- ДНК и РНК. Устройство генома

Содержание
- 2. Схема молекулы ДНК Расстояние между нуклеотидами – 0,34 нм. Диаметр молекулы – 2 нм. Число нуклеотидов
- 3. Молекула ДНК полярна
- 4. Кариотип и хромосома Структура хромосомы: плечи, первичная перетяжка, теломеры.
- 5. ДНК + белки=хроматин Хроматин упакован в хромосомы Каждая хромосома – 1 молекула ДНК В – S
- 6. Хромосомы человека, дифференциальная окраска
- 7. Кариотип человека - таблица
- 8. Генетическая неоднородность хромосом Большая часть ДНК у эукариот представлена некодирующими последовательностями (свыше 90%). Расположение генов, кодирующих
- 9. Функциональная неоднородность хромосом Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными рекомбинациями (перестановками). Часто встречающиеся
- 10. Функциональная неоднородность хромосом Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными рекомбинациями (перестановками). Часто встречающиеся
- 11. Каждый кариотип уникален
- 12. Хромосомные территории в ядре
- 13. 18 и 19 хромосомы в ядре лимфоцита человека
- 14. Плечи хромосом также занимают в интерфазном ядре определенные территории
- 15. Строение генома прокариот Кольцевая молекула ДНК (одна) Кодирующие РНК участки – гены составляют основную часть генома
- 16. Несколько линейных молекул ДНК (хромосом). Кодирующие РНК участки составляют малую часть генома (~1,5% у человека). Каждый
- 17. Прокариоты – от 500 до 7500 генов; длина – 0,5-10 млн. нуклеотидов. Эукариоты – от 6300
- 18. Структура выделенной хроматиновой фибриллы А – в низкой ионной силе, В – в высокой ионной силе
- 19. ДНК и гистоны образуют нуклеосомы (первый уровень укладки хроматина)
- 20. Модель хроматиновой 30-нм фибриллы – основную роль играет гистон H1
- 21. Модификации хвостов гистонов изменяют компактность хроматина
- 22. Хроматин Эухроматин (транскрипционно активный) Хроматин – комплекс ДНК и белков (~40:60) с небольшим количеством РНК (около
- 23. Гетерохроматин конститутивный – связан со специальными участками ДНК (например, сателлитная ДНК) и конденсирован во всех клетках
- 24. Основные негистоновые белки хроматина ДНК-полимераза и связанные с ней ферменты Белок гетерохроматина 1 (HP1) – отвечает
- 25. Структурные белки хроматина Smc-белки (structural maintenance of chromosomes) – эволюционно консервативные белки, связывающиеся с ДНК при
- 26. Структурные белки хромосом Когезины – внутри хромосомы, конденсины – межхромосомные взаимодействия гомологов. Все образуют гомодимеры с
- 27. Когезины и конденсины Когезин (слева) образует межмолекулярные связи с ДНК (связь между хроматидами, которая рвется в
- 28. Конденсины в митозе
- 29. Динамика хроматина В интерфазном ядре хроматин пластичен – его структура постоянно изменяется в связи с активацией
- 30. Хроматин в ядрах животных
- 32. Скачать презентацию
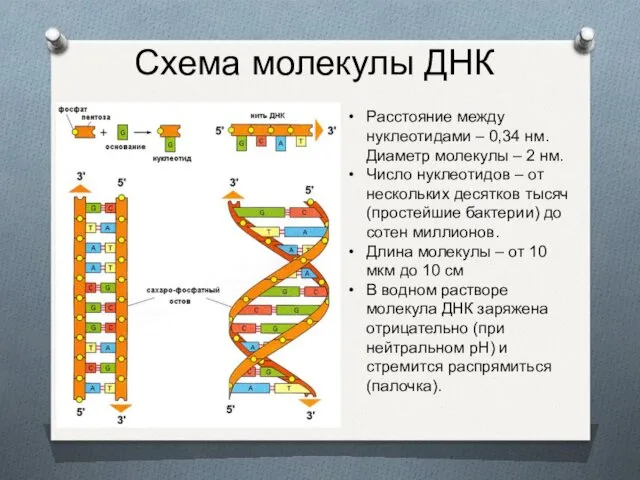
Схема молекулы ДНК
Расстояние между нуклеотидами – 0,34 нм. Диаметр молекулы –
Схема молекулы ДНК
Расстояние между нуклеотидами – 0,34 нм. Диаметр молекулы –
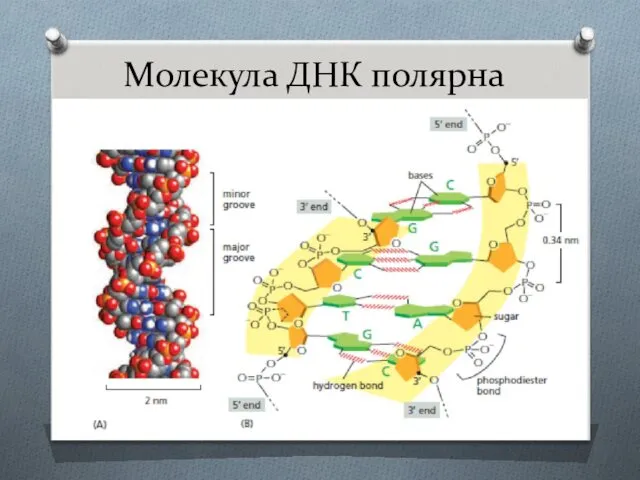
Молекула ДНК полярна
Молекула ДНК полярна

Кариотип и хромосома
Структура хромосомы: плечи, первичная перетяжка, теломеры.
Кариотип и хромосома
Структура хромосомы: плечи, первичная перетяжка, теломеры.
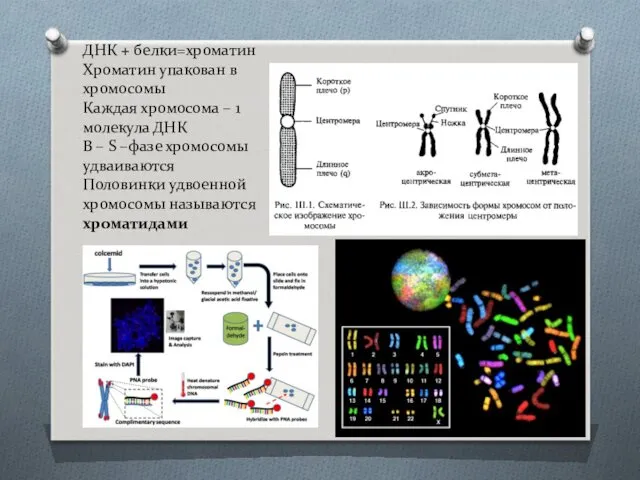
ДНК + белки=хроматин
Хроматин упакован в хромосомы
Каждая хромосома – 1 молекула ДНК
В
ДНК + белки=хроматин Хроматин упакован в хромосомы Каждая хромосома – 1 молекула ДНК В

Хромосомы человека, дифференциальная окраска
Хромосомы человека, дифференциальная окраска
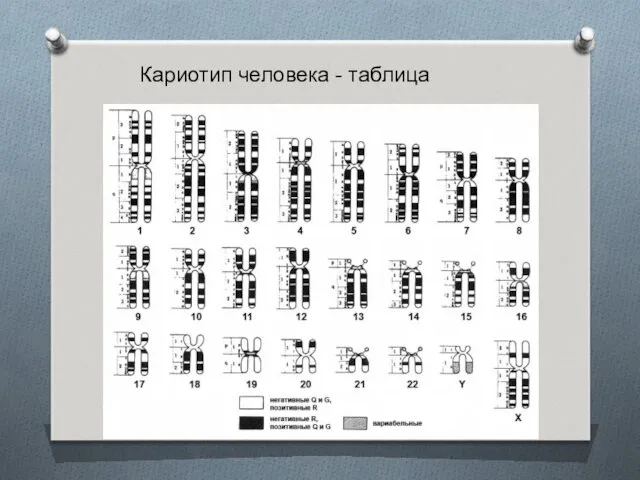
Кариотип человека - таблица
Кариотип человека - таблица

Генетическая неоднородность хромосом
Большая часть ДНК у эукариот представлена некодирующими последовательностями (свыше
Генетическая неоднородность хромосом
Большая часть ДНК у эукариот представлена некодирующими последовательностями (свыше

Функциональная неоднородность хромосом
Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными
Функциональная неоднородность хромосом
Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными

Функциональная неоднородность хромосом
Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными
Функциональная неоднородность хромосом
Неоднородность хромосом доказывается различной частотой мутаций (разрывов) и неслучайными

Каждый кариотип уникален
Каждый кариотип уникален
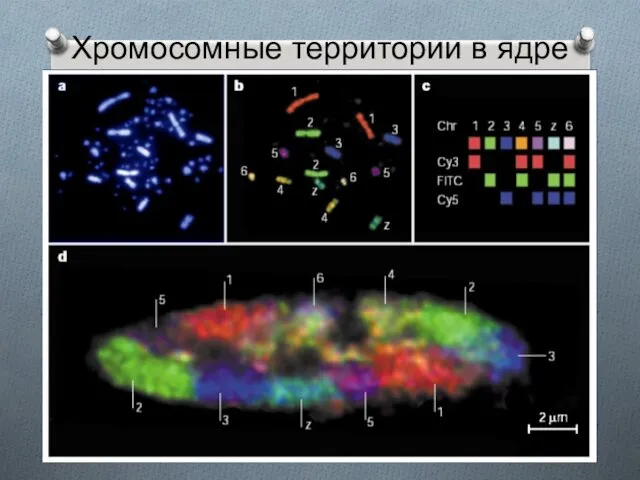
Хромосомные территории в ядре
Хромосомные территории в ядре
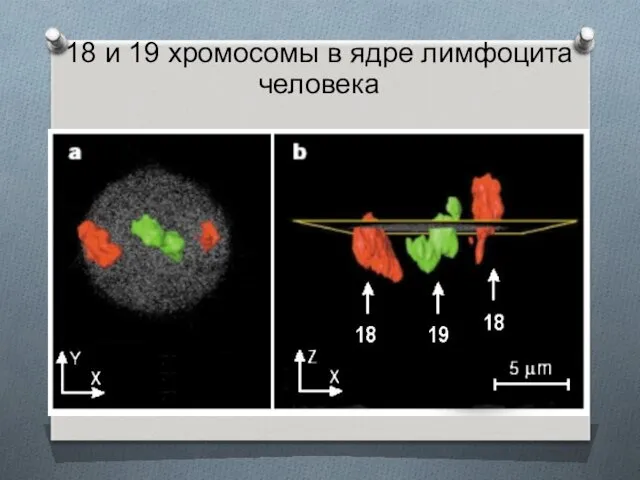
18 и 19 хромосомы в ядре лимфоцита человека
18 и 19 хромосомы в ядре лимфоцита человека
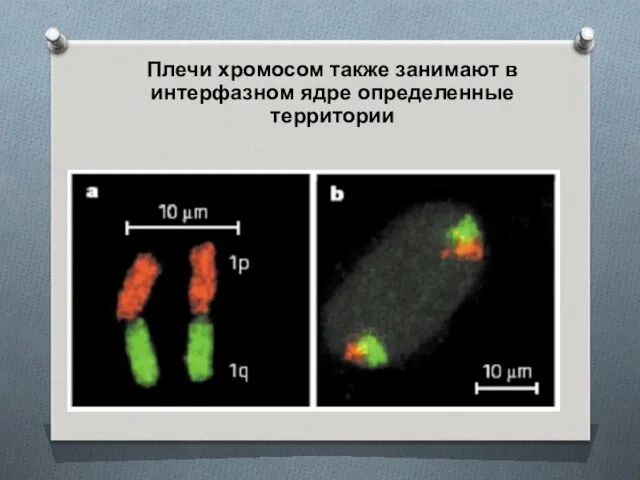
Плечи хромосом также занимают в интерфазном ядре определенные территории
Плечи хромосом также занимают в интерфазном ядре определенные территории

Строение генома прокариот
Кольцевая молекула ДНК (одна)
Кодирующие РНК участки – гены
Строение генома прокариот
Кольцевая молекула ДНК (одна)
Кодирующие РНК участки – гены

Несколько линейных молекул ДНК (хромосом). Кодирующие РНК участки составляют малую часть
Несколько линейных молекул ДНК (хромосом). Кодирующие РНК участки составляют малую часть

Прокариоты – от 500 до 7500 генов; длина – 0,5-10 млн.
Прокариоты – от 500 до 7500 генов; длина – 0,5-10 млн.
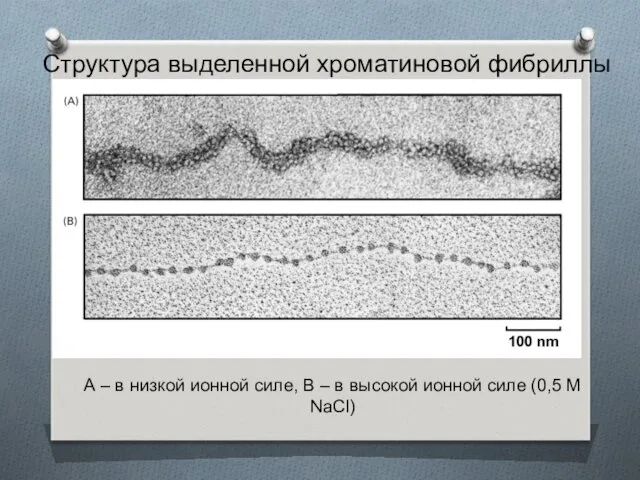
Структура выделенной хроматиновой фибриллы
А – в низкой ионной силе, В –
Структура выделенной хроматиновой фибриллы
А – в низкой ионной силе, В –
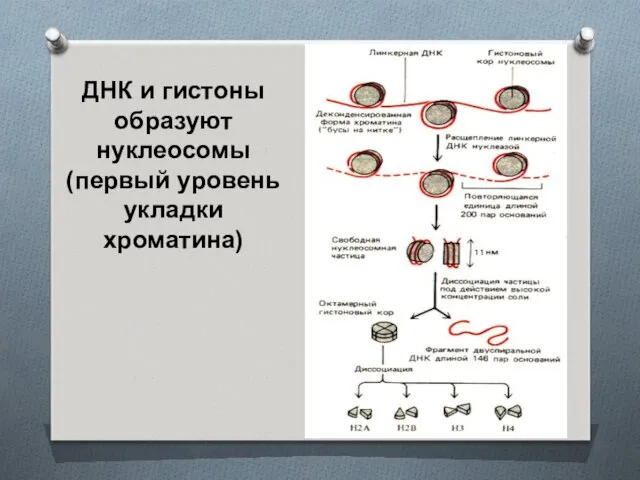
ДНК и гистоны образуют нуклеосомы (первый уровень укладки хроматина)
ДНК и гистоны образуют нуклеосомы (первый уровень укладки хроматина)
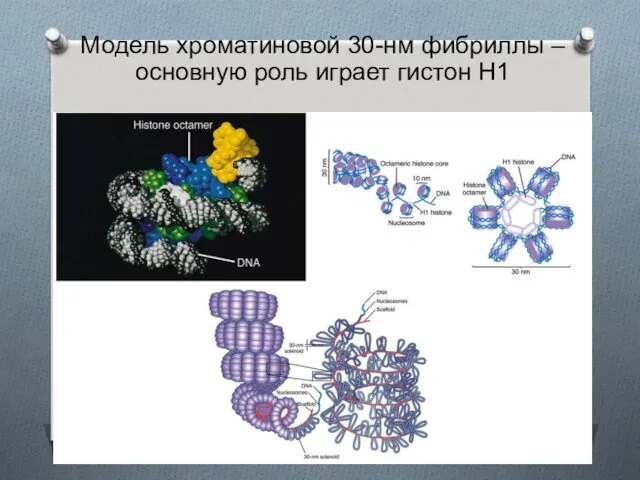
Модель хроматиновой 30-нм фибриллы – основную роль играет гистон H1
Модель хроматиновой 30-нм фибриллы – основную роль играет гистон H1
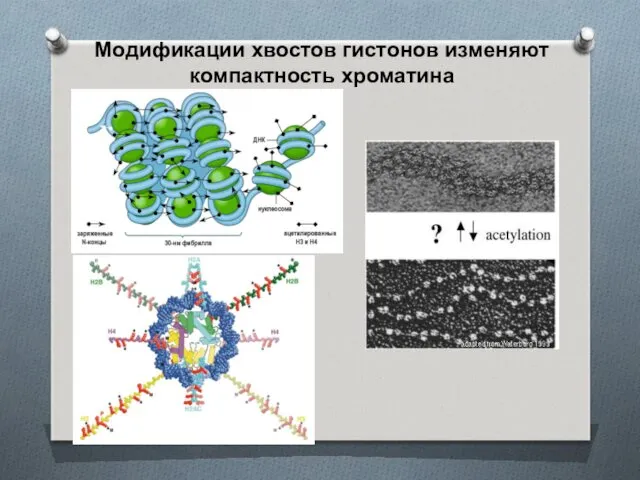
Модификации хвостов гистонов изменяют компактность хроматина
Модификации хвостов гистонов изменяют компактность хроматина
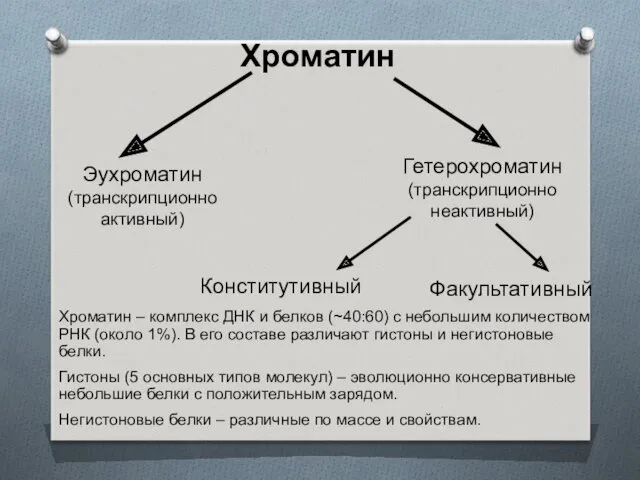
Хроматин
Эухроматин
(транскрипционно
активный)
Хроматин – комплекс ДНК и белков (~40:60) с небольшим количеством
Хроматин
Эухроматин
(транскрипционно
активный)
Хроматин – комплекс ДНК и белков (~40:60) с небольшим количеством
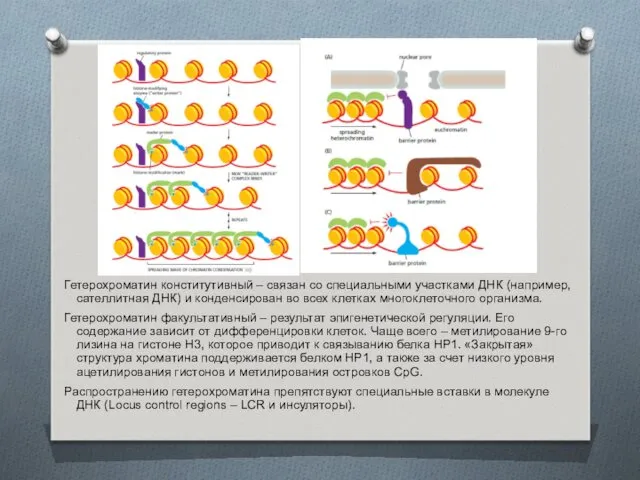
Гетерохроматин конститутивный – связан со специальными участками ДНК (например, сателлитная ДНК)
Гетерохроматин конститутивный – связан со специальными участками ДНК (например, сателлитная ДНК)

Основные негистоновые белки хроматина
ДНК-полимераза и связанные с ней ферменты
Белок гетерохроматина 1
Основные негистоновые белки хроматина
ДНК-полимераза и связанные с ней ферменты
Белок гетерохроматина 1

Структурные белки хроматина
Smc-белки (structural maintenance of chromosomes) – эволюционно консервативные белки,
Структурные белки хроматина
Smc-белки (structural maintenance of chromosomes) – эволюционно консервативные белки,
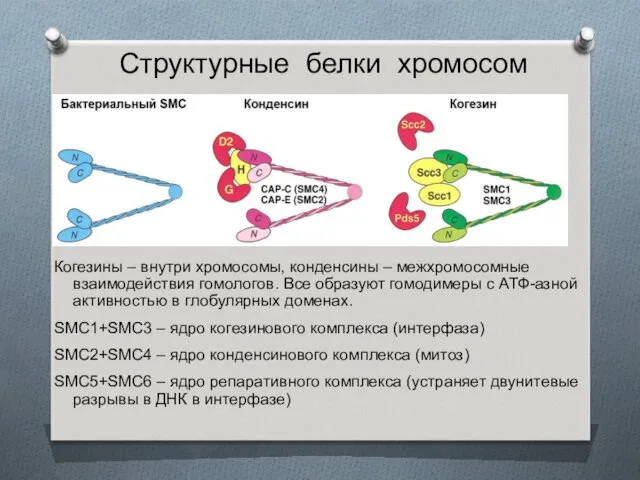
Структурные белки хромосом
Когезины – внутри хромосомы, конденсины – межхромосомные взаимодействия гомологов.
Структурные белки хромосом
Когезины – внутри хромосомы, конденсины – межхромосомные взаимодействия гомологов.
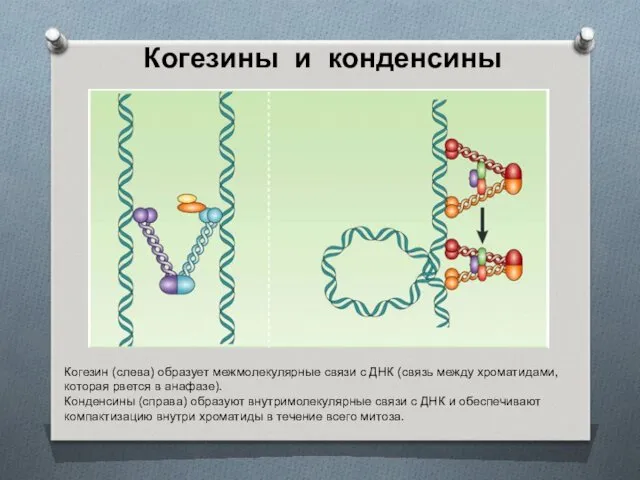
Когезины и конденсины
Когезин (слева) образует межмолекулярные связи с ДНК (связь между
Когезины и конденсины
Когезин (слева) образует межмолекулярные связи с ДНК (связь между
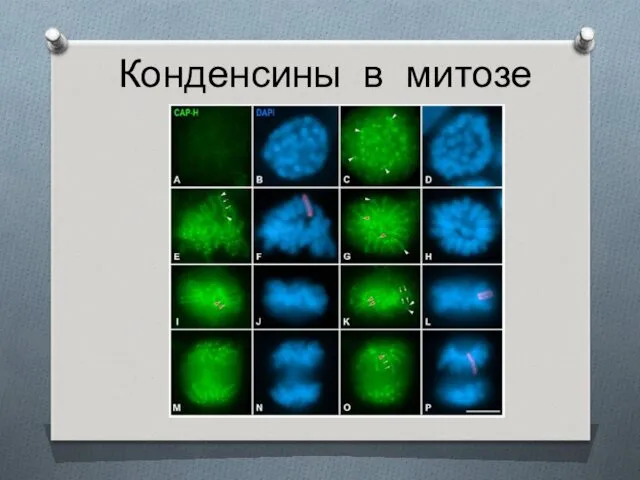
Конденсины в митозе
Конденсины в митозе
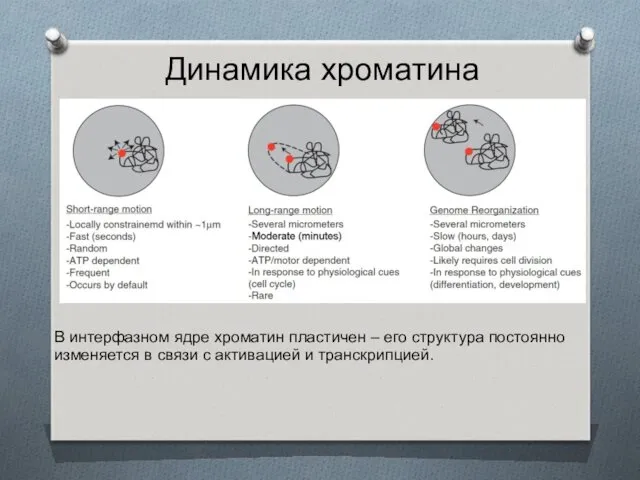
Динамика хроматина
В интерфазном ядре хроматин пластичен – его структура постоянно изменяется
Динамика хроматина
В интерфазном ядре хроматин пластичен – его структура постоянно изменяется
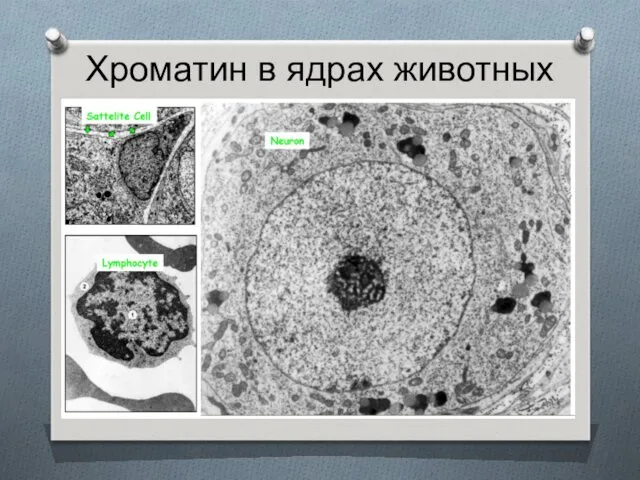
Хроматин в ядрах животных
Хроматин в ядрах животных
 Страны Северной Европы
Страны Северной Европы Ұлпа қабынуы туралы түсінік ұлпа қабынуын емдеу әдістері, тіс ұлпасының девитализациясы
Ұлпа қабынуы туралы түсінік ұлпа қабынуын емдеу әдістері, тіс ұлпасының девитализациясы Родительское собрание на тему Воспитание самостоятельности
Родительское собрание на тему Воспитание самостоятельности Лекция №4. Профилактика туберкулеза. Организация медицинской помощи больным туберкулезом
Лекция №4. Профилактика туберкулеза. Организация медицинской помощи больным туберкулезом Предмет химии
Предмет химии Дигибридное скрещивание
Дигибридное скрещивание Моделирование систем. Управление рисками, системный анализ и моделирование. Лекции 11-12
Моделирование систем. Управление рисками, системный анализ и моделирование. Лекции 11-12 Презентация Нравственные качества, ОРКСЭ, урок №7, учебник Шемшуриной (Дрофа), 4 класс
Презентация Нравственные качества, ОРКСЭ, урок №7, учебник Шемшуриной (Дрофа), 4 класс Туркменистан - государство в Центральной Азии
Туркменистан - государство в Центральной Азии Россия при первых Романовых: перемены в государственном устройстве
Россия при первых Романовых: перемены в государственном устройстве Тайна имени Екатерина
Тайна имени Екатерина Будущая профессия – тренер по боксу
Будущая профессия – тренер по боксу Моя любимая и лучшая подруга
Моя любимая и лучшая подруга Обработка деталей на станках с программным управлением
Обработка деталей на станках с программным управлением Дидактическая игра-тренажёр Щенок и чудак
Дидактическая игра-тренажёр Щенок и чудак Выступление на педсовете Подготовка учащихся 9, 10, 11 классов к ГИА и ЕГЭ
Выступление на педсовете Подготовка учащихся 9, 10, 11 классов к ГИА и ЕГЭ Общие сведения о релейной защите
Общие сведения о релейной защите Вода.
Вода. ОСНОВНЫЕ ВИДЫ МУЗЫКАЛЬНЫХ ИГРУШЕК В РАЗНЫХ ВОЗРАСТНЫХ ГРУППАХ
ОСНОВНЫЕ ВИДЫ МУЗЫКАЛЬНЫХ ИГРУШЕК В РАЗНЫХ ВОЗРАСТНЫХ ГРУППАХ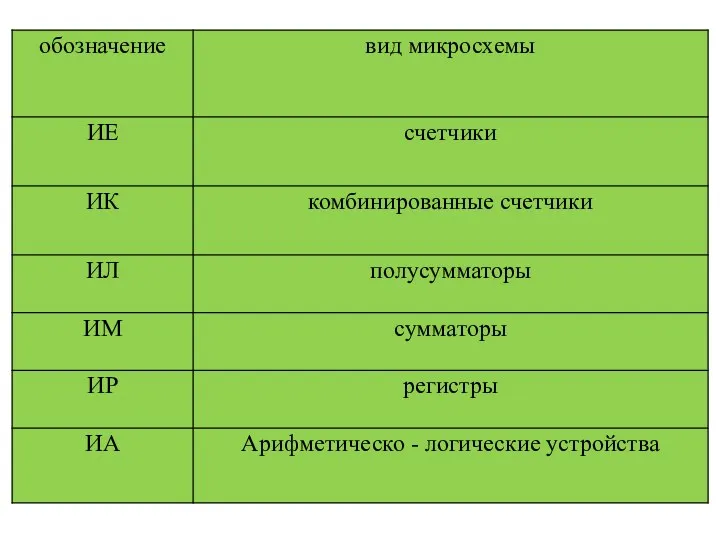 Характеристики интегральных схем (ИС)
Характеристики интегральных схем (ИС) Қан топтары. Қан құю әдісі
Қан топтары. Қан құю әдісі ГО ВПО Донецкий национальный университет экономики и торговли имени Михаила Туганбарановского. Кафедра международной экономики
ГО ВПО Донецкий национальный университет экономики и торговли имени Михаила Туганбарановского. Кафедра международной экономики Антропометрическое соответствие одежды
Антропометрическое соответствие одежды Истеъмолчиларга ичимлик суви ва оқова сув хизматларини кўрсатиш қоидалари
Истеъмолчиларга ичимлик суви ва оқова сув хизматларини кўрсатиш қоидалари Обновления в CSS3
Обновления в CSS3 презентация Листопад
презентация Листопад Организация оказания медицинской помощи детям Заягорбского района
Организация оказания медицинской помощи детям Заягорбского района Гигиена. Понятие. Этапы. Задачи
Гигиена. Понятие. Этапы. Задачи