Глутаминовая кислота и ГАМК – главные медиаторы ЦНС: синтез, типы рецепторов, инактивация презентация
- Глутаминовая кислота и ГАМК – главные медиаторы ЦНС: синтез, типы рецепторов, инактивация

Содержание
- 2. Медиаторы-аминокислоты являются главными медиаторами ЦНС. Глутаминовая кислота (Glu) – главный возбуждающий меди-атор (около 40% всех нейронов);
- 3. Глутаминовая кислота и ГАМК – конкуренты и партнёры. При нарушении баланса Glu и ГАМК: синдром дефицита
- 4. Глутаминовая кислота. Пищевая аминокислота – входит в состав белков пищи и белков нашего тела; самая распростра-
- 5. R Другой («быстрый») путь синтеза, характерный для нейронов: взаимные превращения Glu и очень сходной пищевой а/к
- 6. R Другой («быстрый») путь синтеза, характерный для нейронов: взаимные превращения Glu и очень сходной пищевой а/к
- 7. Всех тканях организма (в т.ч. в мозге) очень много Glu. В связи с этим долгое время
- 8. Всех тканях организма (в т.ч. в мозге) очень много Glu. В связи с этим долгое время
- 9. Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые клетки-рецепторы языка («вкус белка»). Существуют
- 10. Рецепторы глутамата. В настоящее время выделяют три типа ионотропных и не менее трех типов метаботропных рецепторов
- 11. NMDA-рецепторы (агонист N-метил-D-аспартат) АМРА-рецепторы (агонист амино-гидрокси-метил- изоксазол-пропионовая кислота) Каинатные рецепторы (агонист каиновая кислота) Рецепторы разных типов
- 12. Наиболее изучены NMDA- рецепторы. Каждый такой рец-р состоит из 4-х белковых молекул; в открытом положении он
- 13. Наиболее очевидный способ удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА), находящихся на той же
- 14. Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать ион Mg2+ («магниевая пробка»). В
- 15. Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную ДНК, активацией генов не-NMDA-рецепторов, синтезом
- 16. Пусть мы учим крысу прыгать на полку в ответ на звонок (иначе она получает удар электрическим
- 17. Бонус: еще 2 схемы, где показаны упомянутые выше пластические изме-нения – выбивание Mg2+- пробок, фосфорилирование АМРА-рецепторов
- 18. Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты, «проверенные временем». Антагонисты Glu. Основные
- 19. Инактивация Glu: Из синаптич. щели Glu ● переносится в глиальные клетки, где превращается в глутамин (Gln)
- 20. ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается не столько торможение ЦНС, сколько
- 21. ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается не столько торможение ЦНС, сколько
- 22. Ноотропы, являющиеся производными ГАМК, улучшают выработку энергии нейронами. Но есть и другие группы ноотропов, улучшающие состояние
- 23. Рецепторы ГАМК. Выделяют два основных типа рецепторов ГАМК: (GABA – gamma-aminobutyric acid) ГАМКА – ионотропные, из
- 24. Антагонисты ГАМК: вызывают судороги. Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа; токсин североамериканского травянистого
- 25. Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины. Барбитураты: открыты более 100 лет назад в день Св. Варвары. продолжительное
- 26. Барбитураты: открыты более 100 лет назад в день Св. Варвары. продолжительное общее тормозящ. действие; используются для
- 27. Инактивация ГАМК. В значительной мере сходна с инактивацией Glu: обратное всасывание ГАМК в пресинаптическое окончание либо
- 28. Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы в ЦНС и, наряду с
- 29. Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов ГАМК), снижающих активность центров заднего
- 30. Тестирование транквилизаторов на эксперимен-тальных животных Тестирование потен-циальных лекарств на животных – обязательный этап перед их внедрением
- 31. Сон и наркоз. Мозговые центры, регулирующие смену сна и бодрствования – одни из самых древних структур
- 32. Сон и наркоз. Мозговые центры, регулирующие смену сна и бодрствования – одни из самых древних структур
- 33. Эпилепсия. Около 0.5% населения (предрасположены ~2%). В большинстве случаев в мозге больного есть зона постоянной активации
- 34. Эпилепсия. Около 0.5% населения (предрасположены ~2%). В большинстве случаев в мозге больного есть зона постоянной активации
- 35. 5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50% взрослых; наиболее эффективны психотерапия +
- 36. 5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50% взрослых; наиболее эффективны психотерапия +
- 38. Скачать презентацию

Медиаторы-аминокислоты являются главными медиаторами ЦНС.
Глутаминовая кислота (Glu) – главный возбуждающий меди-атор
Медиаторы-аминокислоты являются главными медиаторами ЦНС.
Глутаминовая кислота (Glu) – главный возбуждающий меди-атор

Глутаминовая кислота и ГАМК –
конкуренты и партнёры.
При нарушении баланса Glu
Глутаминовая кислота и ГАМК –
конкуренты и партнёры.
При нарушении баланса Glu
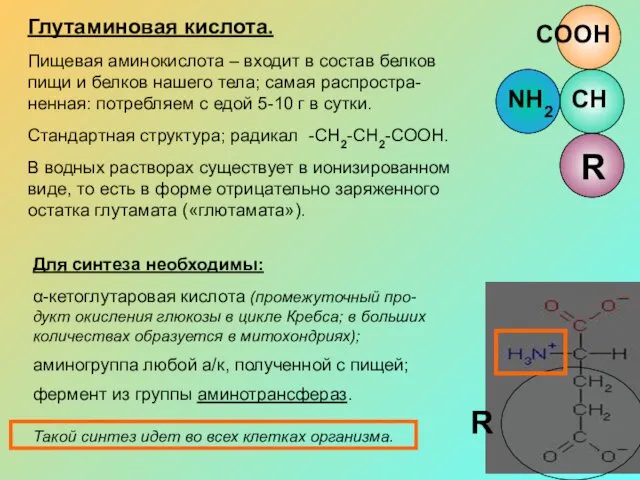
Глутаминовая кислота.
Пищевая аминокислота – входит в состав белков пищи и белков
Глутаминовая кислота.
Пищевая аминокислота – входит в состав белков пищи и белков
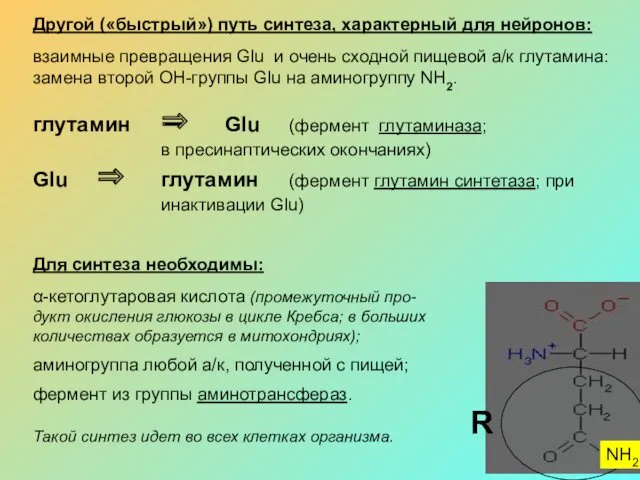
R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень

R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
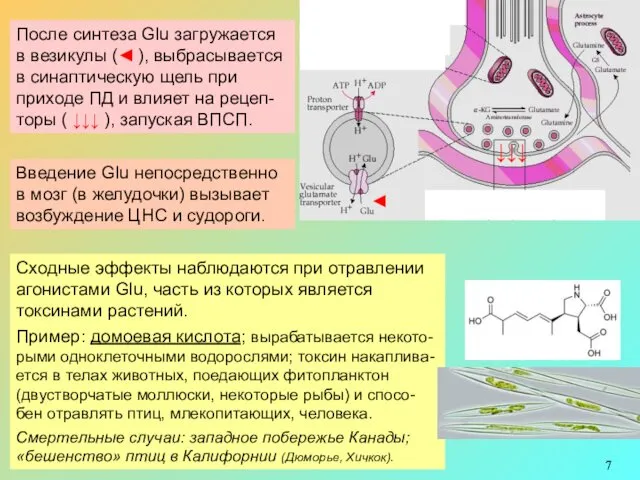
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
Всех тканях организма (в т.ч. в мозге) очень много Glu. В

Всех тканях организма (в т.ч. в мозге) очень много Glu. В
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
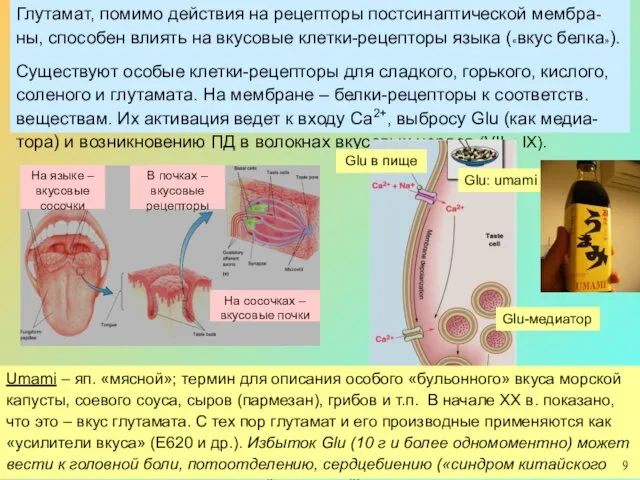
Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые
Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые

Рецепторы глутамата.
В настоящее время выделяют три типа ионотропных и не менее
Рецепторы глутамата.
В настоящее время выделяют три типа ионотропных и не менее

NMDA-рецепторы (агонист N-метил-D-аспартат)
АМРА-рецепторы (агонист амино-гидрокси-метил-
изоксазол-пропионовая кислота)
Каинатные рецепторы (агонист каиновая кислота)
Рецепторы разных
NMDA-рецепторы (агонист N-метил-D-аспартат)
АМРА-рецепторы (агонист амино-гидрокси-метил-
изоксазол-пропионовая кислота)
Каинатные рецепторы (агонист каиновая кислота)
Рецепторы разных
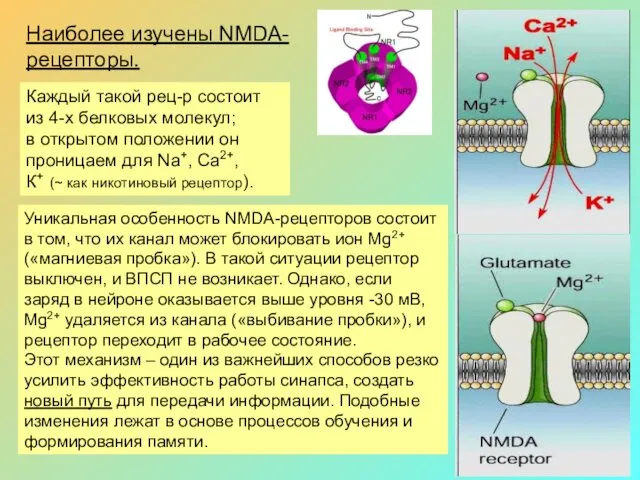
Наиболее изучены NMDA-
рецепторы.
Каждый такой рец-р состоит из 4-х белковых молекул;
в
Наиболее изучены NMDA-
рецепторы.
Каждый такой рец-р состоит из 4-х белковых молекул;
в
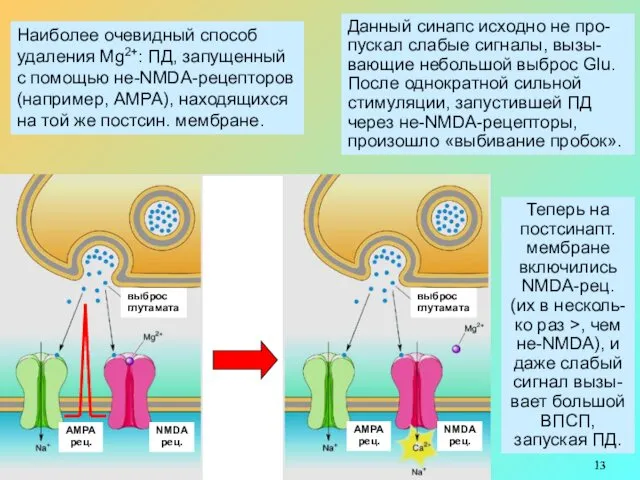
Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА),
Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА),

Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать
Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать
Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную
Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную
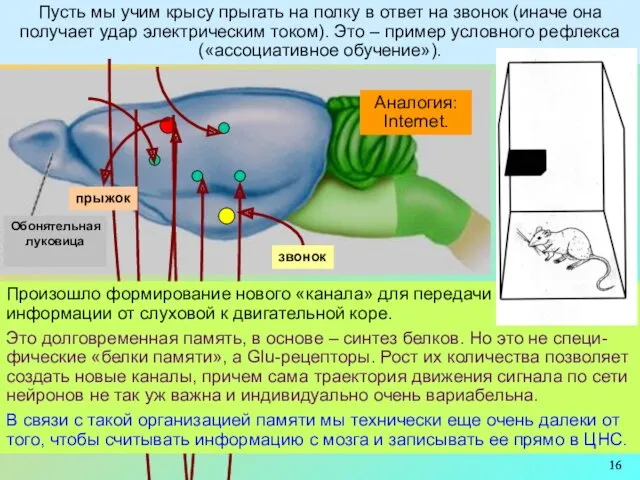
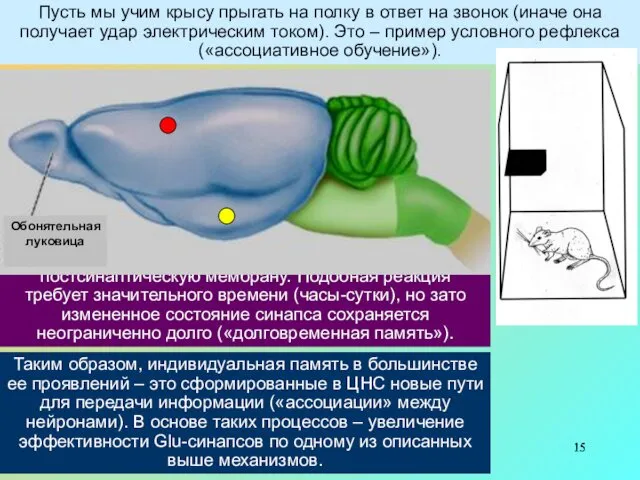
Пусть мы учим крысу прыгать на полку в ответ на звонок
Пусть мы учим крысу прыгать на полку в ответ на звонок
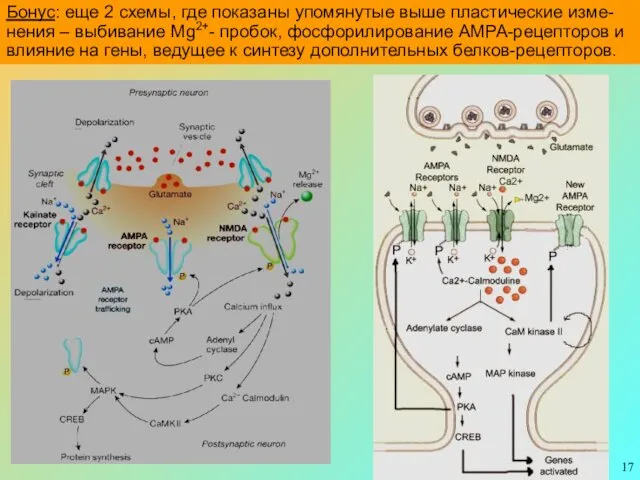
Бонус: еще 2 схемы, где показаны упомянутые выше пластические изме-нения –
Бонус: еще 2 схемы, где показаны упомянутые выше пластические изме-нения –

Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты,
Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты,
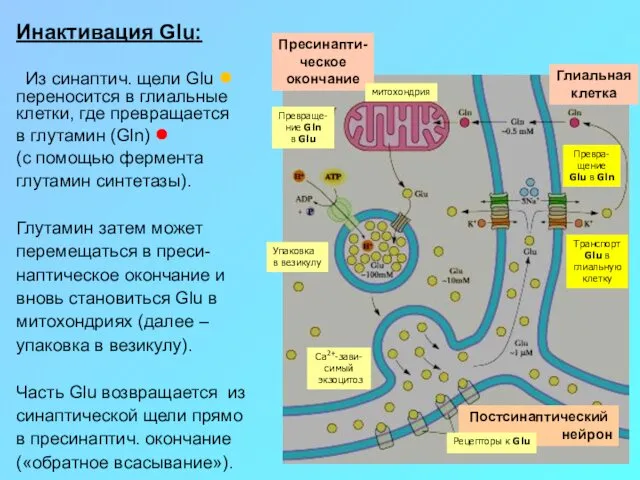
Инактивация Glu:
Из синаптич. щели Glu ● переносится в глиальные клетки, где
Инактивация Glu:
Из синаптич. щели Glu ● переносится в глиальные клетки, где
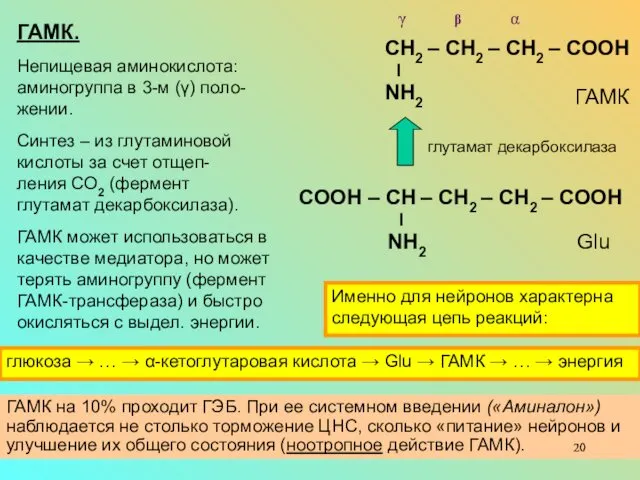
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
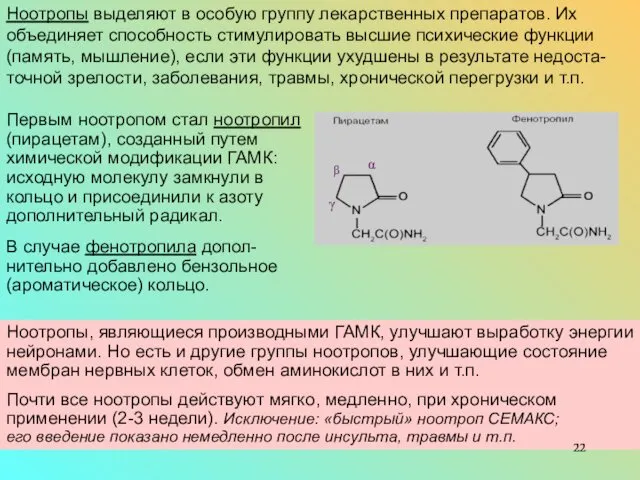
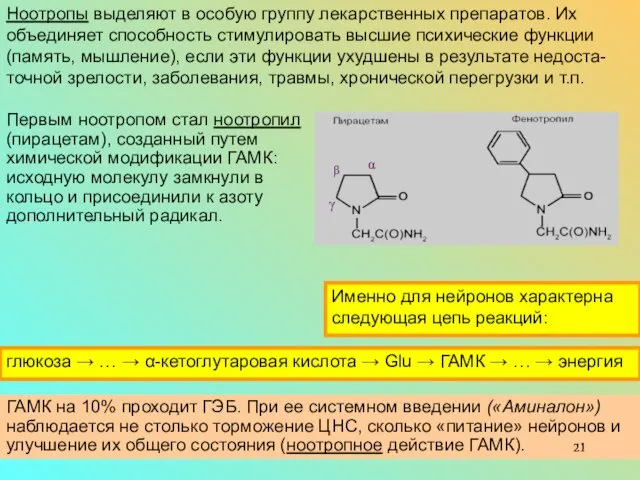
Ноотропы, являющиеся производными ГАМК, улучшают выработку энергии нейронами. Но есть и
Ноотропы, являющиеся производными ГАМК, улучшают выработку энергии нейронами. Но есть и
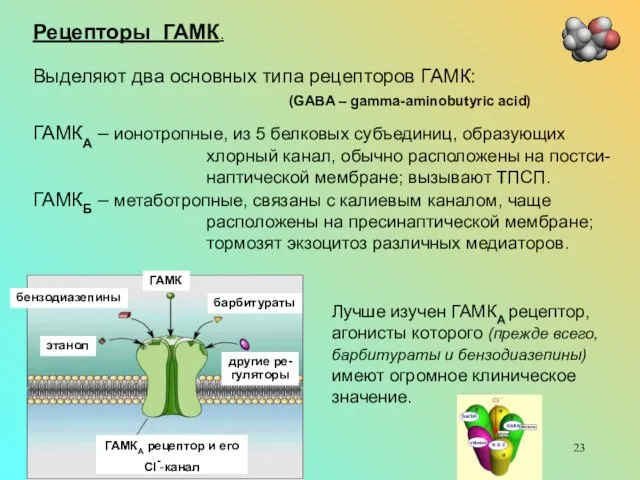
Рецепторы ГАМК.
Выделяют два основных типа рецепторов ГАМК:
(GABA – gamma-aminobutyric acid)
Рецепторы ГАМК.
Выделяют два основных типа рецепторов ГАМК:
(GABA – gamma-aminobutyric acid)

Антагонисты ГАМК: вызывают судороги.
Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа;
Антагонисты ГАМК: вызывают судороги.
Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа;

Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины.
Барбитураты: открыты более 100 лет назад в
Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины.
Барбитураты: открыты более 100 лет назад в

Барбитураты: открыты более 100 лет назад в день Св. Варвары.
продолжительное
Барбитураты: открыты более 100 лет назад в день Св. Варвары.
продолжительное
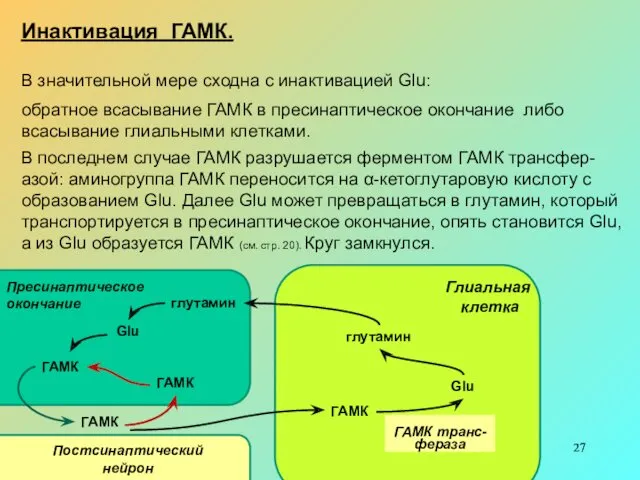
Инактивация ГАМК.
В значительной мере сходна с инактивацией Glu:
обратное всасывание ГАМК
Инактивация ГАМК.
В значительной мере сходна с инактивацией Glu:
обратное всасывание ГАМК
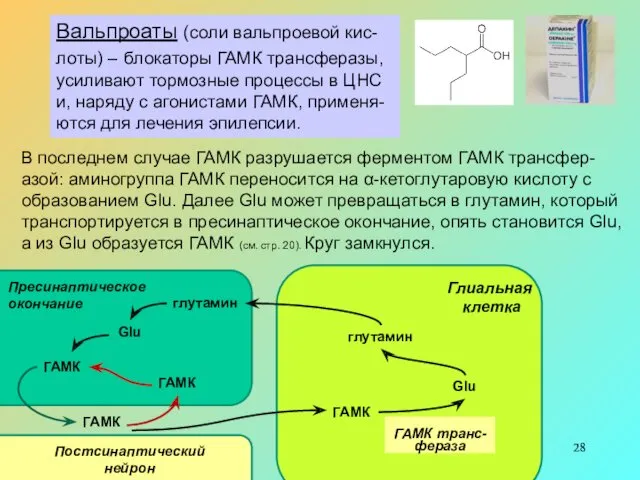
Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы
Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы

Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов
Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов

Тестирование транквилизаторов на эксперимен-тальных животных
Тестирование потен-циальных лекарств на животных – обязательный
Тестирование транквилизаторов на эксперимен-тальных животных
Тестирование потен-циальных лекарств на животных – обязательный
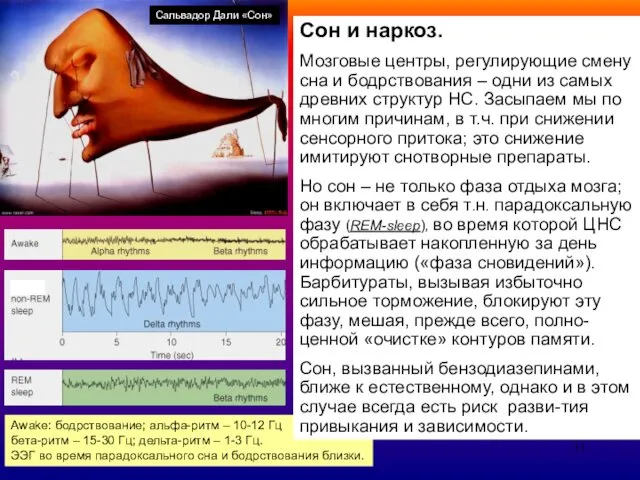
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
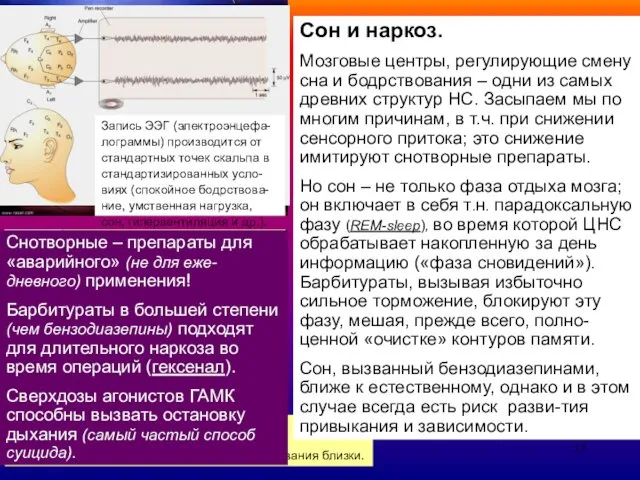
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
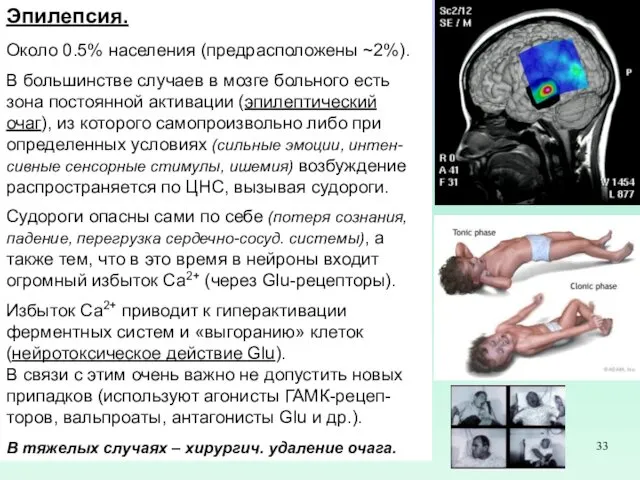
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
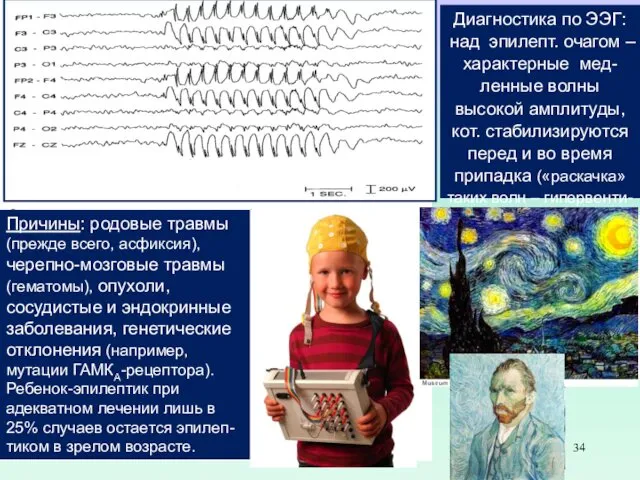
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
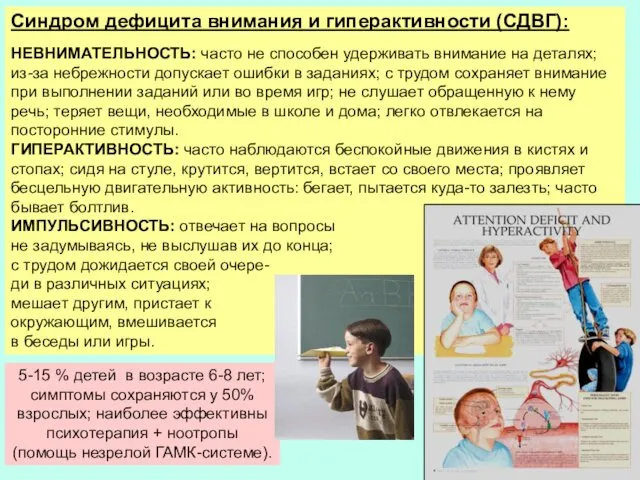
5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%

5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
 Современный урок. 4 лекция
Современный урок. 4 лекция Презентация на пед.совете:Причины и профилактика самовольных уходов детей.
Презентация на пед.совете:Причины и профилактика самовольных уходов детей. Презентация ИКТ на уроках математики
Презентация ИКТ на уроках математики Воды суши. Подземные воды.
Воды суши. Подземные воды. Технология производства молока, приготовление продуктов и блюд из него
Технология производства молока, приготовление продуктов и блюд из него Правила поведения в школе
Правила поведения в школе Основные положения методики построения занятий урочного типа в физическом воспитании
Основные положения методики построения занятий урочного типа в физическом воспитании Подбор персонала в системе управления персоналом
Подбор персонала в системе управления персоналом Реалізація програмного комплексу ip-телефонії на основі PBX Asterisk
Реалізація програмного комплексу ip-телефонії на основі PBX Asterisk Преподобный Леонид Устьнедумский
Преподобный Леонид Устьнедумский Изменения в правовом регулировании деятельности учреждений
Изменения в правовом регулировании деятельности учреждений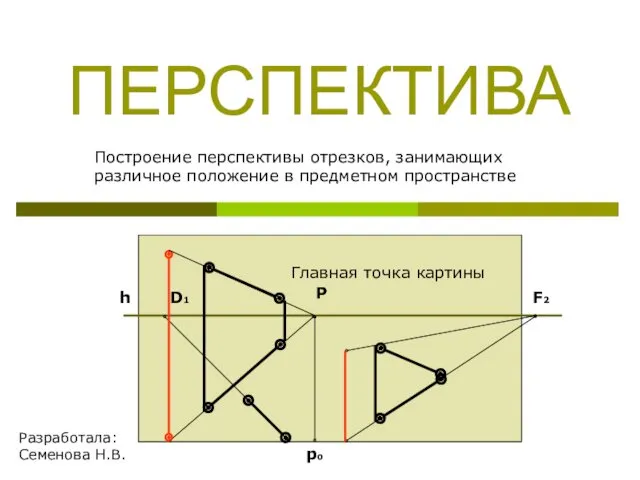 Построение перспективы отрезков, занимающих различное положение в предметном пространстве
Построение перспективы отрезков, занимающих различное положение в предметном пространстве Развитие рынка газомоторного топлива в Российской Федерации
Развитие рынка газомоторного топлива в Российской Федерации Русская равнина
Русская равнина Презентация по теме самообразования Учебная мотивация как показатель качества обучения младших школьников.
Презентация по теме самообразования Учебная мотивация как показатель качества обучения младших школьников. Социальные сети
Социальные сети Казан каласы! Диск
Казан каласы! Диск Установите счётчики. Принцип работы счётчика
Установите счётчики. Принцип работы счётчика Региональная экономика
Региональная экономика Корпус и Блок Питания
Корпус и Блок Питания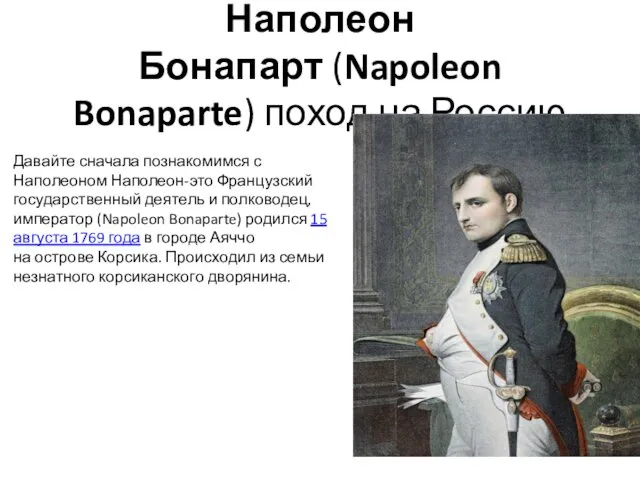 Наполеон Бонапарт (Napoleon Bonaparte). Поход на Россию
Наполеон Бонапарт (Napoleon Bonaparte). Поход на Россию 172262685746922299.1.2
172262685746922299.1.2 Слово. Транскрипция. Перевод
Слово. Транскрипция. Перевод Итоговое устное собеседование. Чтение текста вслух
Итоговое устное собеседование. Чтение текста вслух презентация к уроку в 8 классе Построение модели невидимого объекта. Возникновение в науке теоретических представлений о веществе и химической реакции.
презентация к уроку в 8 классе Построение модели невидимого объекта. Возникновение в науке теоретических представлений о веществе и химической реакции. Температурный ангармонизм. Рассеяние Мандельштама-Бриллюэна. Вынужденное рассеяние Мандельштама-Бриллюэна
Температурный ангармонизм. Рассеяние Мандельштама-Бриллюэна. Вынужденное рассеяние Мандельштама-Бриллюэна 2.4. Матричные синтезы. Часть 2
2.4. Матричные синтезы. Часть 2 микробы
микробы