- Физиология возбудимых тканей

Содержание
- 2. Физиологический покой, физиологическая активность Биологические системы (организмы, органы, ткани и клетки) – могут находиться в двух
- 3. Раздражимость Раздражимость – это способность живых организмов, клеток, тканей и органов реагировать на внешнее воздействие изменением
- 4. Возбудимость Возбудимость - это способность организма, органа, ткани или клетки быстро отвечать на раздражение специфической реакцией
- 5. Раздражимость, возбудимость, возбуждение Раздражимость и возбудимость характеризуют одно и то же свойство биологической системы – способность
- 6. Возбуждение и торможение Изменение мембранного потенциала клетки является основным признаком возбуждения всех возбудимых структур, независимо от
- 7. Открытие биоэлектрических явлений Адамсон, 1751 г. получил первые данные о существовании «животного электричества» при изучении природы
- 8. Изучение биоэлектрических явлений Г. Гельмгольц (Германия) в 1850 г. измерил скорость распространения нервного импульса по нерву
- 9. Изучение биопотенциалов русскими учеными В. Ю. Чаговец в 1896 предложил первую ионную теорию биоэлектрических явлений и
- 10. Разработка ионно-мембранной теории Г. Бишоп, Дж. Эрлангер и Г. Гассер (США) в 30-40-х гг. XX в
- 11. Цитоплазматическая мембрана – ПЛАЗМОЛЕММА
- 12. Плазмолемма, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку
- 13. История открытия О наличии пограничной мембраны между клетками и окружающей их средой предполагали задолго до появления
- 14. История открытия Овертон предположил, что клеточная мембрана содержит большое количество липидов; вещества растворяются в ней, проходя
- 15. История открытия Изучение натяжения и гибкости пограничного слоя клетки позволило заключить, что в клеточной мембране содержится
- 16. История открытия
- 17. История открытия В 1972 г. Сингер и Николсон предположили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы,
- 18. Химический состав Наружная клеточная мембрана, по современным представлениям, имеет толщину около 10 нм (10-9 м), и
- 19. Химический состав Липидный бислой представлен преимущественно: фосфолипидами (глицерофосфатидами), сфингомиелинами и из стероидных липидов — холестерином (холестеролом)
- 20. Химический состав Свойство молекулы иметь в своем составе как гидрофильные, так и гидрофобные группы, называется амфифильностью.
- 21. Химический состав
- 22. Химический состав Атомы углерода, образующие двойные связи, не могут свободно вращаться, поэтому они занимают фиксированную позицию
- 23. Химический состав
- 24. Химический состав
- 25. Химический состав Вязкость мембранных липидов во многом определяет свойства и поведение мембран. На вязкость липидов в
- 26. Химический состав Липиды различных типов встраиваются в мембрану далеко не случайным образом. Каждая сторона бислоя имеет
- 27. Химический состав Большая часть белков и липидов, входящих в состав мембраны, способны перемещаться, главным образом в
- 28. Химический состав В двойной слой липидов на различную глубину погружены мембранные белки, составляющие более 50% массы
- 29. Химический состав По своему расположению относительно липидного бислоя мембранные белки разделяются на две основные группы –
- 30. Химический состав
- 31. Цитолемма
- 32. Химический состав Углеводные участки гликолипидов и гликопротеинов придают поверхности клетки отрицательный заряд и образуют основу надмембранного
- 33. Химический состав
- 34. Белки мембраны Интегральные белки встроены в липидный бислой, по строению – это глобулярные белки. Это белки
- 35. Функции мембран СТРУКТУРНАЯ ЗАЩИТНАЯ ФЕРМЕНТАТИВНАЯ СОЕДИНИТЕЛЬНАЯ ИЛИ АДГЕЗИВНАЯ (обуславливает существование многоклеточных организмов) РЕЦЕПТОРНАЯ АНТИГЕННАЯ ЭЛЕКТРОГЕННАЯ ТРАНСПОРТНАЯ
- 36. Плазматическая мембрана
- 37. Схема строения плазматической мембраны
- 38. Асимметрия Трансмембранная асимметрия означает, что разные половины бислоя имеют разный состав. Установлено, что интегральные мембранные белки
- 39. Функции Благодаря барьерной функции мембраны, в клетке создается гетерогенная физико-химическая среда, и на разных сторонах мембраны
- 40. Функции Осмотическая функция мембраны связана с регуляцией водного обмена клетки. Благодаря структурной функции поддерживается основа мембран
- 41. Функции Биосинтетическая функция связана с синтезами различных веществ. Рецепторно-регуляторная функция определяется наличием в мембране хемо-, фото-
- 42. Транспорт Основная функция любой биологической мембраны состоит в создании барьера с селективной проницаемостью. Так, выполняя транспортную
- 43. Транспорт Тонкий гидрофобный центральный слой в мембране является очень эффективным барьером для неорганических ионов, но в
- 44. Транспорт Органические ионы или органические хелатные комплексы, содержащие неорганические ионы, могут растворяться в углеводородной области бислоя
- 45. Транспорт Транспорт большинства растворимых молекул через биологические мембраны опосредуется переносчиками или канальными белками. Каналы облегчают транспорт
- 46. Транспорт При облегченной диффузии переносчик, функционирующий в клеточной мембране, на одной стороне мембраны соединяется с молекулой
- 47. Транспорт В отличие от облегченной диффузии активный транспорт – это перемещение веществ против градиентов концентрации; вещества
- 48. Транспорт Осмосом называют прохождение воды через избирательно проницаемую мембрану, в частности через клеточную мембрану. В случае
- 49. Транспорт Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их
- 50. Транспорт Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае внутриклеточные продукты, заключенные
- 51. Транспорт
- 52. Plasmalemma
- 53. Ионные каналы Это белковые макромолекулы, погруженные в липидный бислой плазматической мембраны (трансмембранные белки), образующие заполненные водой
- 54. Механизмы прохождения веществ через клеточную мембрану
- 55. Схема строения ионного канала
- 56. Потенциалзависимый и хемоуправляемый натриевые каналы
- 57. Три разных функциональных состояния потенциал-зависимого натриевого канал
- 58. Воротный механизм
- 59. Натрий-калиевый насос
- 60. ВЛИЯНИЕ СТРЕССОВЫХ ВОЗДЕЙСТВИЙ НА СТРУКТУРУ И ФУНКЦИИ МЕМБРАН (на примере мембран растений)
- 61. Стрессовые воздействия Неспецифические реакции клетки на стрессовые воздействия в значительной степени определяются изменениями мембранного аппарата. Стабильность
- 62. Стрессовые воздействия Мембраны первыми подвергаются действию стрессовых факторов. Они представляют собой мишени первичного воздействия и первую
- 63. Стрессовые воздействия Повышается проницаемость мембран, происходит деполяризация мембранного потенциала плазмалеммы, рН (кислотность) цитоплазмы сдвигается в кислую
- 64. Стрессовые воздействия Нарастает скорость процессов гидролиза, и тормозится синтез белка. Сдвиги в функциональной активности мембран сопровождаются
- 65. Стрессовые воздействия Структурные перестройки в мембранах в значительной мере касаются липидов. При действии стрессора могут происходить
- 66. Стрессовые воздействия Регуляторную роль в клеточном метаболизме играют и белки мембран. Они осуществляют регуляцию конформационных изменений
- 67. Стрессовые воздействия При значительной деструкции клеточных компонентов в результате стрессового воздействия поток кальция из апопласта в
- 68. Стрессовые воздействия Причины возрастания проницаемости мембран разнообразны. Их связывают с изменением отношения Н+/Ca2+ в мембранах. Стабилизатором
- 69. Стрессовые воздействия Таким образом, в устойчивости растений к неблагоприятным воздействиям фундаментальную роль играют клеточные мембраны, интегрирующие
- 70. РОЛЬ КЛЕТОЧНЫХ МЕМБРАН В СТАРЕНИИ КЛЕТОК
- 71. Роль мембран в старении клеток «Мембранная гипотеза старения» был разработана в 1976-1977 гг. и опубликована впервые
- 72. Роль мембран в старении клеток Увеличение внутриклеточного K+ помогает достичь состояния возбуждения на мембранах старых клеток,
- 73. Роль мембран в старении клеток Во внутриклеточной среде повышенной вязкости скорости синтеза РНК и белка должны
- 75. Скачать презентацию
Физиологический покой, физиологическая активность
Биологические системы (организмы, органы, ткани и клетки) – могут находиться
Физиологический покой, физиологическая активность
Биологические системы (организмы, органы, ткани и клетки) – могут находиться

Раздражимость
Раздражимость – это способность живых организмов, клеток, тканей и органов реагировать на внешнее
Раздражимость
Раздражимость – это способность живых организмов, клеток, тканей и органов реагировать на внешнее

Возбудимость
Возбудимость - это способность организма, органа, ткани или клетки быстро отвечать на раздражение
Возбудимость
Возбудимость - это способность организма, органа, ткани или клетки быстро отвечать на раздражение

Раздражимость, возбудимость, возбуждение
Раздражимость и возбудимость характеризуют одно и то же свойство биологической системы
Раздражимость, возбудимость, возбуждение
Раздражимость и возбудимость характеризуют одно и то же свойство биологической системы

Возбуждение и торможение
Изменение мембранного потенциала клетки является основным признаком возбуждения всех возбудимых структур,
Возбуждение и торможение
Изменение мембранного потенциала клетки является основным признаком возбуждения всех возбудимых структур,

Открытие биоэлектрических явлений
Адамсон, 1751 г. получил первые данные о существовании «животного электричества» при
Открытие биоэлектрических явлений
Адамсон, 1751 г. получил первые данные о существовании «животного электричества» при

Изучение биоэлектрических явлений
Г. Гельмгольц (Германия) в 1850 г. измерил скорость распространения нервного импульса
Изучение биоэлектрических явлений
Г. Гельмгольц (Германия) в 1850 г. измерил скорость распространения нервного импульса

Изучение биопотенциалов русскими учеными
В. Ю. Чаговец в 1896 предложил первую ионную теорию биоэлектрических
Изучение биопотенциалов русскими учеными
В. Ю. Чаговец в 1896 предложил первую ионную теорию биоэлектрических

Разработка ионно-мембранной теории
Г. Бишоп, Дж. Эрлангер и Г. Гассер (США) в 30-40-х гг.
Разработка ионно-мембранной теории
Г. Бишоп, Дж. Эрлангер и Г. Гассер (США) в 30-40-х гг.

Цитоплазматическая мембрана – ПЛАЗМОЛЕММА
Цитоплазматическая мембрана – ПЛАЗМОЛЕММА

Плазмолемма, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не
Плазмолемма, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не

История открытия
О наличии пограничной мембраны между клетками и окружающей их средой предполагали задолго
История открытия
О наличии пограничной мембраны между клетками и окружающей их средой предполагали задолго

История открытия
Овертон предположил, что клеточная мембрана содержит большое количество липидов; вещества растворяются в
История открытия
Овертон предположил, что клеточная мембрана содержит большое количество липидов; вещества растворяются в

История открытия
Изучение натяжения и гибкости пограничного слоя клетки позволило заключить, что в клеточной
История открытия
Изучение натяжения и гибкости пограничного слоя клетки позволило заключить, что в клеточной

История открытия
История открытия

История открытия
В 1972 г. Сингер и Николсон предположили жидкостно-мозаичную модель мембраны, согласно которой
История открытия
В 1972 г. Сингер и Николсон предположили жидкостно-мозаичную модель мембраны, согласно которой

Химический состав
Наружная клеточная мембрана, по современным представлениям, имеет толщину около 10 нм
Химический состав
Наружная клеточная мембрана, по современным представлениям, имеет толщину около 10 нм

Химический состав
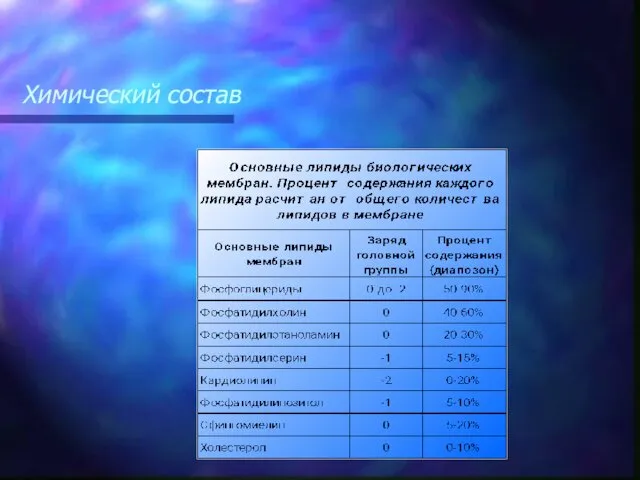
Липидный бислой представлен преимущественно:
фосфолипидами (глицерофосфатидами),
сфингомиелинами
и из стероидных
Химический состав
Липидный бислой представлен преимущественно:
фосфолипидами (глицерофосфатидами),
сфингомиелинами
и из стероидных

Химический состав
Свойство молекулы иметь в своем составе как гидрофильные, так и гидрофобные
Химический состав
Свойство молекулы иметь в своем составе как гидрофильные, так и гидрофобные

Химический состав
Химический состав
Химический состав
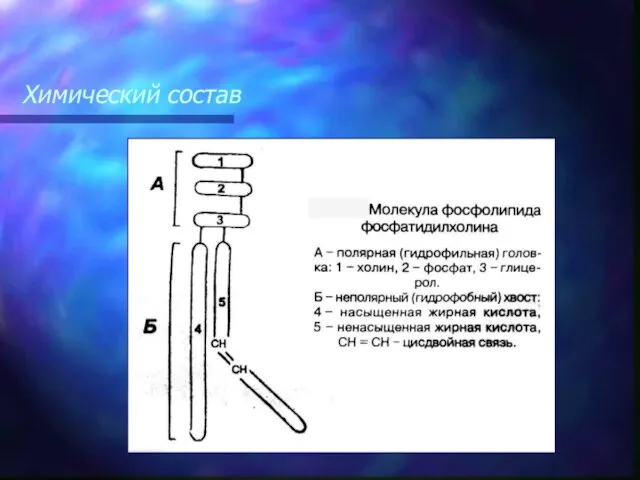
Атомы углерода, образующие двойные связи, не могут свободно вращаться, поэтому они
Химический состав
Атомы углерода, образующие двойные связи, не могут свободно вращаться, поэтому они

Химический состав
Химический состав
Химический состав
Химический состав

Химический состав
Вязкость мембранных липидов во многом определяет свойства и поведение мембран.
На
Химический состав
Вязкость мембранных липидов во многом определяет свойства и поведение мембран.
На

Химический состав
Липиды различных типов встраиваются в мембрану далеко не случайным образом.
Каждая
Химический состав
Липиды различных типов встраиваются в мембрану далеко не случайным образом.
Каждая

Химический состав
Большая часть белков и липидов, входящих в состав мембраны, способны перемещаться,
Химический состав
Большая часть белков и липидов, входящих в состав мембраны, способны перемещаться,

Химический состав
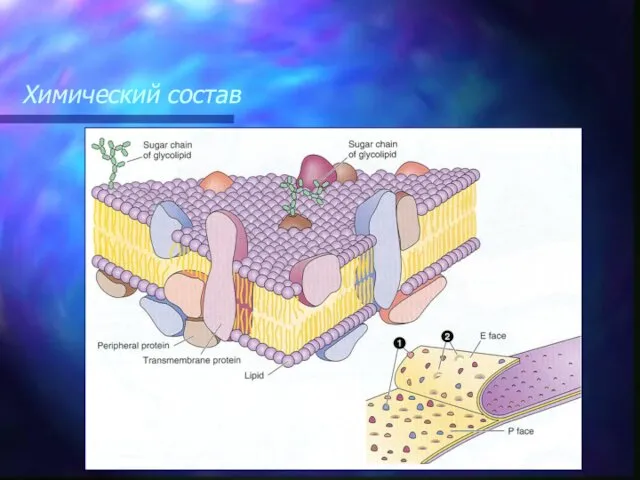
В двойной слой липидов на различную глубину погружены мембранные белки, составляющие
Химический состав
В двойной слой липидов на различную глубину погружены мембранные белки, составляющие

Химический состав
По своему расположению относительно липидного бислоя мембранные белки разделяются на две
Химический состав
По своему расположению относительно липидного бислоя мембранные белки разделяются на две

Химический состав
Химический состав
Цитолемма
Цитолемма
Химический состав
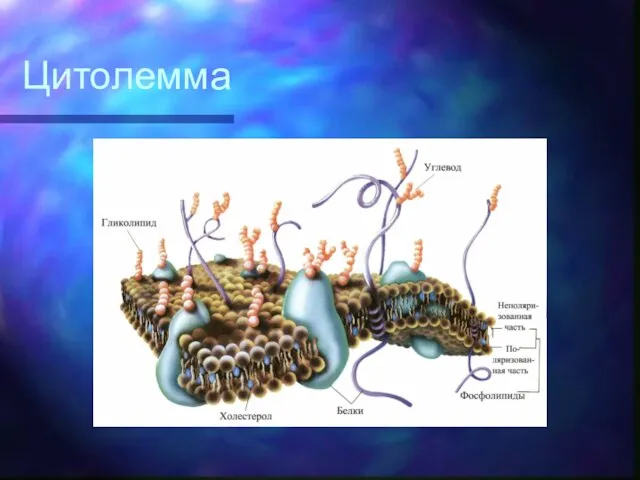
Углеводные участки гликолипидов и гликопротеинов придают поверхности клетки отрицательный заряд и
Химический состав
Углеводные участки гликолипидов и гликопротеинов придают поверхности клетки отрицательный заряд и

Химический состав
Химический состав
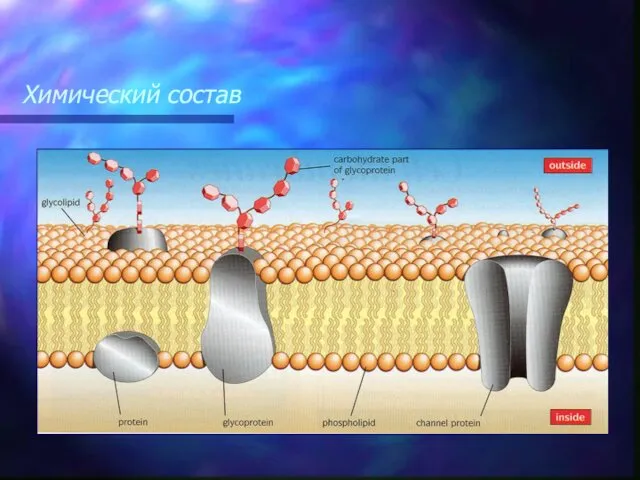
Белки мембраны
Интегральные белки встроены в липидный бислой, по строению – это глобулярные белки.
Белки мембраны
Интегральные белки встроены в липидный бислой, по строению – это глобулярные белки.

Функции мембран
СТРУКТУРНАЯ
ЗАЩИТНАЯ
ФЕРМЕНТАТИВНАЯ
СОЕДИНИТЕЛЬНАЯ ИЛИ АДГЕЗИВНАЯ (обуславливает существование многоклеточных организмов)
РЕЦЕПТОРНАЯ
АНТИГЕННАЯ
ЭЛЕКТРОГЕННАЯ
ТРАНСПОРТНАЯ
Функции мембран
СТРУКТУРНАЯ
ЗАЩИТНАЯ
ФЕРМЕНТАТИВНАЯ
СОЕДИНИТЕЛЬНАЯ ИЛИ АДГЕЗИВНАЯ (обуславливает существование многоклеточных организмов)
РЕЦЕПТОРНАЯ
АНТИГЕННАЯ
ЭЛЕКТРОГЕННАЯ
ТРАНСПОРТНАЯ

Плазматическая мембрана
Плазматическая мембрана
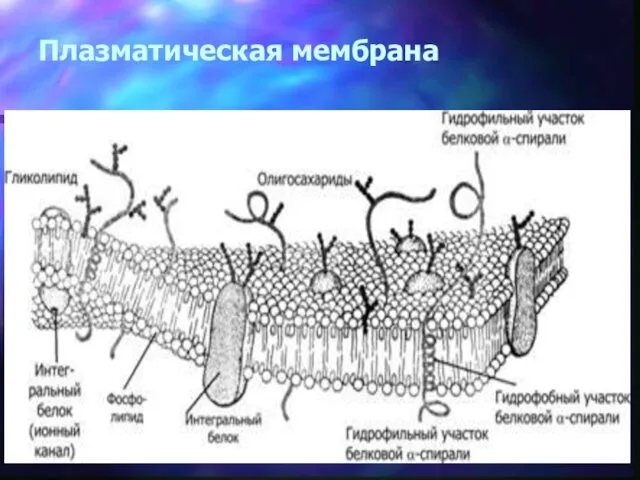
Схема строения плазматической мембраны
Схема строения плазматической мембраны
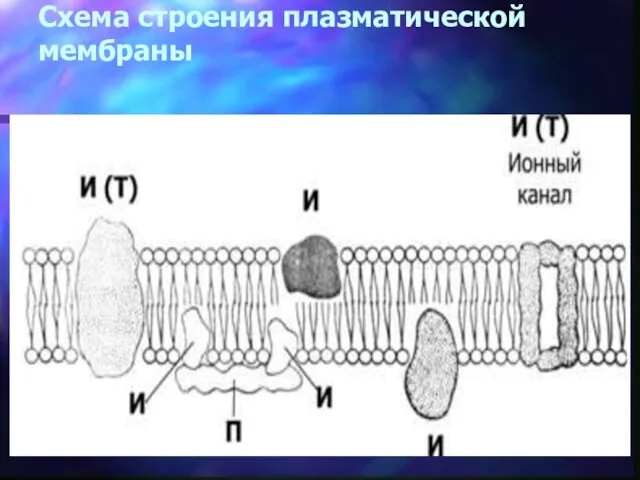
Асимметрия
Трансмембранная асимметрия означает, что разные половины бислоя имеют разный состав. Установлено, что
Асимметрия
Трансмембранная асимметрия означает, что разные половины бислоя имеют разный состав. Установлено, что

Функции
Благодаря барьерной функции мембраны, в клетке создается гетерогенная физико-химическая среда, и на разных
Функции
Благодаря барьерной функции мембраны, в клетке создается гетерогенная физико-химическая среда, и на разных

Функции
Осмотическая функция мембраны связана с регуляцией водного обмена клетки. Благодаря структурной функции поддерживается
Функции
Осмотическая функция мембраны связана с регуляцией водного обмена клетки. Благодаря структурной функции поддерживается

Функции
Биосинтетическая функция связана с синтезами различных веществ.
Рецепторно-регуляторная функция определяется наличием в мембране
Функции
Биосинтетическая функция связана с синтезами различных веществ.
Рецепторно-регуляторная функция определяется наличием в мембране

Транспорт
Основная функция любой биологической мембраны состоит в создании барьера с селективной проницаемостью. Так,
Транспорт
Основная функция любой биологической мембраны состоит в создании барьера с селективной проницаемостью. Так,

Транспорт
Тонкий гидрофобный центральный слой в мембране является очень эффективным барьером для неорганических ионов,
Транспорт
Тонкий гидрофобный центральный слой в мембране является очень эффективным барьером для неорганических ионов,

Транспорт
Органические ионы или органические хелатные комплексы, содержащие неорганические ионы, могут растворяться в углеводородной
Транспорт
Органические ионы или органические хелатные комплексы, содержащие неорганические ионы, могут растворяться в углеводородной

Транспорт
Транспорт большинства растворимых молекул через биологические мембраны опосредуется переносчиками или канальными белками. Каналы
Транспорт
Транспорт большинства растворимых молекул через биологические мембраны опосредуется переносчиками или канальными белками. Каналы

Транспорт
При облегченной диффузии переносчик, функционирующий в клеточной мембране, на одной стороне мембраны соединяется
Транспорт
При облегченной диффузии переносчик, функционирующий в клеточной мембране, на одной стороне мембраны соединяется

Транспорт
В отличие от облегченной диффузии активный транспорт – это перемещение веществ против градиентов
Транспорт
В отличие от облегченной диффузии активный транспорт – это перемещение веществ против градиентов

Транспорт
Осмосом называют прохождение воды через избирательно проницаемую мембрану, в частности через клеточную мембрану.
Транспорт
Осмосом называют прохождение воды через избирательно проницаемую мембрану, в частности через клеточную мембрану.

Транспорт
Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и
Транспорт
Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и

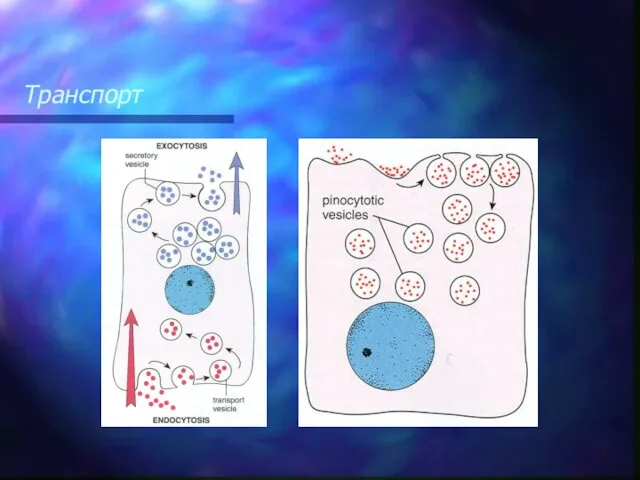
Транспорт
Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае внутриклеточные
Транспорт
Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае внутриклеточные

Транспорт
Транспорт
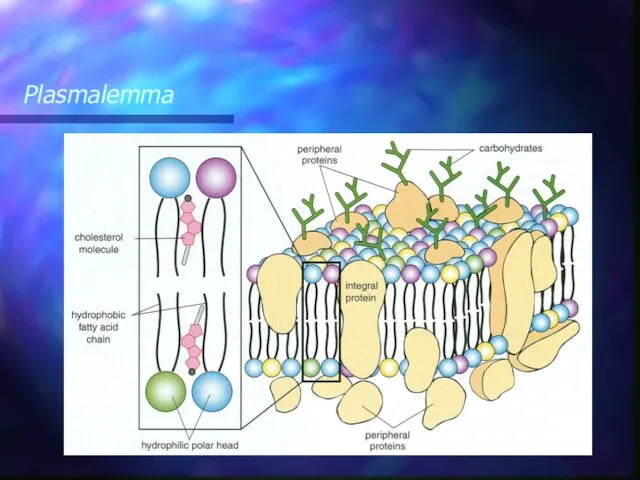
Plasmalemma
Plasmalemma
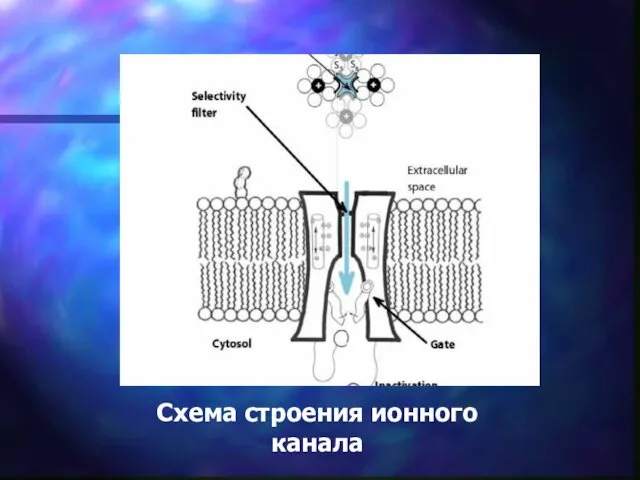
Ионные каналы
Это белковые макромолекулы, погруженные в липидный бислой плазматической мембраны (трансмембранные белки), образующие
Ионные каналы
Это белковые макромолекулы, погруженные в липидный бислой плазматической мембраны (трансмембранные белки), образующие

Механизмы прохождения веществ через клеточную мембрану
Механизмы прохождения веществ через клеточную мембрану
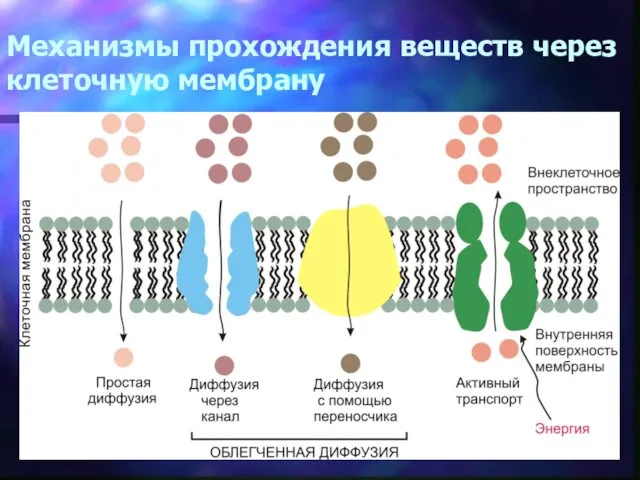
Схема строения ионного канала
Схема строения ионного канала
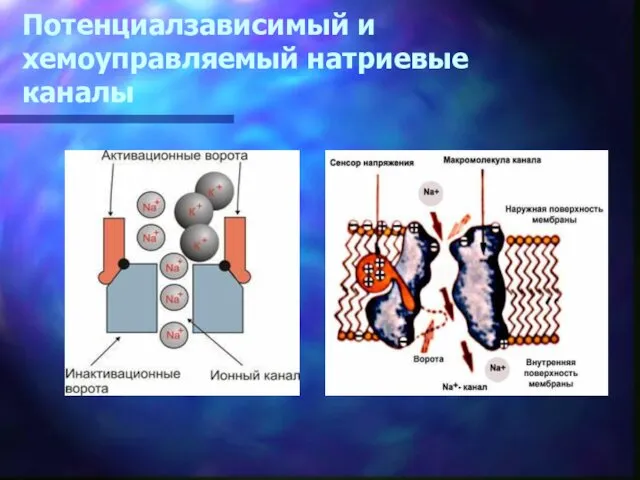
Потенциалзависимый и хемоуправляемый натриевые каналы
Потенциалзависимый и хемоуправляемый натриевые каналы
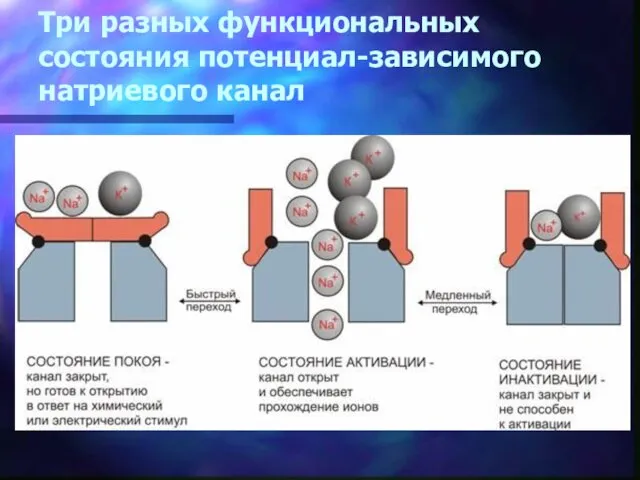
Три разных функциональных состояния потенциал-зависимого натриевого канал
Три разных функциональных состояния потенциал-зависимого натриевого канал
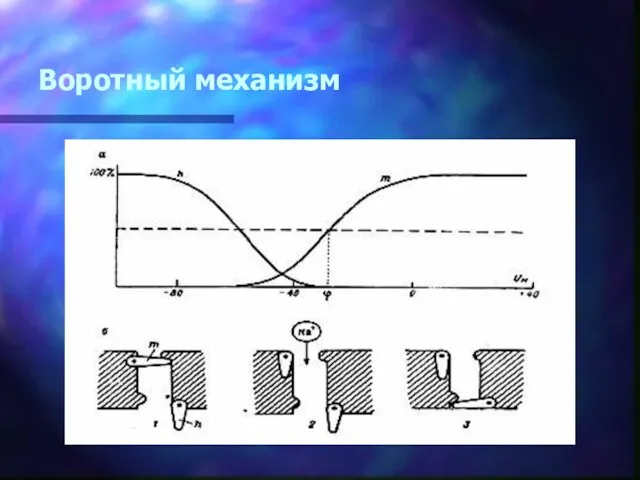
Воротный механизм
Воротный механизм
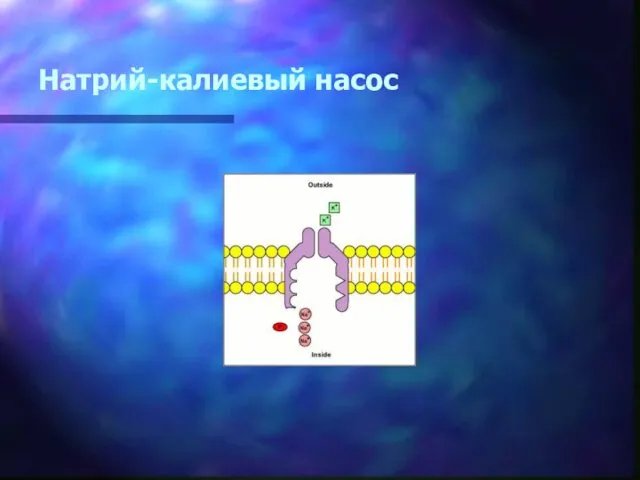
Натрий-калиевый насос
Натрий-калиевый насос
ВЛИЯНИЕ СТРЕССОВЫХ ВОЗДЕЙСТВИЙ НА СТРУКТУРУ И ФУНКЦИИ МЕМБРАН
(на примере мембран растений)
ВЛИЯНИЕ СТРЕССОВЫХ ВОЗДЕЙСТВИЙ НА СТРУКТУРУ И ФУНКЦИИ МЕМБРАН
(на примере мембран растений)

Стрессовые
воздействия
Неспецифические реакции клетки на стрессовые воздействия в значительной степени определяются изменениями мембранного
Стрессовые
воздействия
Неспецифические реакции клетки на стрессовые воздействия в значительной степени определяются изменениями мембранного

Стрессовые
воздействия
Мембраны первыми подвергаются действию стрессовых факторов. Они представляют собой мишени первичного воздействия
Стрессовые
воздействия
Мембраны первыми подвергаются действию стрессовых факторов. Они представляют собой мишени первичного воздействия

Стрессовые
воздействия
Повышается проницаемость мембран, происходит деполяризация мембранного потенциала плазмалеммы, рН (кислотность) цитоплазмы сдвигается
Стрессовые
воздействия
Повышается проницаемость мембран, происходит деполяризация мембранного потенциала плазмалеммы, рН (кислотность) цитоплазмы сдвигается

Стрессовые
воздействия
Нарастает скорость процессов гидролиза, и тормозится синтез белка.
Сдвиги в функциональной активности
Стрессовые
воздействия
Нарастает скорость процессов гидролиза, и тормозится синтез белка.
Сдвиги в функциональной активности

Стрессовые
воздействия
Структурные перестройки в мембранах в значительной мере касаются липидов. При действии стрессора
Стрессовые
воздействия
Структурные перестройки в мембранах в значительной мере касаются липидов. При действии стрессора

Стрессовые
воздействия
Регуляторную роль в клеточном метаболизме играют и белки мембран. Они осуществляют регуляцию
Стрессовые
воздействия
Регуляторную роль в клеточном метаболизме играют и белки мембран. Они осуществляют регуляцию

Стрессовые
воздействия
При значительной деструкции клеточных компонентов в результате стрессового воздействия поток кальция из
Стрессовые
воздействия
При значительной деструкции клеточных компонентов в результате стрессового воздействия поток кальция из

Стрессовые
воздействия
Причины возрастания проницаемости мембран разнообразны. Их связывают с изменением отношения Н+/Ca2+ в
Стрессовые
воздействия
Причины возрастания проницаемости мембран разнообразны. Их связывают с изменением отношения Н+/Ca2+ в

Стрессовые
воздействия
Таким образом, в устойчивости растений к неблагоприятным воздействиям фундаментальную роль играют клеточные
Стрессовые
воздействия
Таким образом, в устойчивости растений к неблагоприятным воздействиям фундаментальную роль играют клеточные

РОЛЬ КЛЕТОЧНЫХ МЕМБРАН В СТАРЕНИИ КЛЕТОК
РОЛЬ КЛЕТОЧНЫХ МЕМБРАН В СТАРЕНИИ КЛЕТОК

Роль мембран
в старении клеток
«Мембранная гипотеза старения» был разработана в 1976-1977 гг. и опубликована
Роль мембран
в старении клеток
«Мембранная гипотеза старения» был разработана в 1976-1977 гг. и опубликована

Роль мембран
в старении клеток
Увеличение внутриклеточного K+ помогает достичь состояния возбуждения на мембранах старых
Роль мембран
в старении клеток
Увеличение внутриклеточного K+ помогает достичь состояния возбуждения на мембранах старых

Роль мембран
в старении клеток
Во внутриклеточной среде повышенной вязкости скорости синтеза РНК и белка
Роль мембран
в старении клеток
Во внутриклеточной среде повышенной вязкости скорости синтеза РНК и белка

 Расчет дуговой электросталеплавильной печи емкостью 5т
Расчет дуговой электросталеплавильной печи емкостью 5т Отборочный тур муниципальной игры Юный пешеход – друг дорог!
Отборочный тур муниципальной игры Юный пешеход – друг дорог! Проектирование реакционного узла для жидкофазных реакторов. Лекция 5
Проектирование реакционного узла для жидкофазных реакторов. Лекция 5 Мера центральной тенденции
Мера центральной тенденции Бизнес - Информатика
Бизнес - Информатика Методика обучения письму и письменной речи в школе
Методика обучения письму и письменной речи в школе Операційний менеджмент як різновид функціонального менеджменту
Операційний менеджмент як різновид функціонального менеджменту Практические вопросы применения 223-ФЗ. Аукцион
Практические вопросы применения 223-ФЗ. Аукцион 20231031_prezentatsiya_po_biologii_na_temu_metody_izucheniya_zhivoy_prirody-izmerenie.5_klass
20231031_prezentatsiya_po_biologii_na_temu_metody_izucheniya_zhivoy_prirody-izmerenie.5_klass Исследование органов пищеварения
Исследование органов пищеварения Класицизм
Класицизм Проект по географии 7 класс Остров Робинзона Крузо: миф или реальность
Проект по географии 7 класс Остров Робинзона Крузо: миф или реальность Конструирование печатного узла и печатной платы. Лекция №10 (1)
Конструирование печатного узла и печатной платы. Лекция №10 (1) Методы производства многослойных печатных плат
Методы производства многослойных печатных плат С днем рождения, Александр! Фотоальбом
С днем рождения, Александр! Фотоальбом Человечество- единый биологический вид
Человечество- единый биологический вид Колонны. Расчет центрально-сжатых колонн по материалам
Колонны. Расчет центрально-сжатых колонн по материалам Презентация .Родительское собраниеРоль книги в жизни младших школьников
Презентация .Родительское собраниеРоль книги в жизни младших школьников Mitosis and the Cell Cycle. Cell reproduction
Mitosis and the Cell Cycle. Cell reproduction Презентация Рука развивает мозг
Презентация Рука развивает мозг Особенности РowerРoint-2007. (Занятие 5, часть 21)
Особенности РowerРoint-2007. (Занятие 5, часть 21) Второе начало термодинамики. Энтропия
Второе начало термодинамики. Энтропия Проект В мире сказки
Проект В мире сказки Нелинейные модели. Подходы к имитационному моделированию
Нелинейные модели. Подходы к имитационному моделированию Отделка женского платья и детской одежды. Обработка воланов
Отделка женского платья и детской одежды. Обработка воланов Как помочь ребенку стать внимательным
Как помочь ребенку стать внимательным Religion. How does religion effect the modern life?
Religion. How does religion effect the modern life? Материал для работы над проектом
Материал для работы над проектом