- Доказательства эволюции

Содержание
- 2. Излюбленным аргументом противников Дарвина с давних пор была кажущаяся неспособность его теории объяснить происхождение камбалообразных рыб.
- 3. В ряде других случаев, напротив, филогенетические ряды достаточно полны для восстановления всей картины эволюции (китообразные, лошади,
- 4. Предки китов, общие с парнокопытными, постепенно приспосабливались к жизни в море, заполняя экологическую нишу, которая освободилась
- 5. В ходе эволюции предков человека за 2.6 млн лет (от других гоминин до рода Номо и
- 6. Австралопитеки (4—2 миллиона лет назад), объем мозга был около 400 см³, как у шимпанзе Человек умелый
- 7. Эволюция гоминин
- 8. «Микро-» и «макроэволюция» могут принципиально отличаться, если крупные изменения происходят скачкообразно. Но чаще макроэволюция - это
- 9. Пример: Переход от лопастеперых рыб к тетраподам растянулся примерно на 25 млн лет (385 - 360
- 10. Жабры Сердце Тело Жабры Сердце Тело Легкие Р ы б ы Т е т р а
- 11. Упрощенная схема перехода от лопастеперых рыб к первым тетраподам. Рисунок использовался на "обезьяньем процессе" в Пенсильвании
- 12. Из вердикта суда (судья Джонс, 2005, Пенсильвания) по поводу включения в курс местной школьной программы идеи
- 13. Костистые рыбы в кайнозое стали предпринимать новые "попытки" освоить сушу, порой весьма успешные. У этих современных
- 14. 4. Морфологические доказательства Если идея эволюции верна, мы должны наблюдать в живой природе многочисленные свидетельства "переделки"
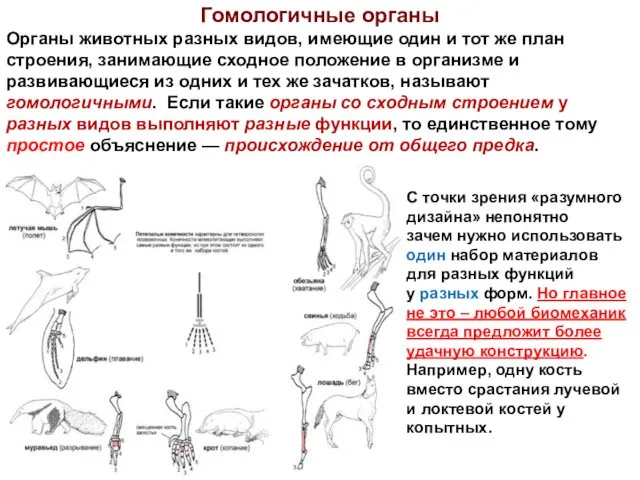
- 15. Гомологичные органы Органы животных разных видов, имеющие один и тот же план строения, занимающие сходное положение
- 17. Рудименты Органы, утратившие своё основное значение в процессе эволюции. Это также структуры, редуцированные и обладающие меньшими
- 18. Какапо – (новозеландский нелетающий) совиный попугай. К сожалению, он, похоже, не только забыл, как летать, но
- 19. Другие примеры рудиментов: У человека к рудиментам относятся: Хвостовые позвонки; волосяной покров туловища, Специальные мышцы которые
- 20. Червеобразный отросток слепой кишки (аппендикс). Многолетние наблюдения показали, что удаление аппендикса не оказывает значимого влияния на
- 21. Рудиментарные задние конечности питона
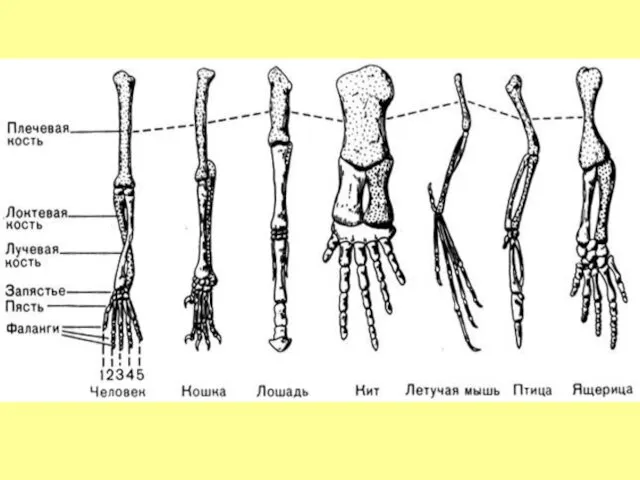
- 22. Рудиментарные кости тазового пояса гренландского кита
- 23. Атавизмы Появление у особи признаков, свойственных отдаленным предкам. Появление атавизмов объясняется тем, что гены, отвечающие за
- 24. Несовершенство строения организмов В ходе эволюции каждая новая конструкция не проектируется с нуля, а получается из
- 25. Расположение 4 –ой ветви блуждающего нерва у не имеющей шеи акулы вполне рационально. У жирафа, унаследовавшего
- 26. Желудочно-кишечный тракт млекопитающих пересекается с дыхательными путями, в результате мы не можем одновременно дышать и глотать,
- 27. Эволюция глаза у позвоночных животных Еще один пример несовершенства в строении организмов — сетчатка позвоночных и
- 28. У позвоночных есть целая серия адаптаций к такому «вывернутому» строению глаза (напр. отсутствует миелиновая оболочка аксонов,
- 29. Первичные фоторецепторы (даны красным) закладываются в ходе развития нервной трубки у позвоночных животных
- 30. Эволюция глаза у моллюсков. У головоногих (самый нижний рисунок) аналогичный по сложности глаз устроен вполне рационально
- 31. 5. Эмбриологические доказательства Свидетельства эволюции в индивидуальном развитии организмов 1. Эволюционируют (меняются) не взрослые организмы и
- 32. 2. Генетическая программа онтогенеза многоклеточных животных сама по себе содержит меньше информации, чем получающийся на ее
- 33. 3. Индивидуальное развитие многоклеточных организмов довольно часто повторяет отдельные этапы его эволюционной истории (биогенетический закон Мюллера-Геккеля
- 34. Даже очень непохожие друг на друга животные проходят одинаковые ранние стадии развития: зигота (оплодотворенное яйцо), бластула,
- 35. У всех позвоночных животных наблюдается сходство зародышей на ранних стадиях развития: форма тела, зачатки жаберных дуг,
- 36. Месячный эмбрион человека. Видны зачатки жабер и хвост. Разумеется, жаберные мешки зародышей млекопитающих в ходе развития
- 37. Эмбрион дельфина. Видны зачатки передних (f) и задних (h) конечностей. Из первых разовьются плавники, вторые исчезнут.
- 38. У сумчатых млекопитающих в ходе развития зародышей на короткое время возникает и потом рассасывается яичная скорлупа,
- 39. У взрослых камбалообразных рыб оба глаза находятся на одной стороне головы (обычно правой), а весь череп
- 40. «Эво-дево»(evolutionary development) : следы макроэволюции Легко представить себе изменчивость особей одного вида по размерам, форме, окраске
- 41. Генетикам давно известны мутации, вызывающие резкие перестройки плана строения организмов, такие как гомеозис (изменение органов) или
- 42. Но: крупные морфологические перестройки могут быть обусловлены не только генетическими, но и эпигенетическими механизмами, связанными с
- 43. При определенных обстоятельствах могут длительно поддерживаться целые популяции таких «монстров». Пример: льнянка (родственник львиного зева). К.Линнеем
- 44. Еще пример: пастушья сумка (родственник арабидопсиса). В природе встречаются популяции у которых 4 лепестка, характерных для
- 45. Важно отметить, что в обоих примерах никакой видимой большей адаптивности за этими эволюционными изменениями не стоит.
- 46. Отсюда возникло новое современное направление – эволюционная генетика развития, или «эво-дево» (evo-devo - сокращение от evolutionary
- 47. В заключительной главе «Происхождения видов» (по изданию 1939г., с.659) Ч. Дарвин пишет: «Но так как в
- 48. Пример «эво-дево»: развитие цветка у семейства орхидных - одного из 2-х самых больших семейств цветковых (орхидных
- 49. Ареал семейства орхидных (самое большое по числу видов семейство однодольных) - пример несомненно успешного таксона. Каждый
- 50. Цветок орхидей поражает своим разнообразием. Но за этим стоит строгая прагматичность: 60% орхидей опыляются только своим
- 51. 6. Молекулярно-генетические и биохимические доказательства Выяснение того факта, что ДНК постоянно мутирует является обоснованием теории эволюции
- 52. Эволюционная теория, в отличие от антиэволюционистов, может логически объяснить почему генетический код почти не меняется в
- 53. 2. Различия между геномами видов хорошо соответствуют независимо построенным филогенетическом деревьям и палеонтологической летописи. Оценка родства
- 54. 3. Несмотря на то, что белки сохраняют функции и после многих замен АК, и несмотря на
- 55. 4. Установление родства по сходству ДНК откалибровано на группах людей с детально известной и датированной родословной
- 56. Для человека, шимпанзе и гориллы по такому же совпадению нейтральных мутаций ДНК, – это ближайшие виды-родственники,
- 57. Биохимическое единство жизни Если не учитывать, что выражение «единство происхождения» конечно можно заменить на «единство творения»,
- 58. 2-я хромосома человека После слияния двух хромосом остаются характерные следы: остатки теломер и рудимертарная центромера. У
- 59. Эндогенные ретровирусы Эндогенные ретровирусы (ЭР)- следы древних вирусных инфекций в ДНК (1% ДНК человека, все вирусы
- 60. Псевдогены Это неработающие, "молчащие" гены, которые возникают в результате мутаций, выводящих нормальные "рабочие" гены из строя.
- 61. У других млекопитающих (например, у крысы) GULO является не псевдогеном, а работающим геном, и поэтому крысам
- 62. Пост-транскрипционная регуляция работы генов. "Несовершенство дизайна организмов" обнаруживается и на молекулярном уровне. Для отключения работы генов
- 63. При этом ферменты «нарочно» вставляют в нуклеотидную последовательность мРНК ошибки при транскрипции (например, преждевременные стоп-кодоны), чтобы
- 65. Скачать презентацию

Излюбленным аргументом противников Дарвина с давних пор была кажущаяся неспособность его теории
Излюбленным аргументом противников Дарвина с давних пор была кажущаяся неспособность его теории

В ряде других случаев, напротив, филогенетические ряды достаточно полны для восстановления
В ряде других случаев, напротив, филогенетические ряды достаточно полны для восстановления

Предки китов, общие с парнокопытными, постепенно приспосабливались к жизни в море,
Предки китов, общие с парнокопытными, постепенно приспосабливались к жизни в море,
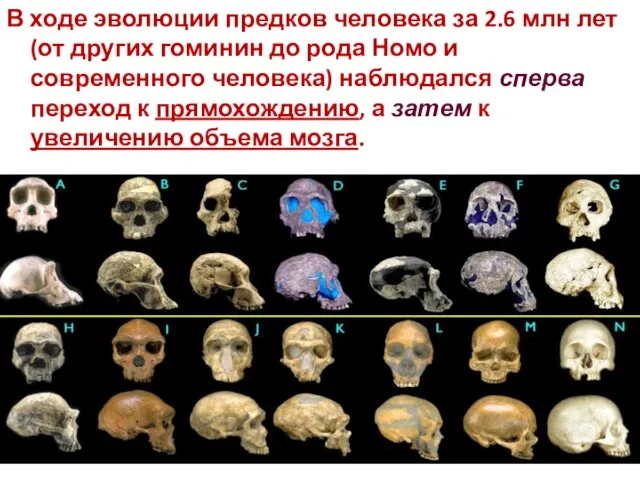
В ходе эволюции предков человека за 2.6 млн лет (от других
В ходе эволюции предков человека за 2.6 млн лет (от других

Австралопитеки (4—2 миллиона лет назад), объем мозга был около 400 см³,
Австралопитеки (4—2 миллиона лет назад), объем мозга был около 400 см³,
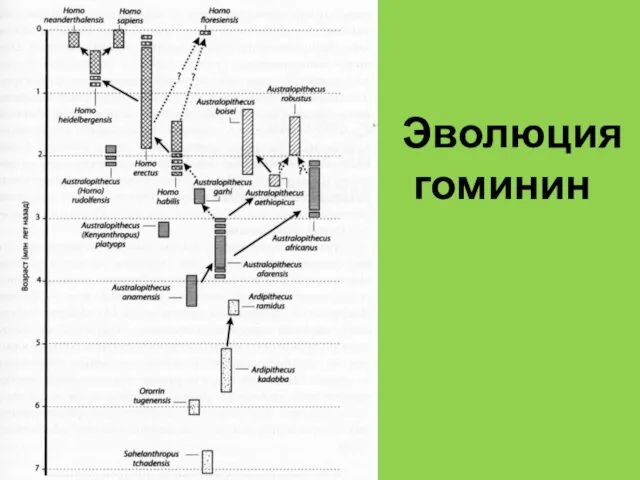
Эволюция
гоминин
Эволюция
гоминин

«Микро-» и «макроэволюция»
могут принципиально отличаться, если крупные изменения происходят скачкообразно.
Но
«Микро-» и «макроэволюция» могут принципиально отличаться, если крупные изменения происходят скачкообразно. Но

Пример:
Переход от лопастеперых рыб к тетраподам растянулся примерно на 25
Пример:
Переход от лопастеперых рыб к тетраподам растянулся примерно на 25
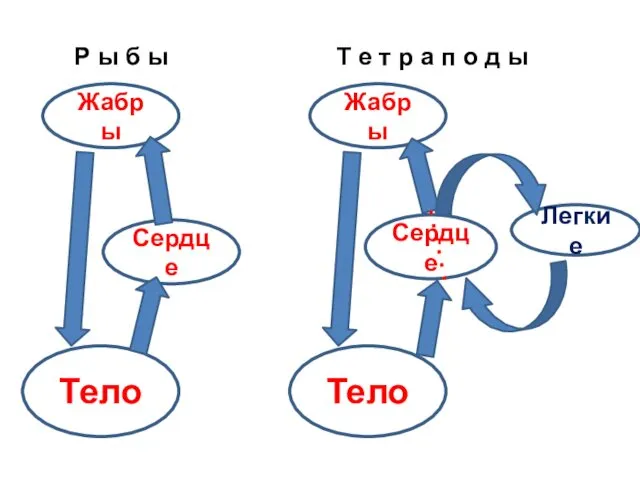
Жабры
Сердце
Тело
Жабры
Сердце
Тело
Легкие
Р ы б ы
Т е т р а п о
Жабры
Сердце
Тело
Жабры
Сердце
Тело
Легкие
Р ы б ы
Т е т р а п о
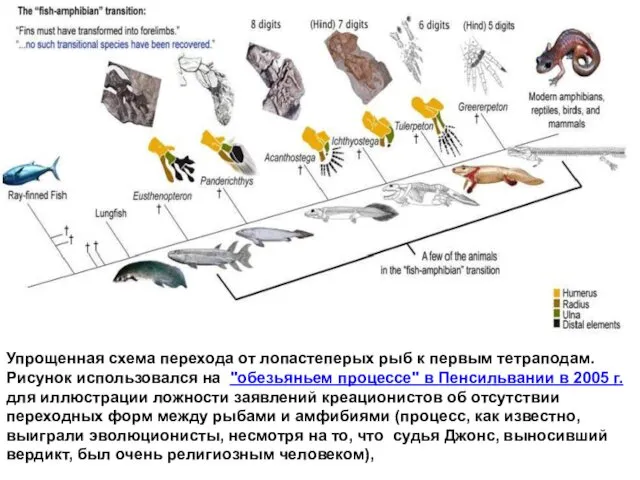
Упрощенная схема перехода от лопастеперых рыб к первым тетраподам. Рисунок использовался
Упрощенная схема перехода от лопастеперых рыб к первым тетраподам. Рисунок использовался

Из вердикта суда (судья Джонс, 2005, Пенсильвания) по поводу включения в
Из вердикта суда (судья Джонс, 2005, Пенсильвания) по поводу включения в

Костистые рыбы в кайнозое стали предпринимать новые "попытки" освоить сушу, порой
Костистые рыбы в кайнозое стали предпринимать новые "попытки" освоить сушу, порой

4. Морфологические доказательства
Если идея эволюции верна, мы должны наблюдать в живой природе
4. Морфологические доказательства
Если идея эволюции верна, мы должны наблюдать в живой природе
Гомологичные органы
Органы животных разных видов, имеющие один и тот же план строения, занимающие
Гомологичные органы
Органы животных разных видов, имеющие один и тот же план строения, занимающие

Рудименты
Органы, утратившие своё основное значение в процессе эволюции. Это также структуры,
Рудименты
Органы, утратившие своё основное значение в процессе эволюции. Это также структуры,

Какапо – (новозеландский нелетающий) совиный попугай. К сожалению, он, похоже, не
Какапо – (новозеландский нелетающий) совиный попугай. К сожалению, он, похоже, не

Другие примеры рудиментов:
У человека к рудиментам относятся:
Хвостовые позвонки; волосяной покров туловища,
Специальные
Другие примеры рудиментов:
У человека к рудиментам относятся:
Хвостовые позвонки; волосяной покров туловища,
Специальные

Червеобразный отросток слепой кишки (аппендикс).
Многолетние наблюдения показали, что удаление аппендикса
Червеобразный отросток слепой кишки (аппендикс).
Многолетние наблюдения показали, что удаление аппендикса

Рудиментарные задние конечности питона
Рудиментарные задние конечности питона
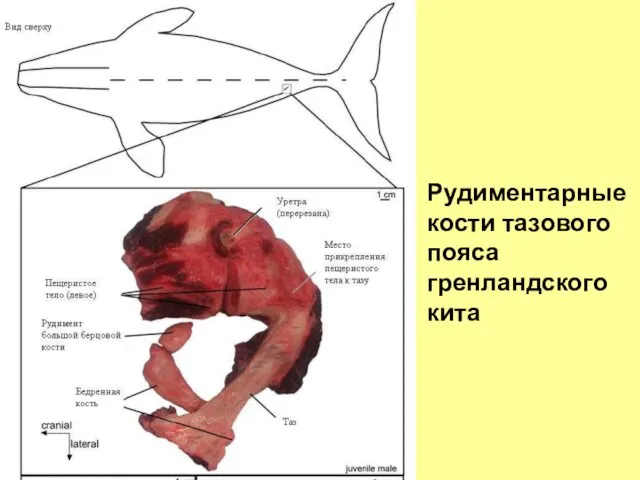
Рудиментарные
кости тазового
пояса
гренландского кита
Рудиментарные
кости тазового
пояса
гренландского кита

Атавизмы
Появление у особи признаков, свойственных отдаленным предкам. Появление атавизмов объясняется тем,
Атавизмы
Появление у особи признаков, свойственных отдаленным предкам. Появление атавизмов объясняется тем,

Несовершенство строения организмов
В ходе эволюции каждая новая конструкция не проектируется с
Несовершенство строения организмов
В ходе эволюции каждая новая конструкция не проектируется с
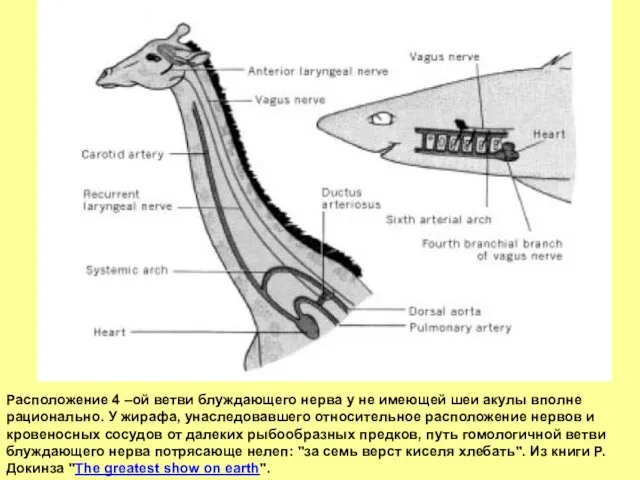
Расположение 4 –ой ветви блуждающего нерва у не имеющей шеи акулы
Расположение 4 –ой ветви блуждающего нерва у не имеющей шеи акулы

Желудочно-кишечный тракт млекопитающих пересекается с дыхательными путями, в результате мы не
Желудочно-кишечный тракт млекопитающих пересекается с дыхательными путями, в результате мы не

Эволюция глаза у позвоночных животных
Еще один пример несовершенства в строении организмов — сетчатка
Эволюция глаза у позвоночных животных
Еще один пример несовершенства в строении организмов — сетчатка
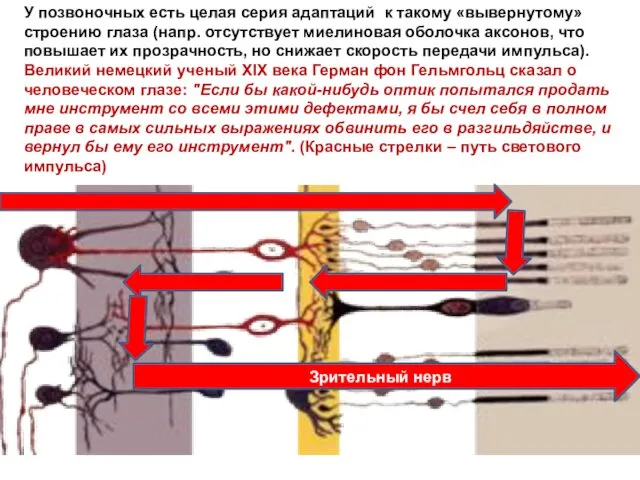
У позвоночных есть целая серия адаптаций к такому «вывернутому» строению глаза
У позвоночных есть целая серия адаптаций к такому «вывернутому» строению глаза
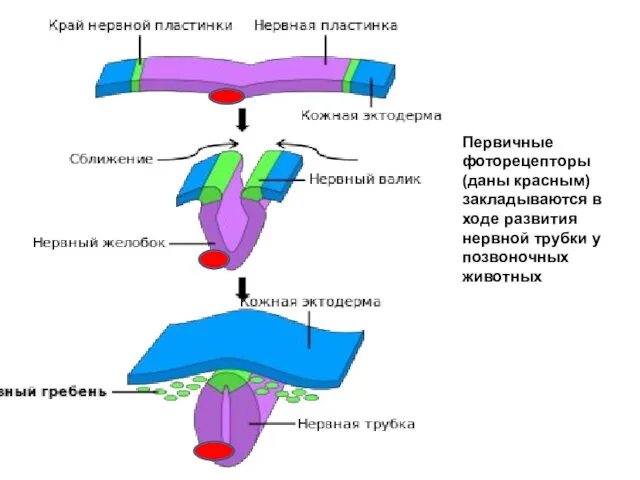
Первичные
фоторецепторы
(даны красным)
закладываются в
ходе развития нервной трубки у
позвоночных животных
Первичные
фоторецепторы
(даны красным)
закладываются в
ходе развития нервной трубки у
позвоночных животных
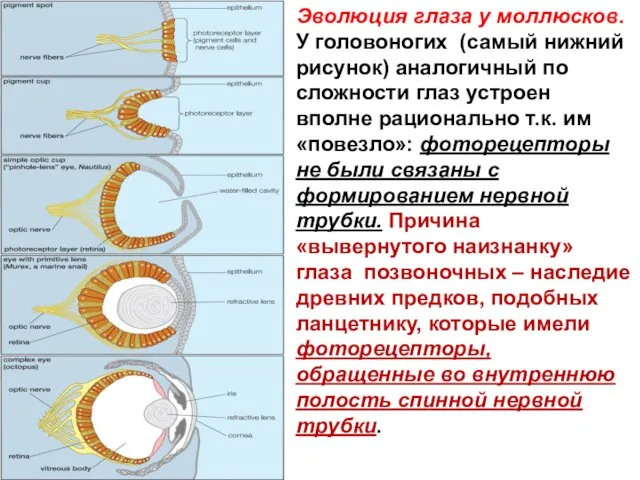
Эволюция глаза у моллюсков.
У головоногих (самый нижний рисунок) аналогичный по
Эволюция глаза у моллюсков.
У головоногих (самый нижний рисунок) аналогичный по

5. Эмбриологические
доказательства
Свидетельства эволюции в индивидуальном развитии организмов
1. Эволюционируют (меняются) не взрослые
5. Эмбриологические
доказательства
Свидетельства эволюции в индивидуальном развитии организмов
1. Эволюционируют (меняются) не взрослые

2. Генетическая программа онтогенеза многоклеточных животных сама по себе содержит меньше
2. Генетическая программа онтогенеза многоклеточных животных сама по себе содержит меньше

3. Индивидуальное развитие многоклеточных организмов довольно часто повторяет отдельные этапы его
3. Индивидуальное развитие многоклеточных организмов довольно часто повторяет отдельные этапы его

Даже очень непохожие друг на друга животные проходят одинаковые ранние стадии
Даже очень непохожие друг на друга животные проходят одинаковые ранние стадии

У всех позвоночных животных наблюдается сходство зародышей на ранних стадиях развития:
У всех позвоночных животных наблюдается сходство зародышей на ранних стадиях развития:
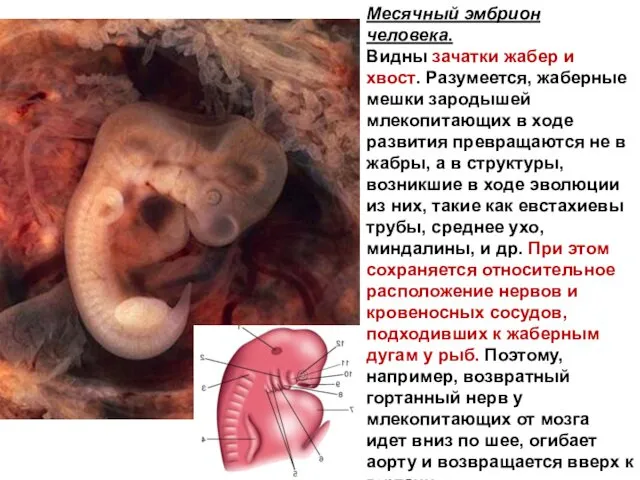
Месячный эмбрион
человека.
Видны зачатки жабер и хвост. Разумеется, жаберные мешки зародышей
Месячный эмбрион
человека.
Видны зачатки жабер и хвост. Разумеется, жаберные мешки зародышей
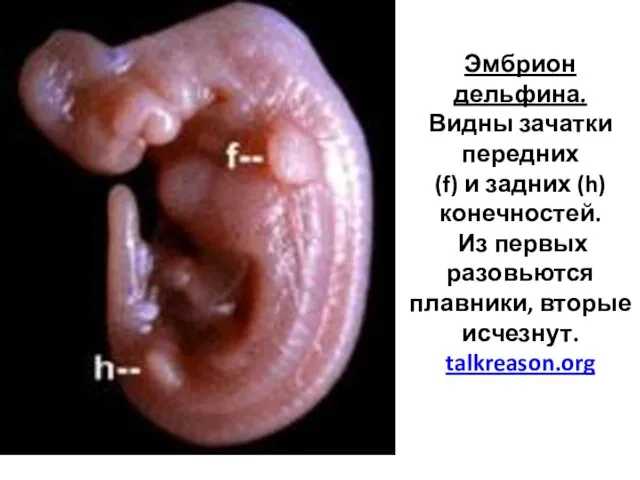
Эмбрион дельфина.
Видны зачатки передних (f) и задних (h)
конечностей.
Из первых разовьются плавники, вторые исчезнут.
Эмбрион дельфина.
Видны зачатки передних (f) и задних (h)
конечностей.
Из первых разовьются плавники, вторые исчезнут.

У сумчатых млекопитающих в ходе развития зародышей на короткое время возникает
У сумчатых млекопитающих в ходе развития зародышей на короткое время возникает

У взрослых камбалообразных рыб оба глаза находятся на одной стороне головы
У взрослых камбалообразных рыб оба глаза находятся на одной стороне головы

«Эво-дево»(evolutionary development)
: следы макроэволюции
Легко представить себе изменчивость особей одного вида по
«Эво-дево»(evolutionary development)
: следы макроэволюции
Легко представить себе изменчивость особей одного вида по

Генетикам давно известны мутации, вызывающие резкие перестройки плана строения организмов, такие
Генетикам давно известны мутации, вызывающие резкие перестройки плана строения организмов, такие

Но: крупные морфологические перестройки могут быть обусловлены не только генетическими, но
Но: крупные морфологические перестройки могут быть обусловлены не только генетическими, но

При определенных обстоятельствах могут длительно
поддерживаться целые популяции таких «монстров».
Пример: льнянка
При определенных обстоятельствах могут длительно
поддерживаться целые популяции таких «монстров».
Пример: льнянка

Еще пример: пастушья сумка (родственник арабидопсиса). В природе встречаются популяции у
Еще пример: пастушья сумка (родственник арабидопсиса). В природе встречаются популяции у

Важно отметить, что в обоих примерах никакой видимой
большей адаптивности за
Важно отметить, что в обоих примерах никакой видимой
большей адаптивности за

Отсюда возникло новое современное направление – эволюционная генетика развития, или «эво-дево»
Отсюда возникло новое современное направление – эволюционная генетика развития, или «эво-дево»

В заключительной главе «Происхождения видов» (по изданию 1939г., с.659) Ч. Дарвин
В заключительной главе «Происхождения видов» (по изданию 1939г., с.659) Ч. Дарвин

Пример «эво-дево»: развитие цветка у семейства орхидных -
одного из 2-х
Пример «эво-дево»: развитие цветка у семейства орхидных -
одного из 2-х
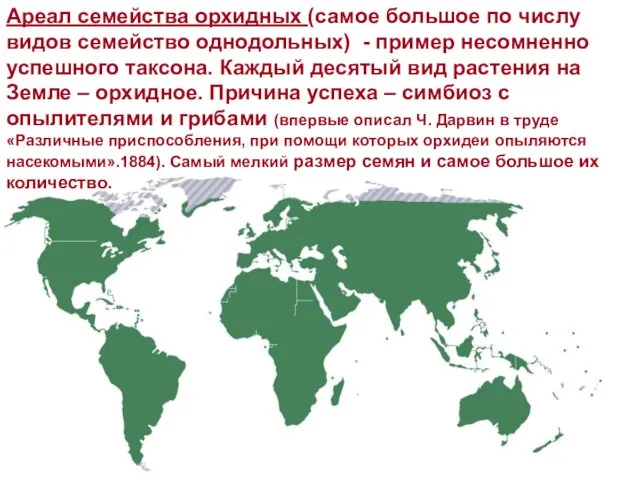
Ареал семейства орхидных (самое большое по числу видов семейство однодольных) -
Ареал семейства орхидных (самое большое по числу видов семейство однодольных) -

Цветок орхидей поражает своим разнообразием. Но за этим стоит строгая прагматичность:
Цветок орхидей поражает своим разнообразием. Но за этим стоит строгая прагматичность:

6. Молекулярно-генетические
и биохимические доказательства
Выяснение того факта, что ДНК постоянно мутирует является
6. Молекулярно-генетические
и биохимические доказательства
Выяснение того факта, что ДНК постоянно мутирует является

Эволюционная теория, в отличие от антиэволюционистов, может логически объяснить почему генетический
Эволюционная теория, в отличие от антиэволюционистов, может логически объяснить почему генетический

2. Различия между геномами видов хорошо соответствуют независимо построенным филогенетическом деревьям
2. Различия между геномами видов хорошо соответствуют независимо построенным филогенетическом деревьям

3. Несмотря на то, что белки сохраняют функции и после многих
3. Несмотря на то, что белки сохраняют функции и после многих

4. Установление родства по сходству ДНК откалибровано на группах людей с
4. Установление родства по сходству ДНК откалибровано на группах людей с

Для человека, шимпанзе и гориллы по такому же совпадению нейтральных мутаций
Для человека, шимпанзе и гориллы по такому же совпадению нейтральных мутаций

Биохимическое единство жизни
Если не учитывать, что выражение «единство происхождения» конечно
Биохимическое единство жизни
Если не учитывать, что выражение «единство происхождения» конечно
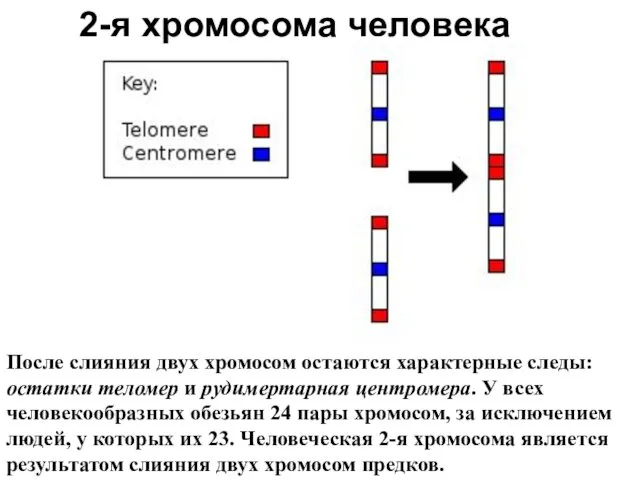
2-я хромосома человека
После слияния двух хромосом остаются характерные следы: остатки теломер
2-я хромосома человека
После слияния двух хромосом остаются характерные следы: остатки теломер

Эндогенные ретровирусы
Эндогенные ретровирусы (ЭР)- следы древних вирусных инфекций в ДНК (1%
Эндогенные ретровирусы
Эндогенные ретровирусы (ЭР)- следы древних вирусных инфекций в ДНК (1%

Псевдогены
Это неработающие, "молчащие" гены, которые возникают в результате мутаций, выводящих нормальные
Псевдогены
Это неработающие, "молчащие" гены, которые возникают в результате мутаций, выводящих нормальные

У других млекопитающих (например, у крысы) GULO является не псевдогеном, а работающим геном,
У других млекопитающих (например, у крысы) GULO является не псевдогеном, а работающим геном,

Пост-транскрипционная регуляция работы генов.
"Несовершенство дизайна организмов" обнаруживается и на молекулярном уровне.
Пост-транскрипционная регуляция работы генов.
"Несовершенство дизайна организмов" обнаруживается и на молекулярном уровне.

При этом ферменты «нарочно» вставляют в нуклеотидную последовательность мРНК ошибки при
При этом ферменты «нарочно» вставляют в нуклеотидную последовательность мРНК ошибки при
 Атомные электростанции (АЭС)
Атомные электростанции (АЭС) Коррекция длины и радиуса инструмента
Коррекция длины и радиуса инструмента Кризис ценностных приоритетов и общественное мнение, роль СМИ в формировании негативного имиджа семьи
Кризис ценностных приоритетов и общественное мнение, роль СМИ в формировании негативного имиджа семьи A Lesson About JOBS (Part 2)
A Lesson About JOBS (Part 2) Методическая разработка мероприятия по теме Русская гармонь в народном творчестве Кубани
Методическая разработка мероприятия по теме Русская гармонь в народном творчестве Кубани Организация учебного процесса в высшем учебном заведении
Организация учебного процесса в высшем учебном заведении Олимпиадный эксперимент – в школьный практикум. Часть 2
Олимпиадный эксперимент – в школьный практикум. Часть 2 Теоретические основы построения теней. Лекция 12
Теоретические основы построения теней. Лекция 12 Удельная теплота плавления
Удельная теплота плавления Могущество католической церкви. Еретики
Могущество католической церкви. Еретики Coachella – is a three-day festival which takes place in California every April
Coachella – is a three-day festival which takes place in California every April Актуальні питання історії та культури України
Актуальні питання історії та культури України Органика
Органика Вторичный амилоидоз
Вторичный амилоидоз Урок по технологии 4 класс
Урок по технологии 4 класс Цитология – наука о клетке. Современные методы исследования
Цитология – наука о клетке. Современные методы исследования О воспитанности
О воспитанности Ваша личная бизнес-модель профессионального развития
Ваша личная бизнес-модель профессионального развития Техническая эксплуатация железнодорожного подвижного состава
Техническая эксплуатация железнодорожного подвижного состава Закрытие месяца в Бухгалтерии предприятия, ред. 3.0
Закрытие месяца в Бухгалтерии предприятия, ред. 3.0 Моё портфолио
Моё портфолио Организация ЭВМ и систем
Организация ЭВМ и систем Согласные шипящие звуки
Согласные шипящие звуки Ауспалы егіс
Ауспалы егіс История аэропорта Красноярск-Северный
История аэропорта Красноярск-Северный Otpusk_i_starenie_stali
Otpusk_i_starenie_stali Границы применимости классической механики. Кинематика
Границы применимости классической механики. Кинематика Предметно-протранственная среда в ДОУ, как фактор формирования личности ребёнка дошкольного возраста
Предметно-протранственная среда в ДОУ, как фактор формирования личности ребёнка дошкольного возраста