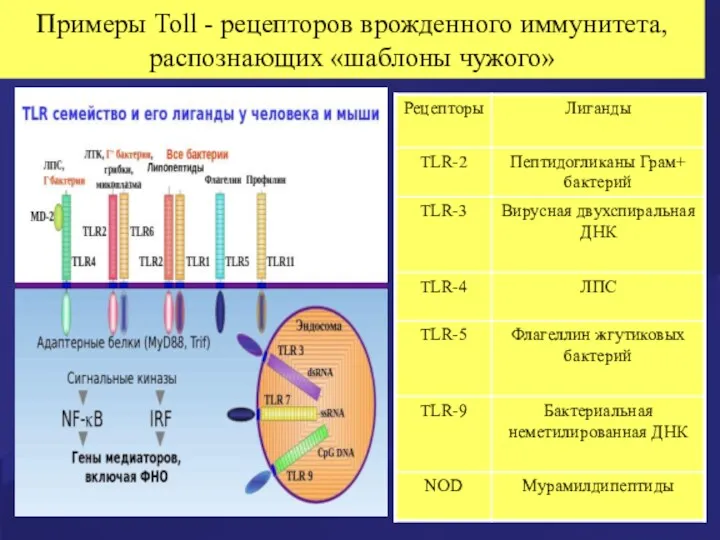
Примеры Toll - рецепторов врожденного иммунитета, распознающих «шаблоны чужого»
Примеры Toll - рецепторов врожденного иммунитета, распознающих «шаблоны чужого»

ПРОВЕДЕНИЕ СИГНАЛОВ С TOLL-ПОДОБНЫХ РЕЦЕПТОРОВ
Все TLR используют одинаковую принципиальную схему передачи
ПРОВЕДЕНИЕ СИГНАЛОВ С TOLL-ПОДОБНЫХ РЕЦЕПТОРОВ
Все TLR используют одинаковую принципиальную схему передачи
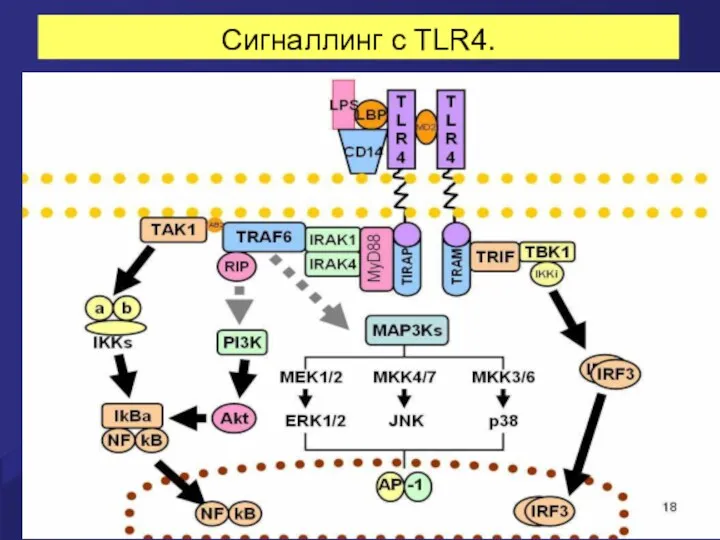
Сигналлинг с TLR4.
Сигналлинг с TLR4.

Цитоплазматические рецепторы: NOD-рецепторы (NOD1 и NOD2)
NOD-рецепторы (NOD1 и NOD2) находятся в цитозоле и
Цитоплазматические рецепторы: NOD-рецепторы (NOD1 и NOD2)
NOD-рецепторы (NOD1 и NOD2) находятся в цитозоле и

Цитоплазматические рецепторы: RIG-подобные рецепторы (RLR, RIG-Like Receptors)
RIG-подобные рецепторы (RLR, RIG-Like Receptors):
RIG-I (Retinoic acid-Inducible Gene- I)
MDA5 (Melanoma
Цитоплазматические рецепторы: RIG-подобные рецепторы (RLR, RIG-Like Receptors)
RIG-подобные рецепторы (RLR, RIG-Like Receptors):
RIG-I (Retinoic acid-Inducible Gene- I)
MDA5 (Melanoma

Хемоаттракция
Аттракция - привлечение. Хемоаттракция – привлечение эффекторных клеток в очаг
Хемоаттракция
Аттракция - привлечение. Хемоаттракция – привлечение эффекторных клеток в очаг
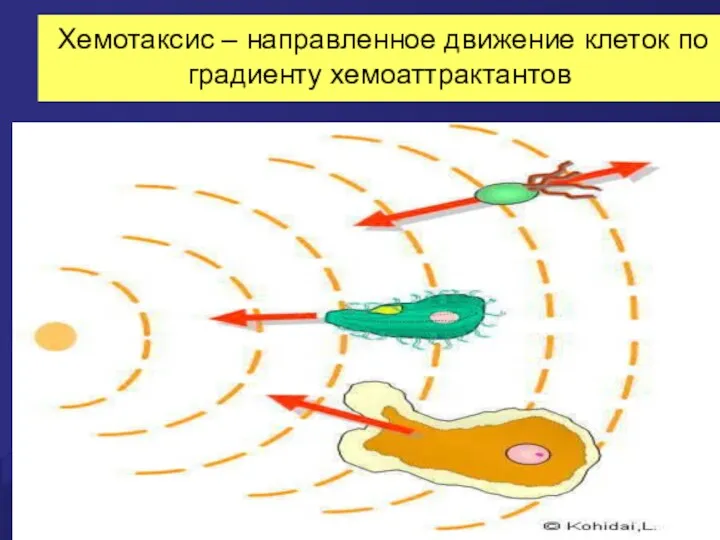
Хемотаксис – направленное движение клеток по градиенту хемоаттрактантов
Хемотаксис – направленное движение клеток по градиенту хемоаттрактантов
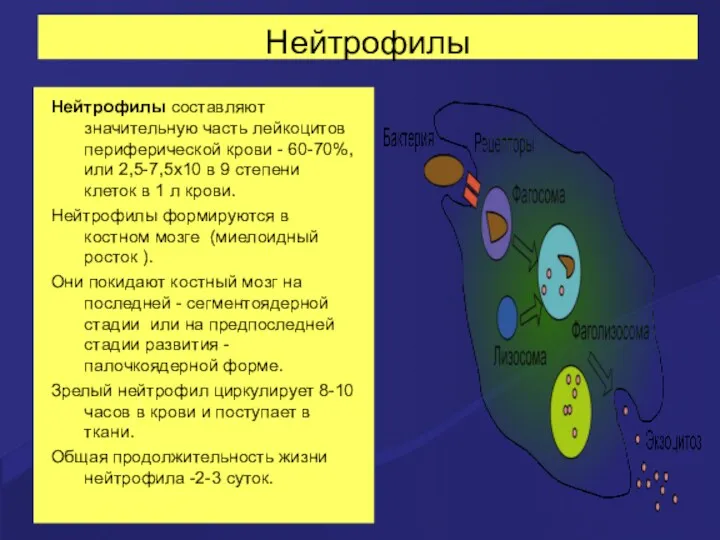
Нейтрофилы
Нейтрофилы составляют значительную часть лейкоцитов периферической крови - 60-70%, или 2,5-7,5х10 в
Нейтрофилы
Нейтрофилы составляют значительную часть лейкоцитов периферической крови - 60-70%, или 2,5-7,5х10 в
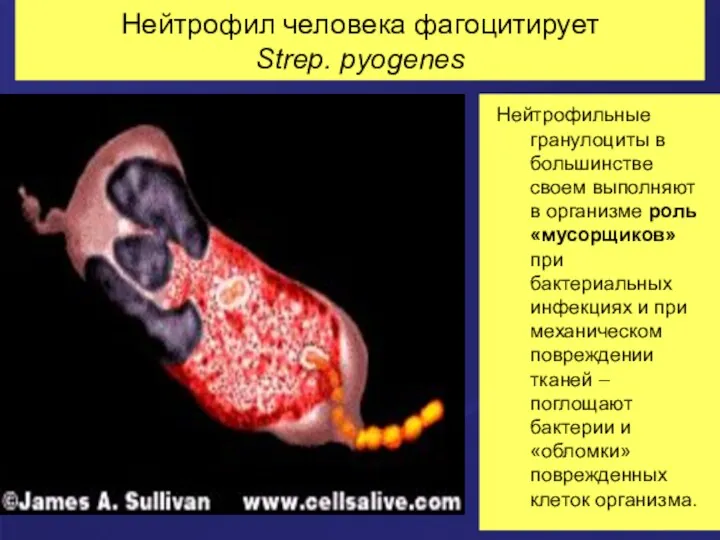
Нейтрофил человека фагоцитирует
Strep. pyogenes
Нейтрофильные гранулоциты в большинстве своем выполняют в
Нейтрофил человека фагоцитирует
Strep. pyogenes
Нейтрофильные гранулоциты в большинстве своем выполняют в
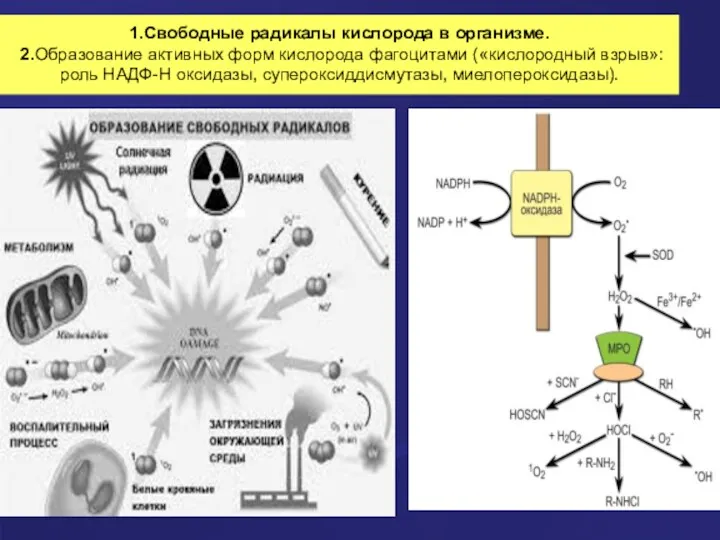
1.Свободные радикалы кислорода в организме.
2.Образование активных форм кислорода фагоцитами («кислородный взрыв»:
1.Свободные радикалы кислорода в организме. 2.Образование активных форм кислорода фагоцитами («кислородный взрыв»:

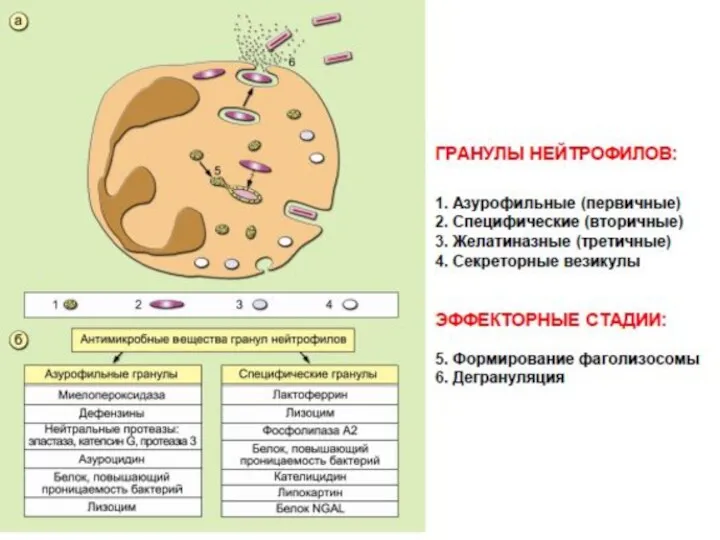
Основные процессы, в которых принимают участие нейтрофилы: миграция, поглощение, дегрануляция, внутриклеточный
Основные процессы, в которых принимают участие нейтрофилы: миграция, поглощение, дегрануляция, внутриклеточный
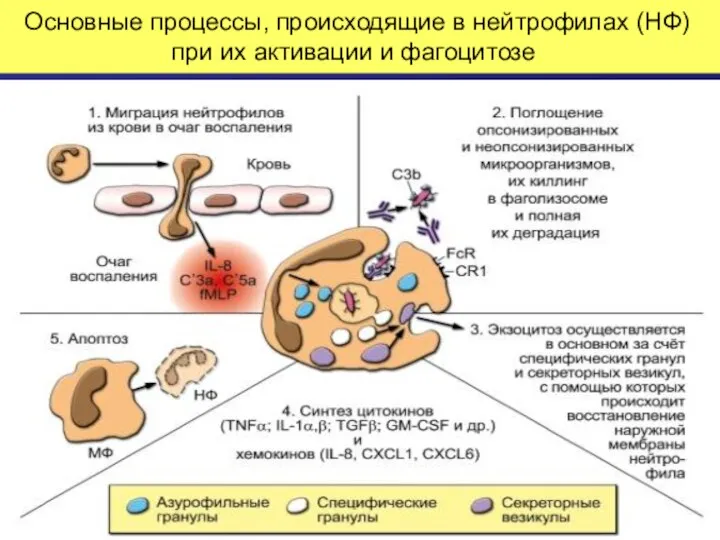
Основные процессы, происходящие в нейтрофилах (НФ) при их активации и фагоцитозе
Основные процессы, происходящие в нейтрофилах (НФ) при их активации и фагоцитозе
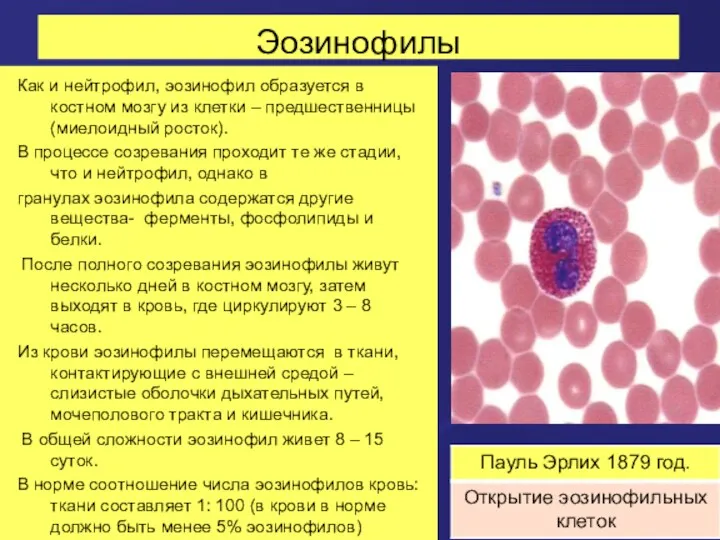
Эозинофилы
Как и нейтрофил, эозинофил образуется в костном мозгу из клетки –
Эозинофилы
Как и нейтрофил, эозинофил образуется в костном мозгу из клетки –
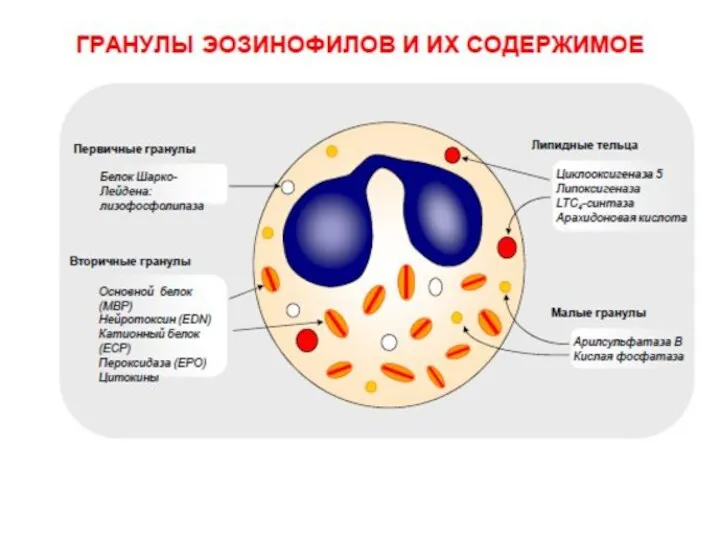
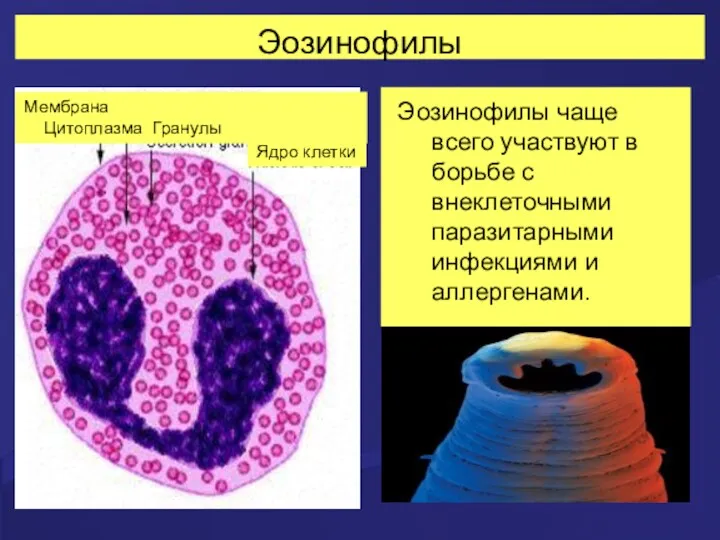
Эозинофилы
Эозинофилы чаще всего участвуют в борьбе с внеклеточными паразитарными инфекциями и
Эозинофилы
Эозинофилы чаще всего участвуют в борьбе с внеклеточными паразитарными инфекциями и

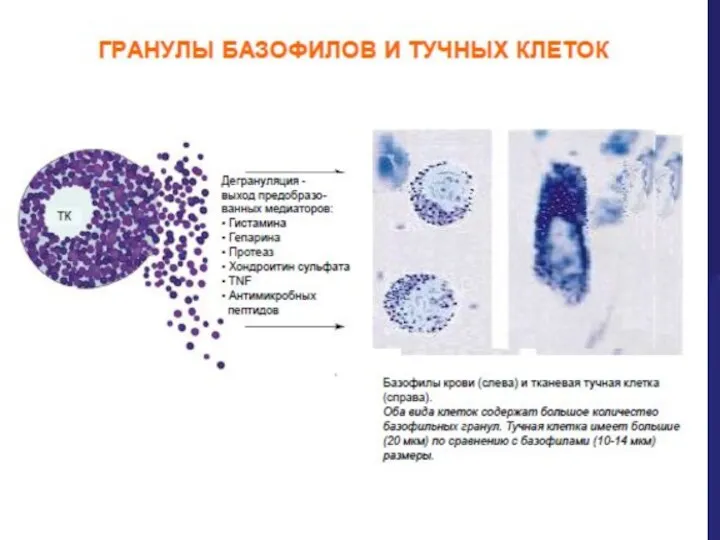
Базофилы и тучные клетки(ТК)
Тучные клетки при активации способны освобождать в окружающее
Базофилы и тучные клетки(ТК)
Тучные клетки при активации способны освобождать в окружающее
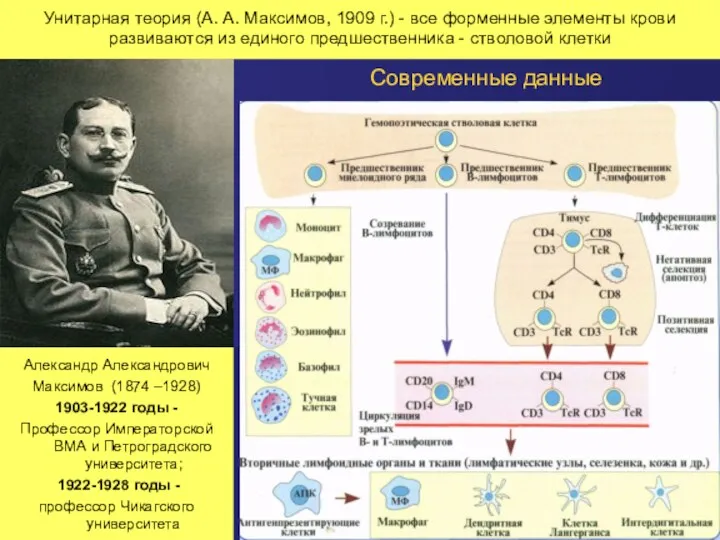
Унитарная теория (А. А. Максимов, 1909 г.) - все форменные элементы
Унитарная теория (А. А. Максимов, 1909 г.) - все форменные элементы

Макрофаги – потомки моноцитов крови
Моноциты, макрофаги, нейтрофилы происходят от клеток –предшественников
Макрофаги – потомки моноцитов крови
Моноциты, макрофаги, нейтрофилы происходят от клеток –предшественников
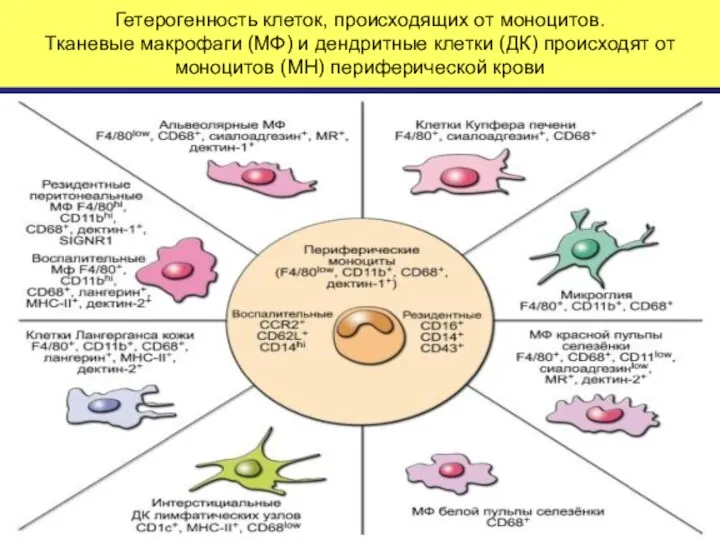
Гетерогенность клеток, происходящих от моноцитов.
Тканевые макрофаги (МФ) и дендритные клетки
Гетерогенность клеток, происходящих от моноцитов. Тканевые макрофаги (МФ) и дендритные клетки

Макрофаг х 5000
Макрофаги во всех органах и тканях способны к :
антигенной
Макрофаг х 5000
Макрофаги во всех органах и тканях способны к :
антигенной

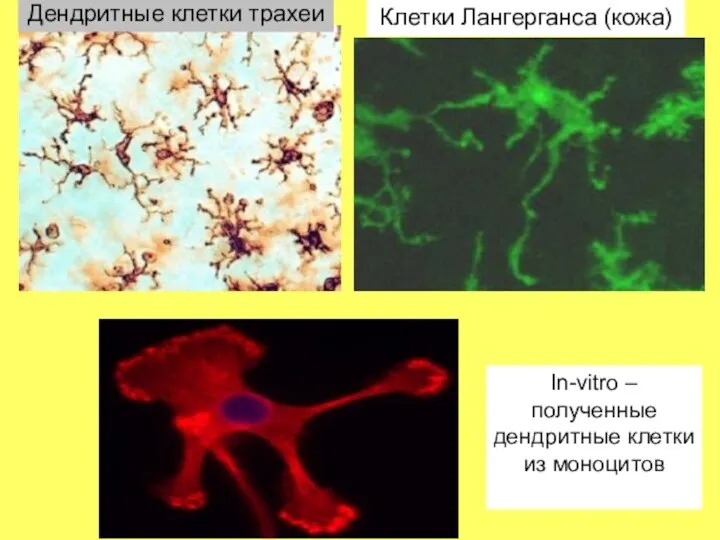
Дендритная клетка х 5000
Дендритные клетки- самые важные антигенпрезентирующие клетки (АПК),
так
Дендритная клетка х 5000
Дендритные клетки- самые важные антигенпрезентирующие клетки (АПК),
так
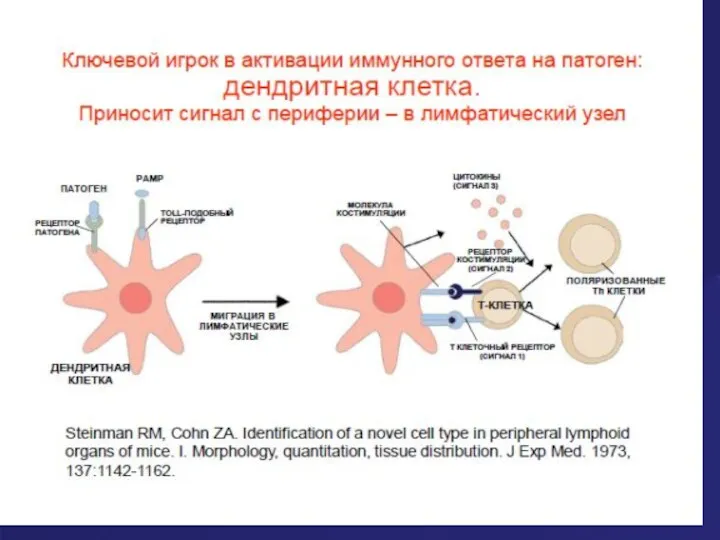

Естественные киллеры (NK) –клетки врожденного иммунитета
Большие гранулярные лимфоциты («нулевые клетки», «не
Естественные киллеры (NK) –клетки врожденного иммунитета
Большие гранулярные лимфоциты («нулевые клетки», «не

Естественные киллеры (NK)
Число NK в крови - 5-15%; в лимфоузлах
Естественные киллеры (NK)
Число NK в крови - 5-15%; в лимфоузлах
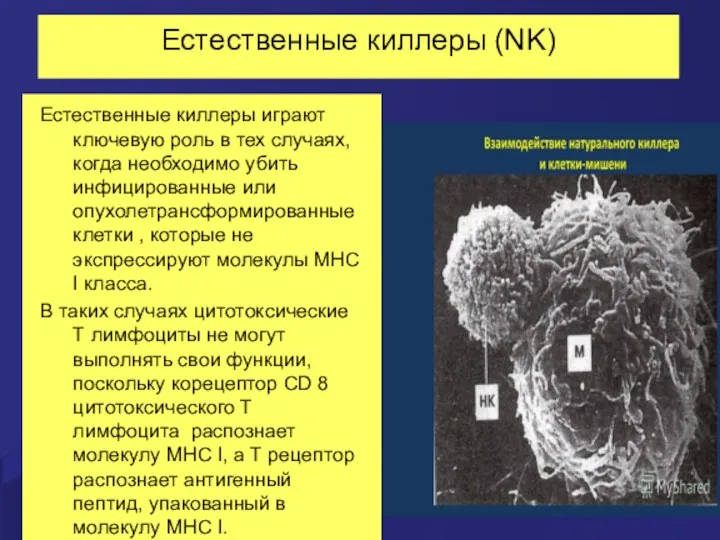
Естественные киллеры (NK)
Естественные киллеры играют ключевую роль в тех случаях, когда
Естественные киллеры (NK)
Естественные киллеры играют ключевую роль в тех случаях, когда

Рецепторы естественных киллеров: для лизиса клетки-мишени недостаточно одной только отсутствия подавляющего
Рецепторы естественных киллеров: для лизиса клетки-мишени недостаточно одной только отсутствия подавляющего
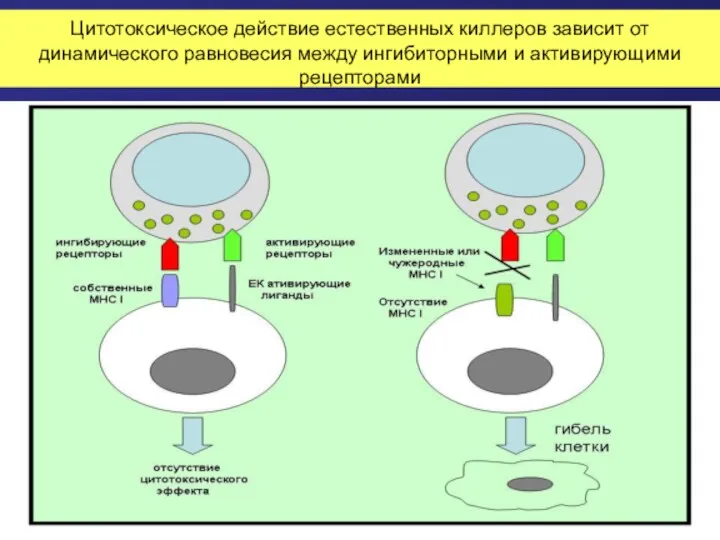
Цитотоксическое действие естественных киллеров зависит от динамического равновесия между ингибиторными и
Цитотоксическое действие естественных киллеров зависит от динамического равновесия между ингибиторными и
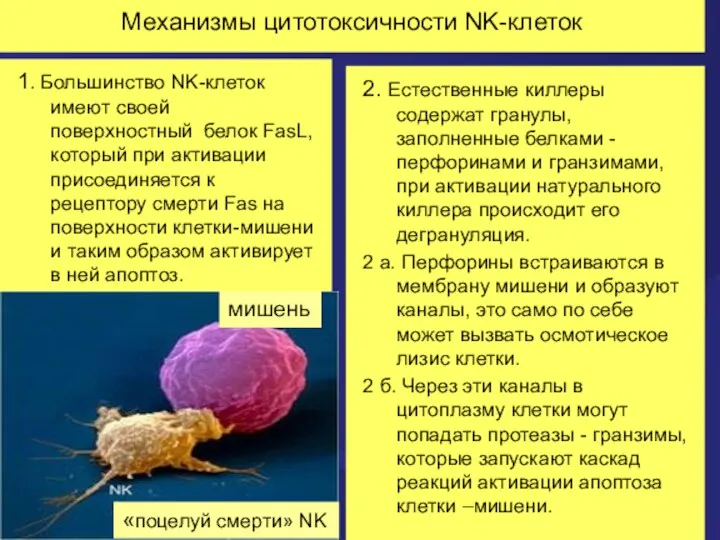
Механизмы цитотоксичности NK-клеток
1. Большинство NK-клеток имеют своей поверхностный белок FasL,
Механизмы цитотоксичности NK-клеток
1. Большинство NK-клеток имеют своей поверхностный белок FasL,
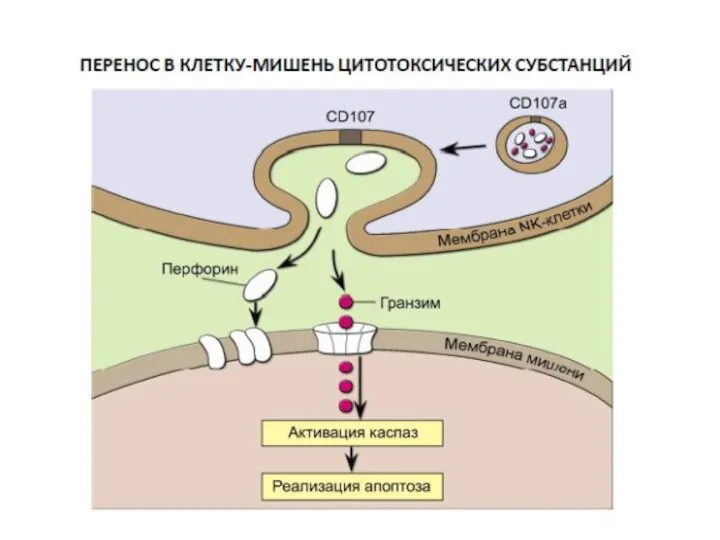
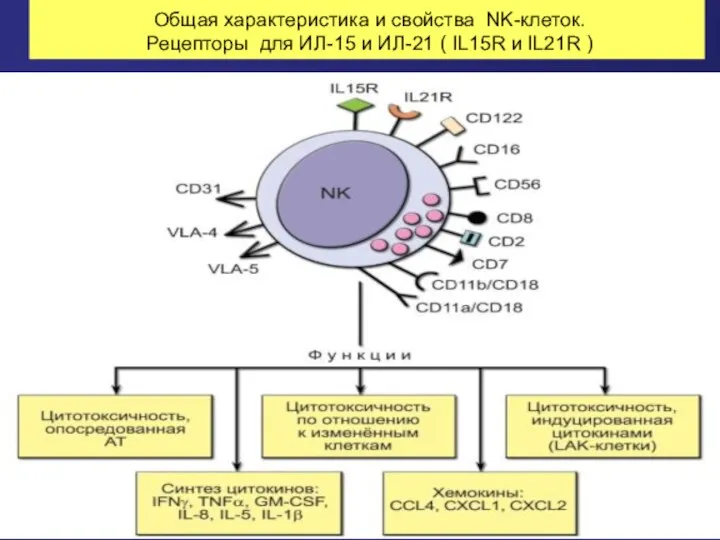
Общая характеристика и свойства NK-клеток.
Рецепторы для ИЛ-15 и ИЛ-21 (
Общая характеристика и свойства NK-клеток. Рецепторы для ИЛ-15 и ИЛ-21 (

Гуморальные факторы врожденного иммунитета - дефенсины
Дефензины (от англ. Defens защита) - катионные пептиды,
Гуморальные факторы врожденного иммунитета - дефенсины
Дефензины (от англ. Defens защита) - катионные пептиды,

Гуморальные факторы врожденного иммунитета – лизоцим
Лизоцим (мурамидаза) — фермент, обладающий специфической
Гуморальные факторы врожденного иммунитета – лизоцим
Лизоцим (мурамидаза) — фермент, обладающий специфической

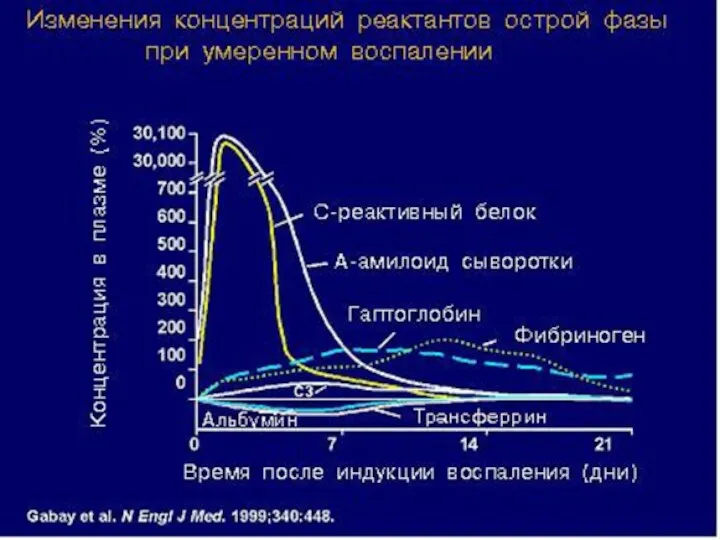
Гуморальные факторы врожденного иммунитета – белки острой фазы
Осторофазовые белки – большая
Гуморальные факторы врожденного иммунитета – белки острой фазы
Осторофазовые белки – большая
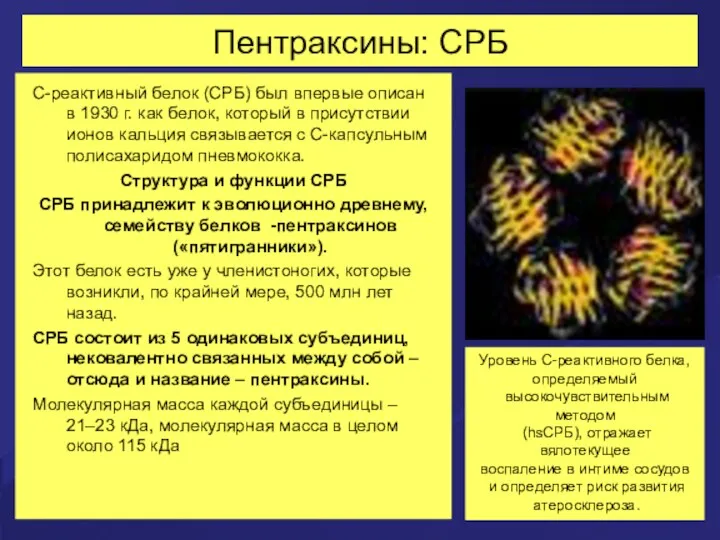
Пентраксины: СРБ
С-реактивный белок (СРБ) был впервые описан в 1930 г. как
Пентраксины: СРБ
С-реактивный белок (СРБ) был впервые описан в 1930 г. как
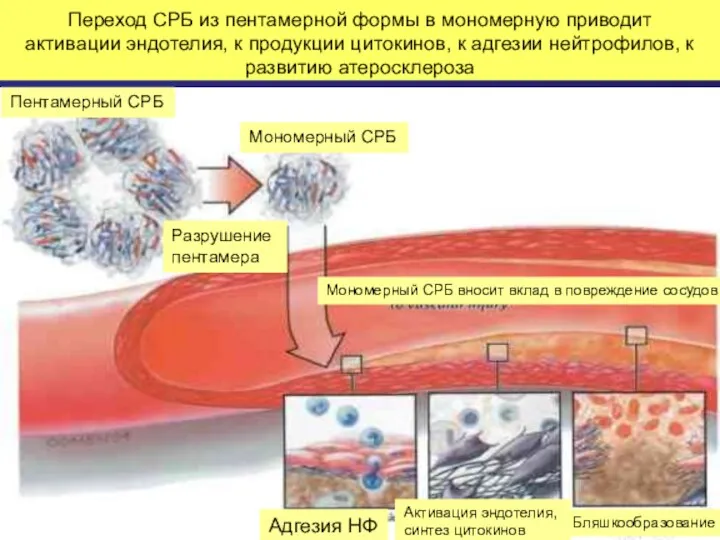
Переход СРБ из пентамерной формы в мономерную приводит
активации эндотелия, к
Переход СРБ из пентамерной формы в мономерную приводит активации эндотелия, к
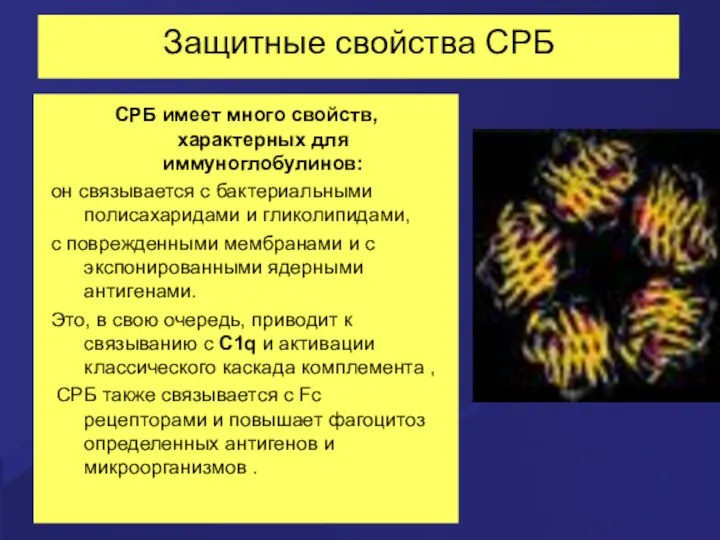
Защитные свойства СРБ
СРБ имеет много свойств, характерных для иммуноглобулинов:
он связывается
Защитные свойства СРБ
СРБ имеет много свойств, характерных для иммуноглобулинов:
он связывается
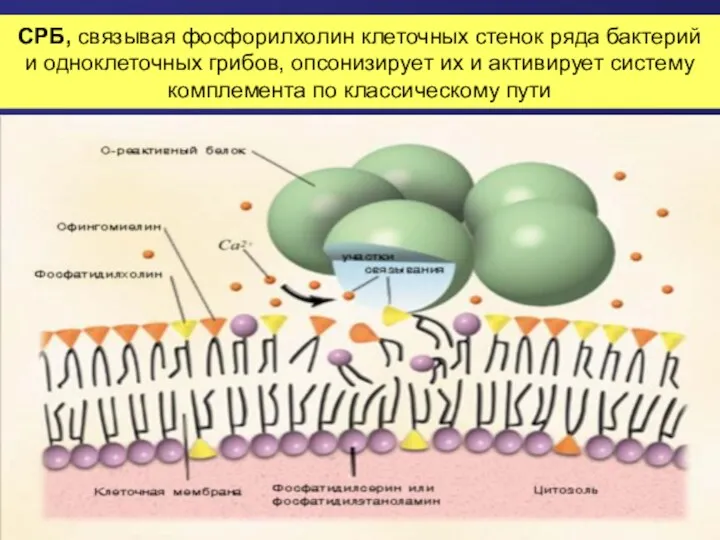
СРБ, связывая фосфорилхолин клеточных стенок ряда бактерий и одноклеточных грибов, опсонизирует их
СРБ, связывая фосфорилхолин клеточных стенок ряда бактерий и одноклеточных грибов, опсонизирует их
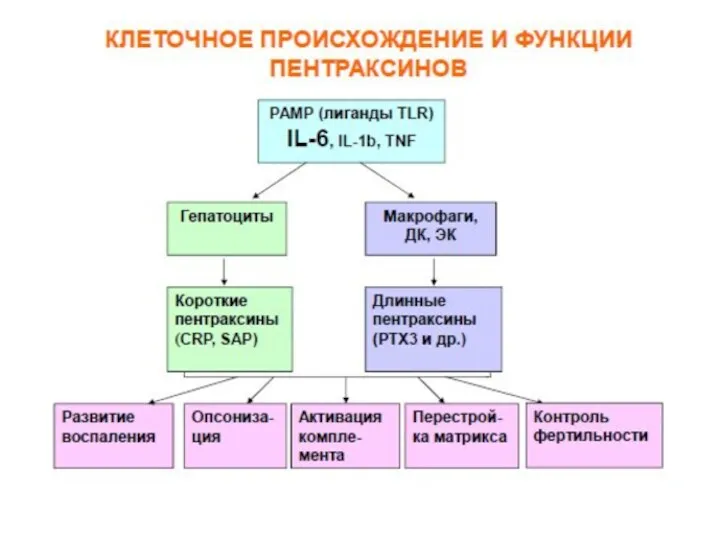

Гуморальные факторы врожденного иммунитета: цитокины
Цитокины — составная часть молекулярных механизмов как врожденного,
Гуморальные факторы врожденного иммунитета: цитокины
Цитокины — составная часть молекулярных механизмов как врожденного,
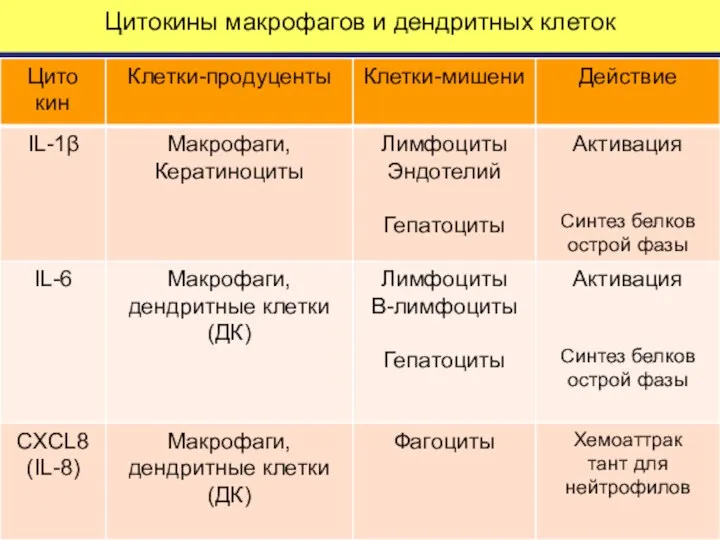
Цитокины макрофагов и дендритных клеток
Цитокины макрофагов и дендритных клеток
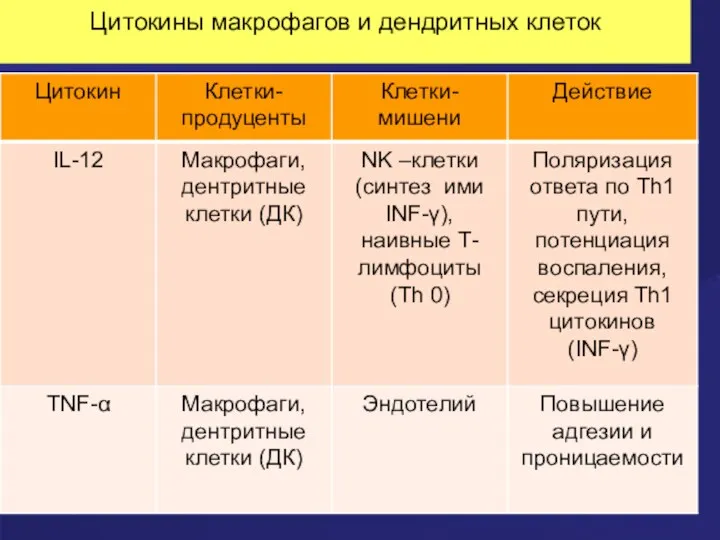
Цитокины макрофагов и дендритных клеток
Цитокины макрофагов и дендритных клеток

Гуморальные факторы врожденного иммунитета - интерфероны 1 типа –противовирусная активность
Клетки врожденного
Гуморальные факторы врожденного иммунитета - интерфероны 1 типа –противовирусная активность
Клетки врожденного

Роль цитокинов дендритных клеток (ДК) в запуске адаптивного иммунного ответа (этот
Роль цитокинов дендритных клеток (ДК) в запуске адаптивного иммунного ответа (этот

СИСТЕМА КОМПЛЕМЕНТА –основная система гуморальных факторов врожденного иммунитета
Термин «комплемент» ввёл Пауль Эрлих
СИСТЕМА КОМПЛЕМЕНТА –основная система гуморальных факторов врожденного иммунитета
Термин «комплемент» ввёл Пауль Эрлих

Классический путь активации СК
Последовательность открытия компонентов системы комплемента не соответствует очередности
Классический путь активации СК
Последовательность открытия компонентов системы комплемента не соответствует очередности
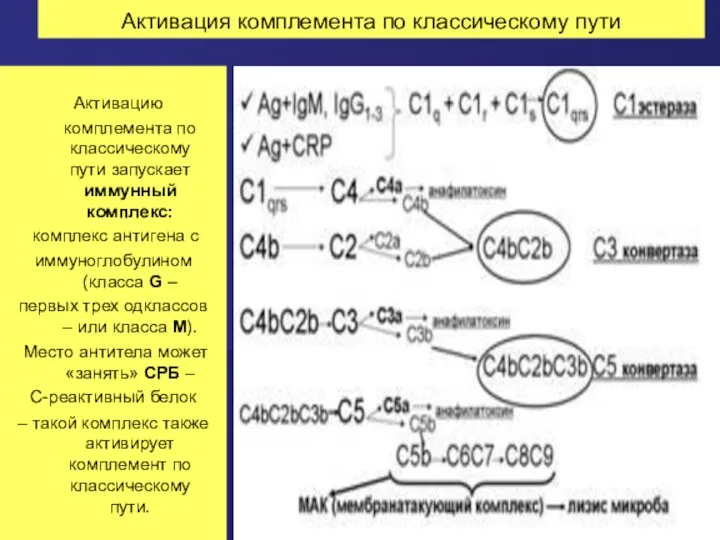
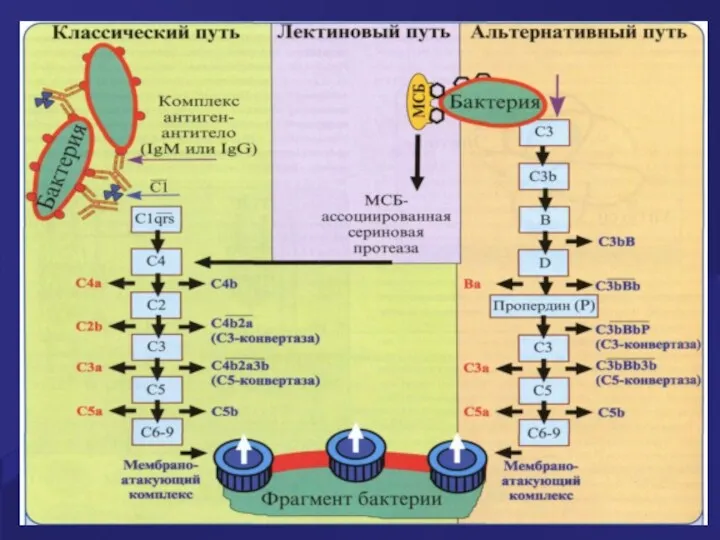
Активация комплемента по классическому пути
Активацию комплемента по классическому пути запускает
Активация комплемента по классическому пути
Активацию комплемента по классическому пути запускает

Каскад активации системы комплемента по классическому пути
а. Сначала активируется фракция С1:
Каскад активации системы комплемента по классическому пути
а. Сначала активируется фракция С1:
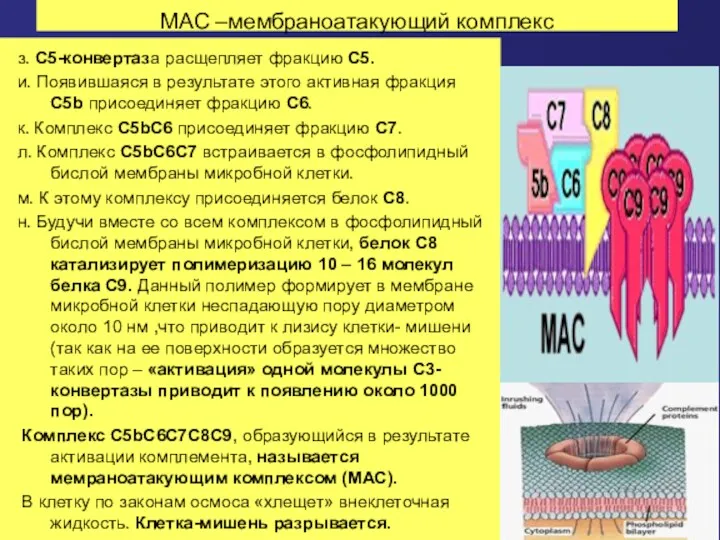
МАС –мембраноатакующий комплекс
з. С5-конвертаза расщепляет фракцию С5.
и. Появившаяся в результате этого
МАС –мембраноатакующий комплекс
з. С5-конвертаза расщепляет фракцию С5.
и. Появившаяся в результате этого

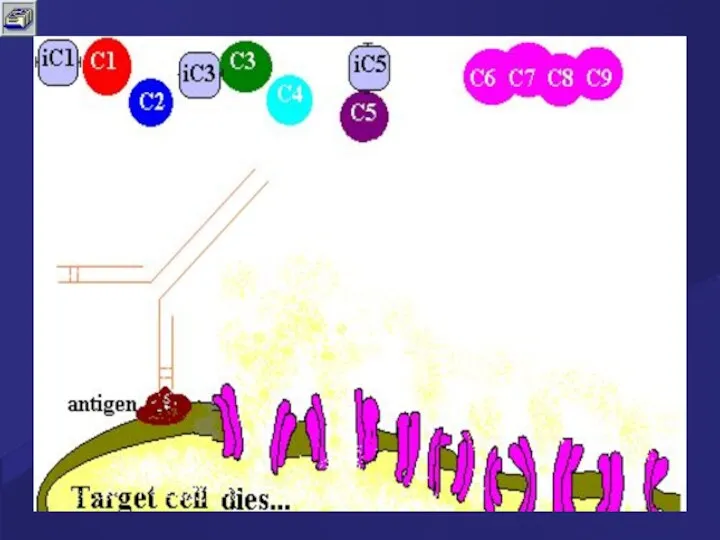
Поверхность клетки – мишени, атакованная МАС
Поверхность клетки – мишени, атакованная МАС

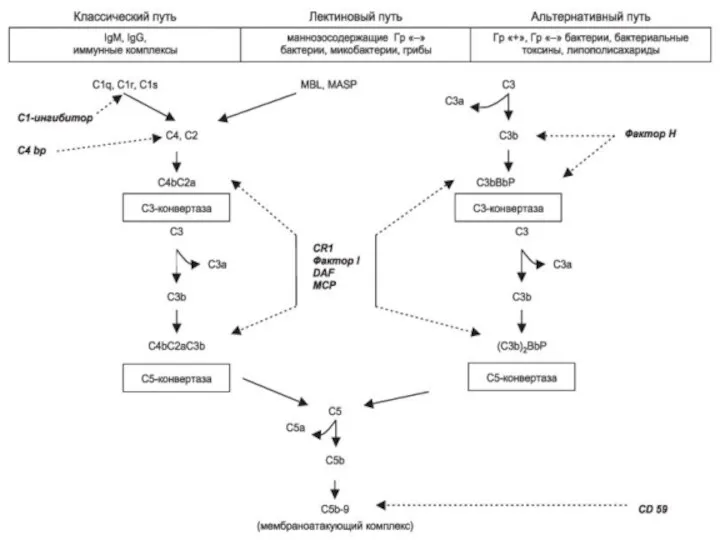
Пути активации системы комплемента
Классический путь- филогенетически более новый (так как появился
Пути активации системы комплемента
Классический путь- филогенетически более новый (так как появился

Врожденный и приобретенный иммунитет: основные различия
Врожденный
1. Существует в любом организме до
Врожденный и приобретенный иммунитет: основные различия
Врожденный
1. Существует в любом организме до

Врожденный и приобретенный иммунитет: основные различия
Врожденный
3. Все клетки врожденного иммунитета активируются
Врожденный и приобретенный иммунитет: основные различия
Врожденный
3. Все клетки врожденного иммунитета активируются

Врожденный и приобретенный иммунитет: основные различия
Врожденный
4. Реакции врожденного иммунитета разворачиваются почти
Врожденный и приобретенный иммунитет: основные различия
Врожденный
4. Реакции врожденного иммунитета разворачиваются почти

Врожденный и приобретенный иммунитет: основные различия
Врожденный
5. Во врожденном иммунитете иммунологическая память
Врожденный и приобретенный иммунитет: основные различия
Врожденный
5. Во врожденном иммунитете иммунологическая память
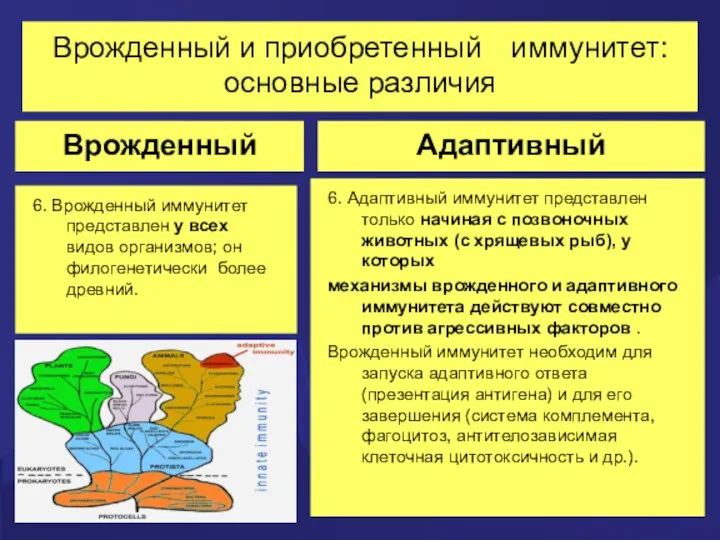
Врожденный и приобретенный иммунитет: основные различия
Врожденный
6. Врожденный иммунитет представлен у всех
Врожденный и приобретенный иммунитет: основные различия
Врожденный
6. Врожденный иммунитет представлен у всех

Дженуэй Чарльз А. – младший: постулаты
Чарльз А. Дженуэй-младший (1943-2003), автор
Дженуэй Чарльз А. – младший: постулаты
Чарльз А. Дженуэй-младший (1943-2003), автор


Дженуэй Чарльз А. – младший: постулаты
1. Врожденный иммунитет защищает все виды
Дженуэй Чарльз А. – младший: постулаты
1. Врожденный иммунитет защищает все виды

Дженуэй Чарльз А. – младший: постулаты
3. Рецепторов, распознающих «чужое» во врожденном
Дженуэй Чарльз А. – младший: постулаты
3. Рецепторов, распознающих «чужое» во врожденном

Дженуэй Чарльз А. – младший: постулаты
5 . Для запуска эффективного иммунного
Дженуэй Чарльз А. – младший: постулаты
5 . Для запуска эффективного иммунного

Дженуэй Чарльз А. – младший: постулаты
7. Образраспознающие рецепторы клеток врожденного иммунитета
Дженуэй Чарльз А. – младший: постулаты
7. Образраспознающие рецепторы клеток врожденного иммунитета

Дженуэй Чарльз А. – младший: постулаты
8. Сигнал 2 (костимуляция) запускает ответ
Дженуэй Чарльз А. – младший: постулаты
8. Сигнал 2 (костимуляция) запускает ответ

Вопросы занятия №2
Определение врожденного иммунитета.
Свойства врожденного иммунитета.
Клетки врожденного иммунитета.
Рецепторы клеток врожденного
Вопросы занятия №2
Определение врожденного иммунитета.
Свойства врожденного иммунитета.
Клетки врожденного иммунитета.
Рецепторы клеток врожденного

Тестовые задания к занятию №2
11. К клеточным факторам врожденного иммунитета не
Тестовые задания к занятию №2
11. К клеточным факторам врожденного иммунитета не

Тестовые задания к занятию №2
3. Распознавание во врожденном иммунитете:
грубое
«образное»
«по отсутствию
Тестовые задания к занятию №2
3. Распознавание во врожденном иммунитете:
грубое
«образное»
«по отсутствию

Тестовые задания к занятию №2
5. Во врожденном иммунитете:
после окончания иммунного ответа
Тестовые задания к занятию №2
5. Во врожденном иммунитете:
после окончания иммунного ответа

Тестовые задания к занятию № 2
7.К фагоцитирующим клеткам не относятся:
Макрофаги
Дендритные клетки
Нейтрофилы
Моноциты
Тромбоциты
8.В
Тестовые задания к занятию № 2
7.К фагоцитирующим клеткам не относятся:
Макрофаги
Дендритные клетки
Нейтрофилы
Моноциты
Тромбоциты
8.В
 Инфузионная терапия при остром повреждении почек: как найти золотую середину
Инфузионная терапия при остром повреждении почек: как найти золотую середину Теории и фазы стресса. Особенности у детей и подростков
Теории и фазы стресса. Особенности у детей и подростков Вибрационная болезнь и нейросенсорная тугоухость
Вибрационная болезнь и нейросенсорная тугоухость Балалардағы пародонт аурулары. Пародонт ауруларының жіктелуі. Этиологиясы, диагностика әдістері
Балалардағы пародонт аурулары. Пародонт ауруларының жіктелуі. Этиологиясы, диагностика әдістері Возможности магнитнорезонансной томографии в диагностике патологических состояний коленного сустава
Возможности магнитнорезонансной томографии в диагностике патологических состояний коленного сустава О вреде абортов
О вреде абортов Общественное здоровье. Основные понятия. (Лекция 2)
Общественное здоровье. Основные понятия. (Лекция 2) Анемия. Анемия түрлері
Анемия. Анемия түрлері Герпесвирусы
Герпесвирусы Врач-уролог
Врач-уролог Гиперфосфатемия у пациентов с ХБП. Профилатика и лечение
Гиперфосфатемия у пациентов с ХБП. Профилатика и лечение Сестринская помощь пациентам с впервые выявленной бронхиальной астмой
Сестринская помощь пациентам с впервые выявленной бронхиальной астмой Молекулярно-цитогенетическая диагностика острых лейкозов
Молекулярно-цитогенетическая диагностика острых лейкозов Основи мікробіології
Основи мікробіології Личная гигиена
Личная гигиена Сүт безінің қатерсіз ісіктері
Сүт безінің қатерсіз ісіктері Развитие глотки, пищевода, желудка, кишечника
Развитие глотки, пищевода, желудка, кишечника Гломерулярные болезни почек. Патология почечных клубочков при системных заболеваниях
Гломерулярные болезни почек. Патология почечных клубочков при системных заболеваниях Особенности травматизма детского возраста
Особенности травматизма детского возраста Судебно-медицинская экспертиза при повреждениях от воздействия тупых орудий
Судебно-медицинская экспертиза при повреждениях от воздействия тупых орудий Уход за полостью рта
Уход за полостью рта Суставной синдром
Суставной синдром Общественное здоровье. Статистика заболеваемости. Источники получения информации
Общественное здоровье. Статистика заболеваемости. Источники получения информации Нарушения менструального цикла. Дисфункциональные маточные кровотечения
Нарушения менструального цикла. Дисфункциональные маточные кровотечения Роль оздоровительной физической культуры в современных условиях
Роль оздоровительной физической культуры в современных условиях Сортировка пострадавших в ЧС с механической травмой
Сортировка пострадавших в ЧС с механической травмой Биологиялық жүйелер люминесценциясы. (Дәріс 15)
Биологиялық жүйелер люминесценциясы. (Дәріс 15) Оздоровчі сили природи та гігієнічні чинники. Лекція 6
Оздоровчі сили природи та гігієнічні чинники. Лекція 6