- Тұқым қуалаушылықтың молекулалық негіздері

Содержание
- 2. ДНҚның негізгі бірлігім-сызықты полимер, төрт мономерлі бірлік арқылы ерекшеленетін дезоксирибонуклеотидтер арқылы нақты сызықты тізбекті анықтайды. Дәл
- 4. ДНҚдағы ақпарат оның дезоксирибонуклеотидтерінің сызықты тізбегінде коделенген. Осы ақпараттың экспрессиясы үшөлшемді жасушаның түзілуіне әкеп соғады. Бірден
- 5. ДНҚдағы дезоксирибонклеотидтердің сызықты қатары ribonucleotide – қосымша тізбектермен рибонуклейн қышқылына айналады. РНҚ тізбегі содан кейін ақуыздың
- 7. Гендер және хромосомалар Негізінен әр жасушаның геномы-генетикалық материалы бірдей болады. Хромосомалар, нуклейн қышқылының молекуласы-жасушадағы ең үлкен
- 8. Гендер туралы біздің түсінік өткен жүздеген жылдықта дамып, белгілі болған. Классикалық ген хромосома бөлігі ретінде анықталған
- 10. Бидл және Татум былай қорытындылады: бұл ген – бір ферментті коделейтін және анықтайтын генетикалық материалдың сегменті.
- 11. Полипептидті тізбектің әр амин қышқылы үш нуклеотид қатарымен кодталған. Олар кодон деп аталады. Эукариоттың көптеген гендері
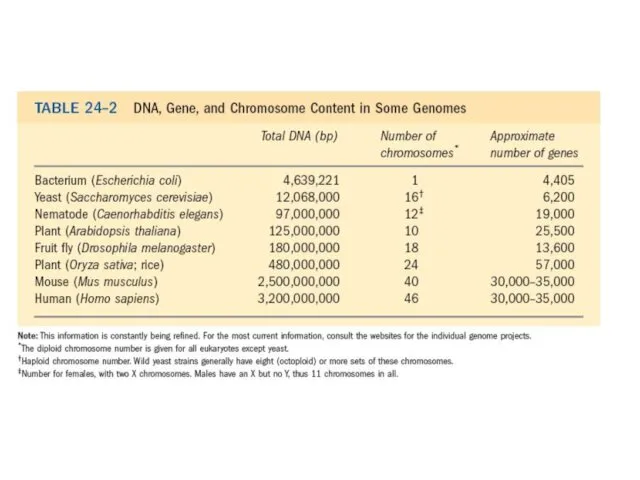
- 12. ДНҚ молекуласы өздері осы ДНҚ дан тұратын клеткаға қарағанда ұзын болып келеді. Ашытқылардың эукариоттық клеткасы еколиға
- 13. Адамның соматикалық жасушасында, мысалы, 46 хромосома бар. эукариоттың әр хромосомасы қос тізбеткі ДНҚдан тұрады. 24 түрлі
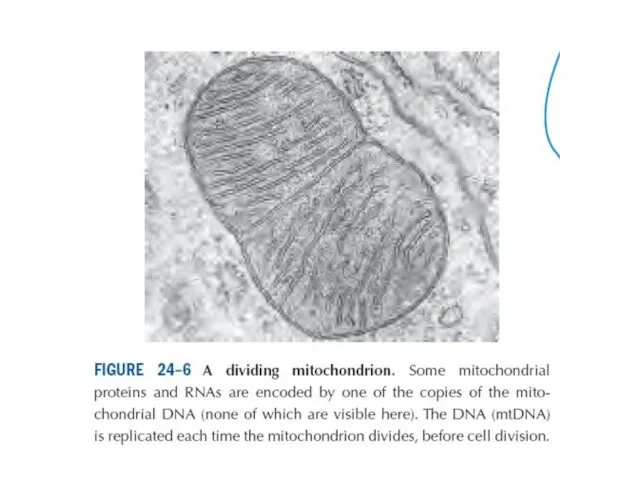
- 14. Эукариот жасушасында сонымен қатар митохондрия және хлоропластар бар. Митохондриялық ДНҚ молекуласы ядролық хромосомаға қарағанда кішкене. Жануар
- 15. Дезоксиаденозин- монофосфат, dAMP Дезокситимидин- монофосфат, dTMP Дезоксигуанозин- монофосфат, dGMP Дезоксицитидин- монофосфат, dCMP
- 16. Полинуклеотидті тізбекке нуклеотидтердің бірігу сызбасы Полинуклеотидті тізбектің сызбасы (сілтеуіш сызықтармен тізбектің өсу бағыты көрсетілген)
- 17. Нуклеотидтер – бұл нуклеинді қышқылдарды құрайтын бірліктер. Қант көміртегінің атомдары үзік сызықты сандармен бөлек нөмірленеді.
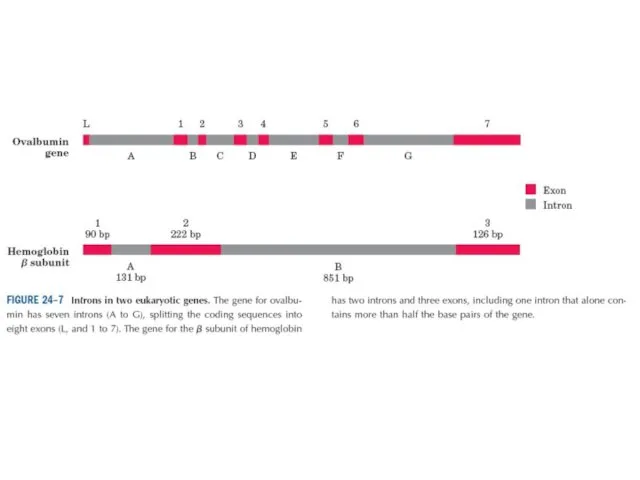
- 23. Эукариоттарда гендер ұйымы күрделірек. Эукариоттық хромосоманың құрылысын зерттеу көптеген күтпеген жағдайларға әкеп соқты. Көбтеген эукариоттық гендерде
- 24. Бірақ бұл элементтер белоктарды және РНҚны коделемейді, олар адам дамуында маңызды орын алған: транспозондардың қозғалысы басқа
- 26. Бұл диаграмма геномды транспазондар, гендер және әртүрлі қатарларға бөледі. Транспозонда рдың төрт басы класы бар. Ұзақ
- 28. ДНҚ бактерияларындағы, бактериофагтердегі және ашықтқылардағы кез келген екі негізіндегі белгілі көршілікте кездесетін жиілік ДНҚ-дағы осы негіздердің
- 29. Әр түрлі ДНҚ-ларындағы кейбір жақын көршілердің кездесуінің жиіліктері
- 30. ДНҚ-да табылған модификацияланған пуриндер мен пиримидиндердің құрылымды формулалары N6-метиладенин 5-метилцитозин 5-гидроксиметилцитозин 5'-гидроксиметил урацил Глюкозил-НМС Гентиобиозил-НМС
- 31. ДНҚ молекуласы шамамен тұрақты диеметрге ие және жиі орналасқан қайталанатын бөлімдерден тұратындығы белгілі, сонымен бірге оның
- 32. ДНҚ-ның екі спиралі түріндегі сызбалық кескін. Кішкентай және үлкен желобоктар көрініп тұр. Жақын негіздердің жұптарының арасындағы
- 33. А және G екі пуриннің және Т және С екі пиримидтердің молярлық қатынастары түрлі ағзалардың ДНҚ-лары
- 34. Сутектік байланыспен ДНҚ-да байланысқан негіздер жұбы. Аденин екі сутектік байланыстың салдарынан тиминмен жұп құрайды, ал гуанин
- 35. Азғантай жиілікпен модификацияланған пуриндер мен пиримидтер ДНҚ-да кездеседі, олар да модификацияланбаған аналогтары сияқты сондай сутектік байланыстарда
- 36. Екі спиральді пішінді ДНҚ-ның В-кеңістіктік моделі. Негіздердің жұптары Кішкентай желобок Үлкен желобок
- 37. Әр түрлі ДНҚ-ның нуклеотидті құрамы.
- 38. ДНҚ-дағы екі спиральдің алтернативті пішіндері негіздер мен қант арасындағы бұрыштар өзгеруі мүмкін болатындығының салдары. Ал дезоксирибозды
- 39. В-, А- және Z-ДНҚ-ның кеңістіктік моделдері. (әрбір модель негіздердің 20 жұбын құрайды). Фосфордың атомдары және олармен
- 40. ДНҚ-дағы В-пішініне тән ерекшелік екі тізбектегі қантфосфатты остов оң спиральді құрайды. Алайда ДНҚ-ның бөліктері белгілі шарттарда
- 41. Әдетте ДНҚ молекуласының өлшемі нуклеоидтер жұбының санында өрнектеледі, сонымен бірге бірлік ретінде нуклеоидтердің мыңдаған жұбын алады.
- 42. Түрлі тектің ДНҚ-ның молекулалық массасы, ұзындығы және құрылымы.
- 43. Ерітіндінің температурасын арттырғандағы екі тізбекті ДНҚ-ның денатурациясы (диссоциация) және салқындату кезіндегі екі комплементарлы тізбектердің ренатурациясы.
- 44. Температураны және рН арттыру арқылы алатын екі тізбекті ДНҚ тәріздес қисық денатурациялар. Тт и рНm -
- 45. Тұздың төмен және жоғары концентрацияларындағы ДНҚ-ның гуанинінің және цитозинінің молекулалық құрамының Tm тәуелділігі. Нүктелер бактериялардың, бактериофагтардың,
- 46. Денатурация – қайтымды процесс, ол толық ыдыраудың нәтижесінде де қайта қалпына келуі мүмкін. Бірігу процесі ренатурация,
- 48. Жасушаларда немесе вирустарда ДНҚ ешқашанда еркін түрде болмайды. форме. Ол төмен молекулалық катионмен байланысты. өзара әсер
- 63. Эурариоттық жасушалардың хромосомалары негізінен хромотиннен тұрады – екі тізбекті ДНҚ кешенінен және бес гистонды ақуыздардан, олар
- 64. Диаметрі 10 нм болатын талшықтар құрылымы жоғарырақ реттіліктің құрылымының пайда болуының салдарынан келесі конденсацияға ұшырауы мүмкін.
- 65. Ұсақ қоректі гистондардың типті сипаттамалары
- 66. Хромотиннің электронды микро суреті. А. CV1 маймылдың бүйректік жасушаларының хромотинінің диаметрі 10 нм болатын талшықтары. Б.
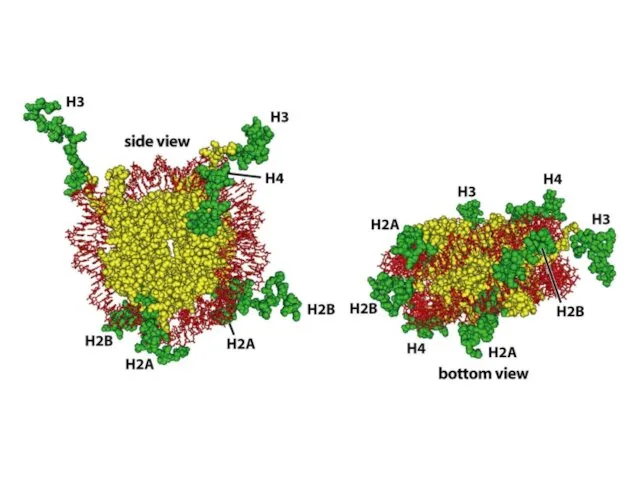
- 67. Төмен және жоғары кеңейтілімнің кристаллографиялық талдауының мәліметтері бойынша салынған нуклеосомды кораның моделі. ДНҚ сегменті (145 незіздердің
- 68. Гистон Н1 нуклеосомды кораны орауды бастайтын және аяқтайтын орындарда ДНҚ-ны «араластырады».
- 69. Конденсацияның түрлі дәрежесіндегі хромотиннің құрылымы. Суреттің төменгі бөлігінде созылған пішіндегі хроматин берілген. Келесі диаметрі 10 нм
- 70. Гистондар ДНҚ-мен байланыса отырып, өз қызметтерін атқарады. Жоғарыда қарастырылған құрылымдық қызметтерден басқа гистондар гендер белсенділігін реттеуде,
- 72. Схеманың жоғарғы бөлігінде-ДНҚ метилденбеген, гистон құйрығы ацетилденген, ол транскрипциялық факторға және РҚ полимеразаға ДНҚ тізбегіне отыруға
- 73. Гистондар көптеген әртүрлі биологиялық процестердің сигналды жолдарын интеграциялайды, соның ішінде ДНҚ метаболизмін. Гистон модификациясы процесінің тепе-теңдікте
- 74. Гистондардың ацетилденуі Гистондардың N- ұштық «құйрықтары" қалдықтарынан тұрады. Құйрықтың аминқышқылдық қатары, әсіресе лизиннің қалдығының орны, қатаң
- 76. Белсенді гендер негізінен гиперацетилденген хроматинде орналасқан, белсенді еместері гипоацетилденген хроматинде орналасқан. Транскрипцияны белсендіретін транскрипциялық факторлар НАТ-ты
- 78. Гистондардың фосфорлануы Гистондардың фосфорлануы сериндік қалдықтар бойынша жүзеге асады. Н3 гистонының 10 сериннің фосфорлануы сүтқоректілер жасушасында
- 79. Гистондардың метилденуі Лдену интенсивтілігі жасушалық цикл аралығында өзгеретіндігі белгілі. Гистондардың метилденуі S -дан G2 – фазаға
- 80. Гистондардың модификациясы гетерохроматиндегі негізгі биомолекулалық әсерлесу болып табылатын 9 лизин қалдығы бойынша метилденетін Н3 гистонмен байланысатын
- 82. Гистондардың убиквитинирленуі Убиквитинирлену дегеніміз үлкен емес ақуыз убиквитиннің ақуызына убиквитин-лигаз арқылы қосылу, деградацияның иницирленуі үшін посттрансляциялы
- 84. Гистондардың сумоилирленуі Сумоилирлену убиквитинирленумен ұқсас, ол үлкен емес убиктивин тектес ақуыздардың қосылысы болып табылады (Small Ubiquitin-like
- 85. Эукариоттармен, геномдармен салыстырғанда прокариот өте ыңшамды құрылған. Кодсыз тізбекті нуклеотидтердің саны минималды, интронды сирек. Ақуыздарды кодтауға
- 86. Хромосомалардағы сияқты эукариот, нуклеоидтің ДНҚ-сы ақуыздармен байланысты болатын көптеген ДНҚ-лармен байланысты. Негізінен гистон тәріздес HU, H-NS
- 87. а – криофиксация көмегімен алыңған бактериалды жасушалардың қималарының электронды-микроскопиялық суреті. 1 және 2 – бір сурет.
- 88. ДE. Coli ДНҚ-ның хросомды ұзындығы ∼4,6 млн. п.о. құрайды.
- 89. Электронды микроскопияның көмегімен жасалған бактериалды жасушаны зерттеу нуклеоидтер рибостардан еркін болатын, диффузиялық боялған аумақтар түрінде болатындығын
- 90. Хроматиннің құрылымының кейбір түрлері тұқым қуалауы мүмкін; яғни құрылым тікелей клеткалардан ұрпаққа берілуі мүмкін. Өйткені бұл
- 92. Құбылыс әсері ДНҚ-ның гетерохроматин өңірлері, әдетте, транскрипцияланбайды және транскрипциялық реттеудің маңызды факторы болып табылады. Егер инверсия
- 93. Көптеген ғалымдар нуклеосомаларды «бейтарап тұрғындар» деп санай отыра, эукариоттардағы гендердің реттелуі нуклеосоманың айналасында жүреді деп есептейді.
- 94. Гистондардың гендердің реттелуі үшін маңызды емес деген көзқарасқа қарсы тұрудың екінші себебі төрт негізгі гистондардың реттіліктеріндегі
- 95. Жасушалардың кейбір субпопуляциясында гендердің репрессияланған күйі тұрақты тұқымқуалауға жақын эухроматикалық аймақтардың гетерохроматизациясы мозаикалық типті әсер (PVE)
- 96. Дрозофила гетерохроматин аймағын эухроматин аймағына тікелей байланыстыратын, хромосомалық бұзылу жерлеріне жақын эухроматикалық гендерді инактивтеуге бейім. Инактивация
- 101. И.Ф. Жимулева зертханасында компактивтіліктің басталу орталықтарының айналасында белгілі бір тығыздауыш белоктарының таралуына негізделген мозаика түрінің позицияларының
- 103. Х-хромосома инактивациясы Гомологтық хромосомалардың және олардың аймақтарының дифференциалдық белсенділігі гендердің дифференциалды белсенділігінің мысалы болып табылады. Лайонизация
- 105. Лайон көрсеткіштеріне сәйкес: 1) гетеропикнотикалық Х хромосомасы бір жануардың әртүрлі жасушаларында атадан немесе анадан шыққан болуы
- 107. Х-хромосомалардың біреуі эмбриональдық дамудың ерте кезеңдерінде инактивтенеді. Х-хромосомалардың екеуі де ооциттерде және ерте бөліну кезінде жұмыс
- 108. Адамдар мен макакалардың жыныстық хроматині 10-12 күндік бластоцистің трофобласт клеткаларында және 16-19-ші күндерде эмбриональды жасушаларда анықталды;
- 109. П. Рассел 1965 жылы. X хромосомасының инактивациялануы Xic (X-inactivation center) деп аталатын және адам мен тінтуірдің
- 110. Х хромосомасын инактивациялаудың молекулалық механизмдерінің екі үлгісі ұсынылған. 1. Лайонизация басталмас бұрын, Xist-транскрипт дамудың нақты реттелетін
- 111. 2. РНК-ның тұрақсыз нұсқасы лайонизация басталғанға дейін екі аллельмен де транскрибирленеді. Х хромосомаларының біреуінің инактивациясының басталуында
- 112. X хромосомасы бойынша инактивация қалай таралады? Бірінші модель. Xist РНҚ хромосомада «цис-жылжыту» арқылы локализацияланған ақуыздың (way-station
- 114. Нуклеосомалық ядроның барлық төрт аминқышқылдарының бүйірлік тізбектері, лизиндердің ацетилденуін, моно-, ди- және триметилденуін қоса әртүрлі ковалентті
- 119. Нуклеосомадағы аминқышқылдардың модификациясы нақты ферменттермен жүзеге асырылады, бұл тек бір немесе бірнеше аймақта әрекет ететін ферменттер.
- 120. Бұл ферменттердің бастапқы «жиналуы» негізінен хромосомалардың ұзындығы бойынша кейбір ДНҚ тізбектерін байланыстыратын гендерді реттейтін ақуыздарға тәуелді
- 123. Негізгі гистондар көбінесе жасушалық циклдың S-фазасы кезінде синтезделеді және репликациялық шанышқыдан кейін ДНҚ-ның жаңа түзілген жіпшелері
- 128. Шағын ақуыздық модульдер басқа модульдермен бірге жұмыс істеп, кодтық оқу кешені құрамында әрекет етеді; бұл хроматиндегі
- 130. Мозаикалық әсердің құбылысы хроматиннің кейбір модифицирленген формаларының хромосомалық ДНҚ молекуласы бойында елеулі қашықтыққа таралуын талап етеді.
- 136. Қазіргі таңда ДНҚ тізбегінің кейбір аймақтарын хромосомаларға қосуға немесе шығаруға мүмкіндік беретін бірнеше «тосқауыл» ДНҚ тізбегі
- 142. Көптеген күрделі ағзалардың, соның ішінде адамдардың әрбір центромері интерфазада сақталатын арнайы центромерлік гетерохроматиннің сегментінде сақталған. Бұл
- 143. Адамдар мен шыбындардың центромерлері жүздеген мың жұп нуклеотидтерге таралады және ДНҚ реттілігіне сай спецификалық центромерасы болмайды.
- 147. De novo центромерасының қалыптасуы бастапқы бастаушы нүктені талап етеді, оның мәні H3 гистонының CENP-A нұсқасының қатысуымен
- 153. Гетерохроматиндер Гетерохроматиндер Эухроматиндер Эухроматиндер
- 155. Скачать презентацию

ДНҚның негізгі бірлігім-сызықты полимер, төрт мономерлі бірлік арқылы ерекшеленетін дезоксирибонуклеотидтер арқылы
ДНҚның негізгі бірлігім-сызықты полимер, төрт мономерлі бірлік арқылы ерекшеленетін дезоксирибонуклеотидтер арқылы
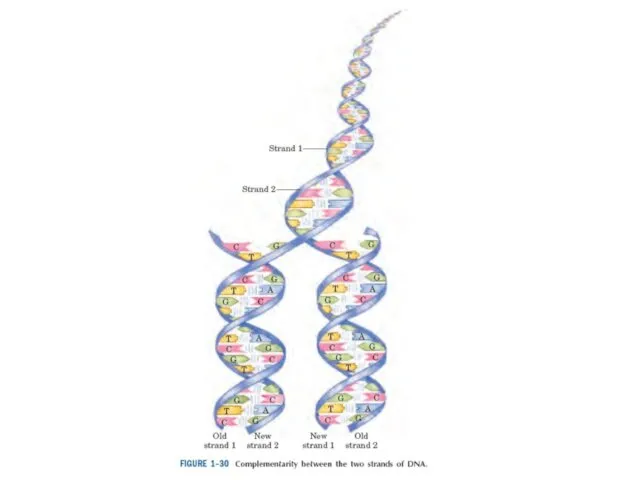

ДНҚдағы ақпарат оның дезоксирибонуклеотидтерінің сызықты тізбегінде коделенген.
Осы ақпараттың экспрессиясы үшөлшемді
ДНҚдағы ақпарат оның дезоксирибонуклеотидтерінің сызықты тізбегінде коделенген.
Осы ақпараттың экспрессиясы үшөлшемді

ДНҚдағы дезоксирибонклеотидтердің сызықты қатары ribonucleotide – қосымша тізбектермен рибонуклейн қышқылына айналады.
ДНҚдағы дезоксирибонклеотидтердің сызықты қатары ribonucleotide – қосымша тізбектермен рибонуклейн қышқылына айналады.
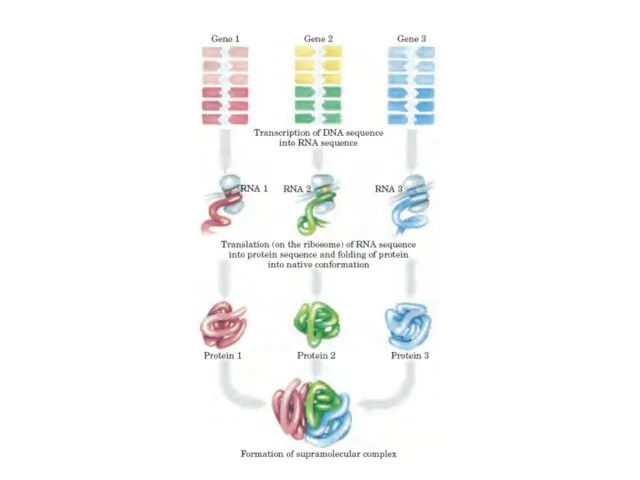

Гендер және хромосомалар
Негізінен әр жасушаның геномы-генетикалық материалы бірдей болады. Хромосомалар, нуклейн
Гендер және хромосомалар
Негізінен әр жасушаның геномы-генетикалық материалы бірдей болады. Хромосомалар, нуклейн

Гендер туралы біздің түсінік өткен жүздеген жылдықта дамып, белгілі болған. Классикалық
Гендер туралы біздің түсінік өткен жүздеген жылдықта дамып, белгілі болған. Классикалық


Бидл және Татум былай қорытындылады: бұл ген – бір ферментті коделейтін
Бидл және Татум былай қорытындылады: бұл ген – бір ферментті коделейтін

Полипептидті тізбектің әр амин қышқылы үш нуклеотид қатарымен кодталған. Олар кодон
Полипептидті тізбектің әр амин қышқылы үш нуклеотид қатарымен кодталған. Олар кодон

ДНҚ молекуласы өздері осы ДНҚ дан тұратын клеткаға қарағанда ұзын болып
ДНҚ молекуласы өздері осы ДНҚ дан тұратын клеткаға қарағанда ұзын болып

Адамның соматикалық жасушасында, мысалы, 46 хромосома бар. эукариоттың әр хромосомасы қос
Адамның соматикалық жасушасында, мысалы, 46 хромосома бар. эукариоттың әр хромосомасы қос

Эукариот жасушасында сонымен қатар митохондрия және хлоропластар бар. Митохондриялық ДНҚ молекуласы
Эукариот жасушасында сонымен қатар митохондрия және хлоропластар бар. Митохондриялық ДНҚ молекуласы
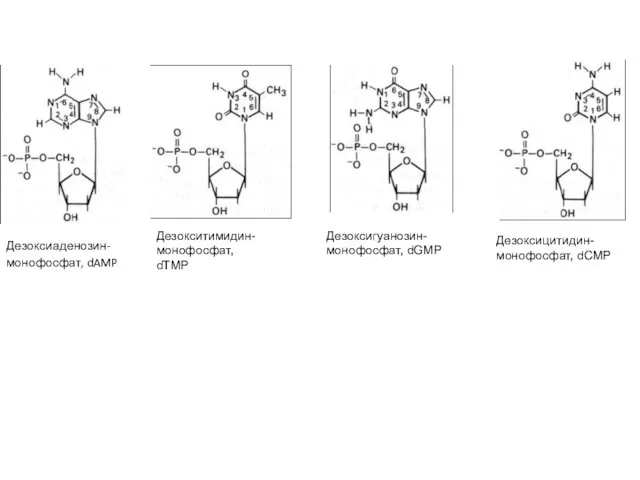
Дезоксиаденозин-
монофосфат, dAMP
Дезокситимидин-
монофосфат, dTMP
Дезоксигуанозин-
монофосфат, dGMP
Дезоксицитидин-
монофосфат, dCMP
Дезоксиаденозин-
монофосфат, dAMP
Дезокситимидин-
монофосфат, dTMP
Дезоксигуанозин-
монофосфат, dGMP
Дезоксицитидин-
монофосфат, dCMP
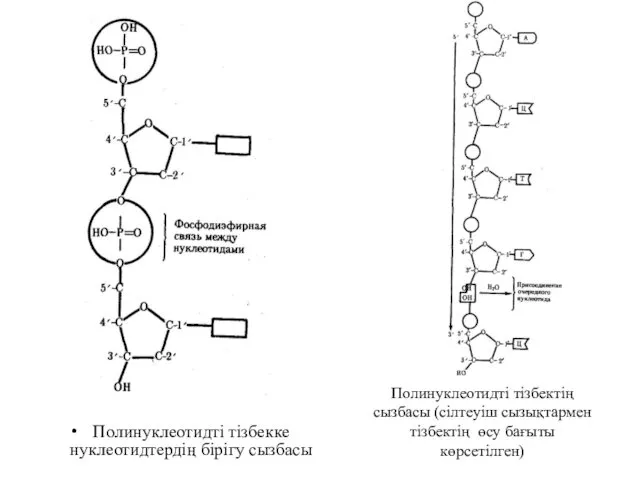
Полинуклеотидті тізбекке нуклеотидтердің бірігу сызбасы
Полинуклеотидті тізбектің сызбасы (сілтеуіш сызықтармен тізбектің өсу
Полинуклеотидті тізбекке нуклеотидтердің бірігу сызбасы
Полинуклеотидті тізбектің сызбасы (сілтеуіш сызықтармен тізбектің өсу
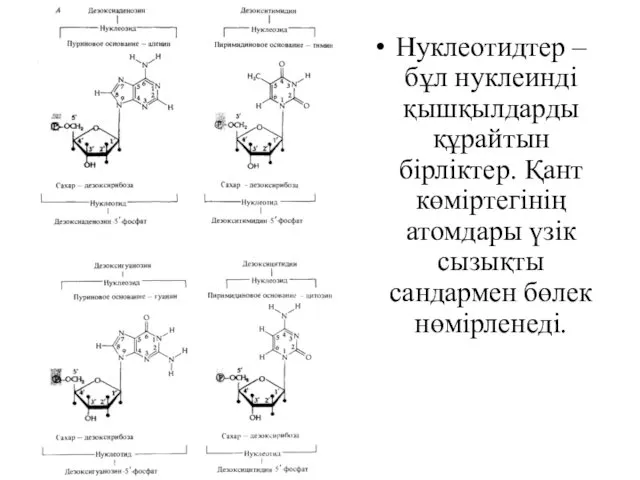
Нуклеотидтер – бұл нуклеинді қышқылдарды құрайтын бірліктер. Қант көміртегінің атомдары үзік
Нуклеотидтер – бұл нуклеинді қышқылдарды құрайтын бірліктер. Қант көміртегінің атомдары үзік



Эукариоттарда гендер ұйымы күрделірек. Эукариоттық хромосоманың құрылысын зерттеу көптеген күтпеген жағдайларға
Эукариоттарда гендер ұйымы күрделірек. Эукариоттық хромосоманың құрылысын зерттеу көптеген күтпеген жағдайларға

Бірақ бұл элементтер белоктарды және РНҚны коделемейді, олар адам дамуында маңызды
Бірақ бұл элементтер белоктарды және РНҚны коделемейді, олар адам дамуында маңызды
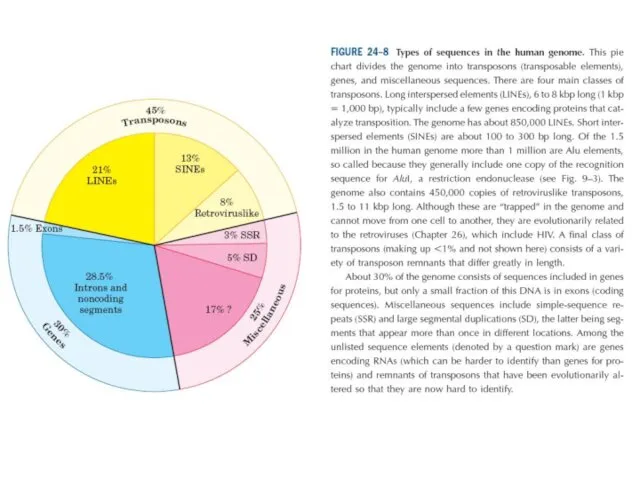

Бұл диаграмма геномды транспазондар, гендер және әртүрлі қатарларға бөледі. Транспозонда рдың
Бұл диаграмма геномды транспазондар, гендер және әртүрлі қатарларға бөледі. Транспозонда рдың

ДНҚ бактерияларындағы, бактериофагтердегі және ашықтқылардағы кез келген екі негізіндегі белгілі көршілікте
ДНҚ бактерияларындағы, бактериофагтердегі және ашықтқылардағы кез келген екі негізіндегі белгілі көршілікте
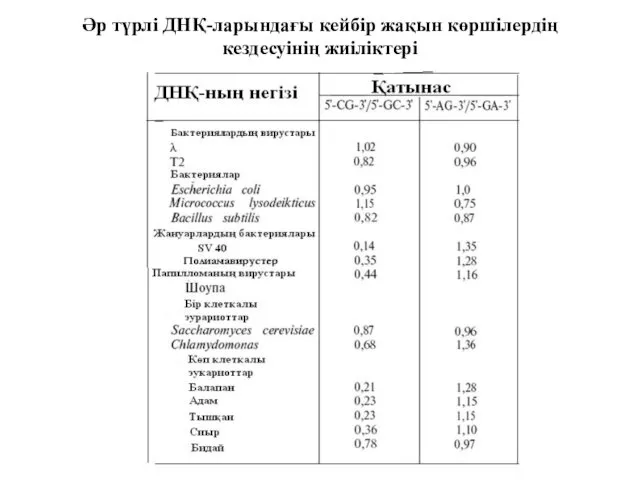
Әр түрлі ДНҚ-ларындағы кейбір жақын көршілердің кездесуінің жиіліктері
Әр түрлі ДНҚ-ларындағы кейбір жақын көршілердің кездесуінің жиіліктері
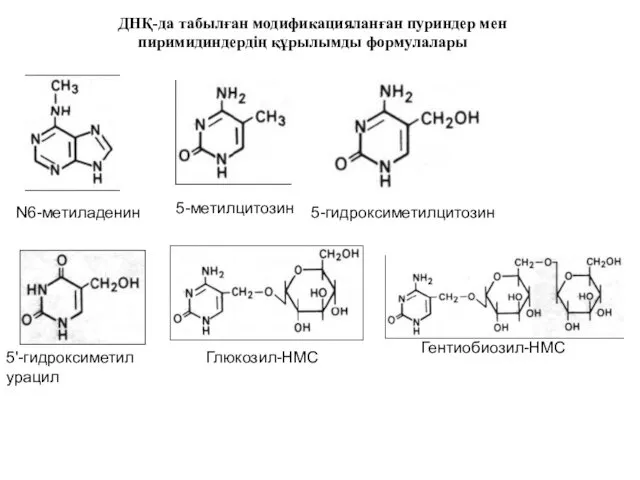
ДНҚ-да табылған модификацияланған пуриндер мен пиримидиндердің құрылымды формулалары
N6-метиладенин
5-метилцитозин
5-гидроксиметилцитозин
5'-гидроксиметил
урацил
Глюкозил-НМС
Гентиобиозил-НМС
ДНҚ-да табылған модификацияланған пуриндер мен пиримидиндердің құрылымды формулалары
N6-метиладенин
5-метилцитозин
5-гидроксиметилцитозин
5'-гидроксиметил
урацил
Глюкозил-НМС
Гентиобиозил-НМС

ДНҚ молекуласы шамамен тұрақты диеметрге ие және жиі орналасқан қайталанатын бөлімдерден
ДНҚ молекуласы шамамен тұрақты диеметрге ие және жиі орналасқан қайталанатын бөлімдерден
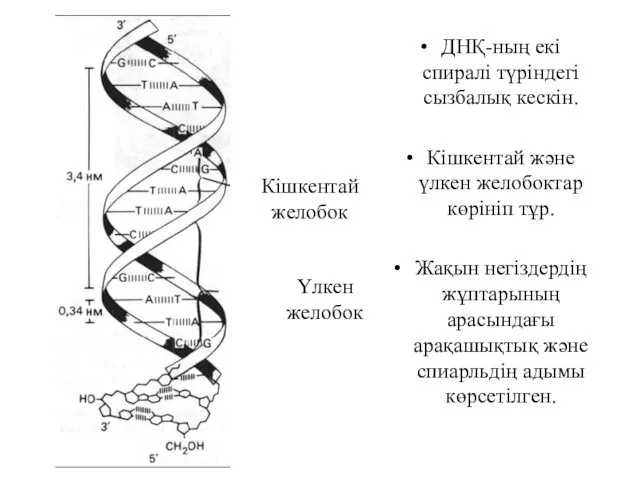
ДНҚ-ның екі спиралі түріндегі сызбалық кескін.
Кішкентай және үлкен желобоктар көрініп
ДНҚ-ның екі спиралі түріндегі сызбалық кескін.
Кішкентай және үлкен желобоктар көрініп

А және G екі пуриннің және Т және С екі пиримидтердің
А және G екі пуриннің және Т және С екі пиримидтердің
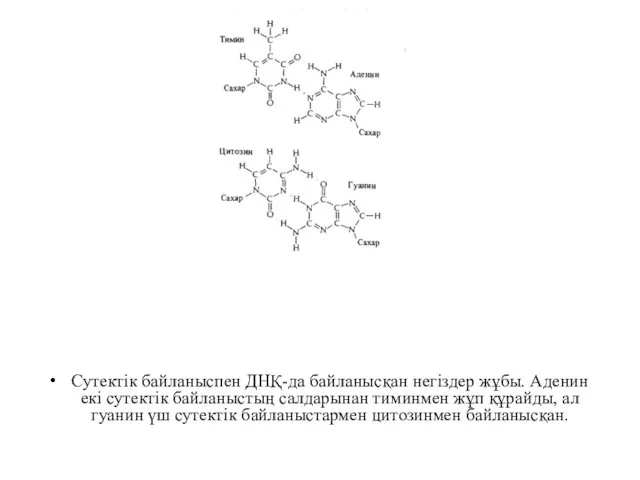
Сутектік байланыспен ДНҚ-да байланысқан негіздер жұбы. Аденин екі сутектік байланыстың салдарынан
Сутектік байланыспен ДНҚ-да байланысқан негіздер жұбы. Аденин екі сутектік байланыстың салдарынан

Азғантай жиілікпен модификацияланған пуриндер мен пиримидтер ДНҚ-да кездеседі, олар да модификацияланбаған
Азғантай жиілікпен модификацияланған пуриндер мен пиримидтер ДНҚ-да кездеседі, олар да модификацияланбаған
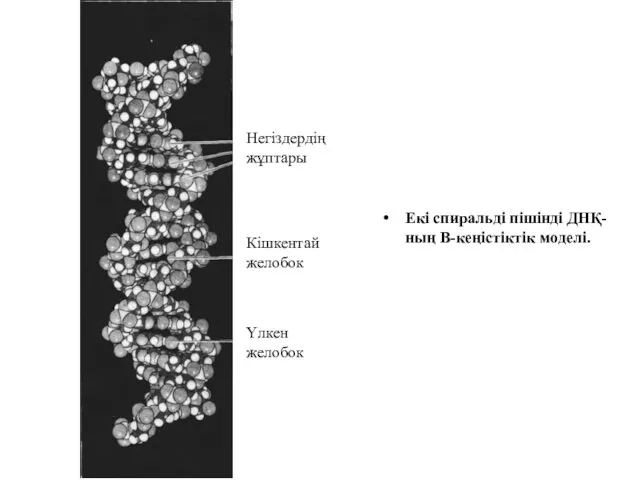
Екі спиральді пішінді ДНҚ-ның В-кеңістіктік моделі.
Негіздердің жұптары
Кішкентай
желобок
Үлкен
желобок
Екі спиральді пішінді ДНҚ-ның В-кеңістіктік моделі.
Негіздердің жұптары
Кішкентай
желобок
Үлкен
желобок
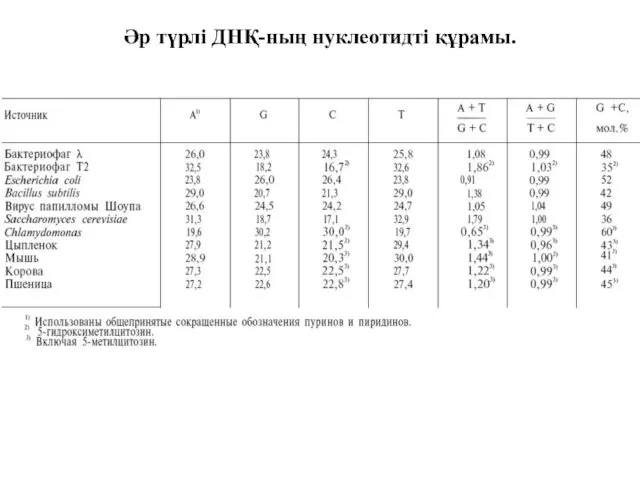
Әр түрлі ДНҚ-ның нуклеотидті құрамы.
Әр түрлі ДНҚ-ның нуклеотидті құрамы.

ДНҚ-дағы екі спиральдің алтернативті пішіндері негіздер мен қант арасындағы бұрыштар өзгеруі
ДНҚ-дағы екі спиральдің алтернативті пішіндері негіздер мен қант арасындағы бұрыштар өзгеруі
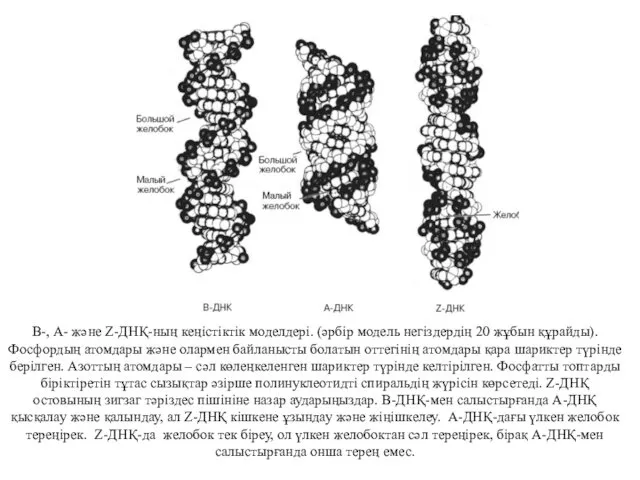
В-, А- және Z-ДНҚ-ның кеңістіктік моделдері. (әрбір модель негіздердің 20 жұбын
В-, А- және Z-ДНҚ-ның кеңістіктік моделдері. (әрбір модель негіздердің 20 жұбын

ДНҚ-дағы В-пішініне тән ерекшелік екі тізбектегі қантфосфатты остов оң спиральді құрайды.

Әдетте ДНҚ молекуласының өлшемі нуклеоидтер жұбының санында өрнектеледі, сонымен бірге бірлік
Әдетте ДНҚ молекуласының өлшемі нуклеоидтер жұбының санында өрнектеледі, сонымен бірге бірлік

Түрлі тектің ДНҚ-ның молекулалық массасы, ұзындығы және құрылымы.
Түрлі тектің ДНҚ-ның молекулалық массасы, ұзындығы және құрылымы.
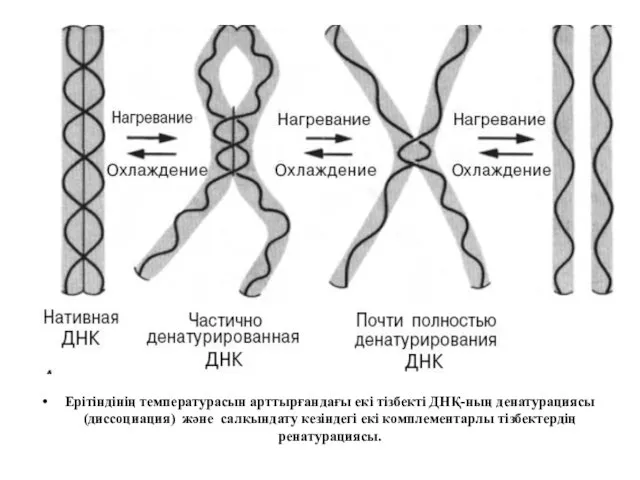
Ерітіндінің температурасын арттырғандағы екі тізбекті ДНҚ-ның денатурациясы (диссоциация) және салқындату кезіндегі
Ерітіндінің температурасын арттырғандағы екі тізбекті ДНҚ-ның денатурациясы (диссоциация) және салқындату кезіндегі
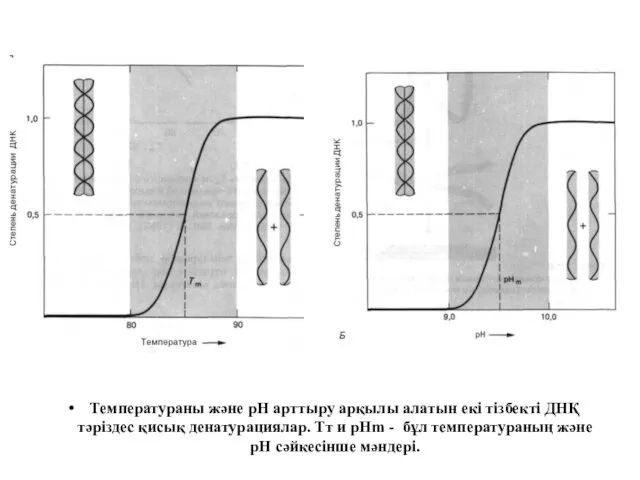
Температураны және рН арттыру арқылы алатын екі тізбекті ДНҚ тәріздес қисық
Температураны және рН арттыру арқылы алатын екі тізбекті ДНҚ тәріздес қисық
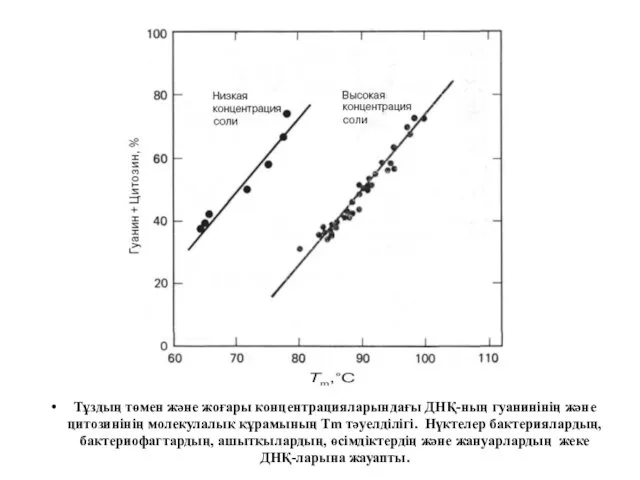
Тұздың төмен және жоғары концентрацияларындағы ДНҚ-ның гуанинінің және цитозинінің молекулалық құрамының
Тұздың төмен және жоғары концентрацияларындағы ДНҚ-ның гуанинінің және цитозинінің молекулалық құрамының

Денатурация – қайтымды процесс, ол толық ыдыраудың нәтижесінде де қайта қалпына
Денатурация – қайтымды процесс, ол толық ыдыраудың нәтижесінде де қайта қалпына
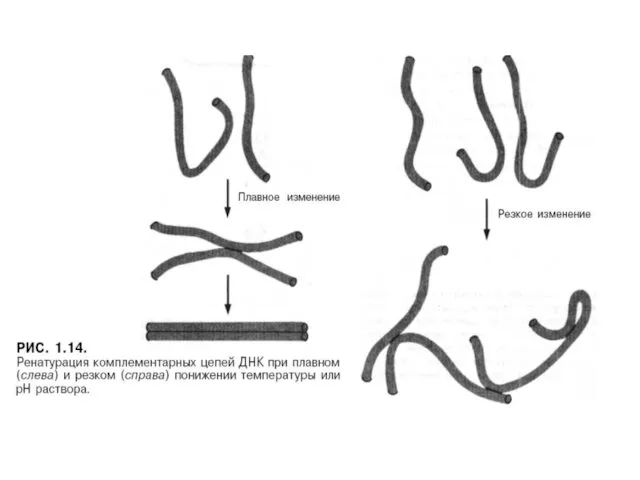

Жасушаларда немесе вирустарда ДНҚ ешқашанда еркін түрде болмайды. форме.
Ол төмен молекулалық
Жасушаларда немесе вирустарда ДНҚ ешқашанда еркін түрде болмайды. форме.
Ол төмен молекулалық
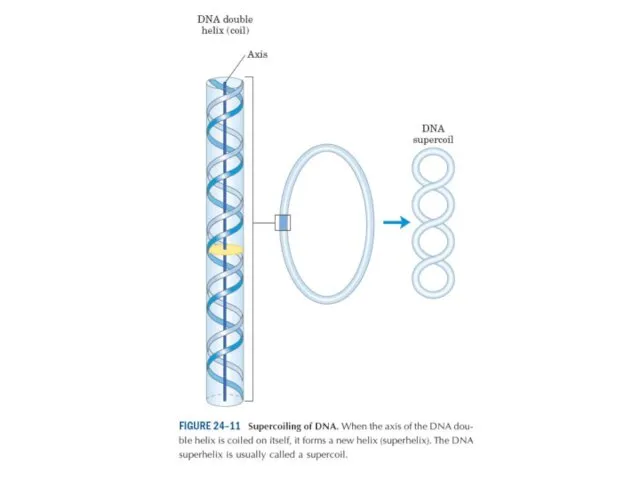

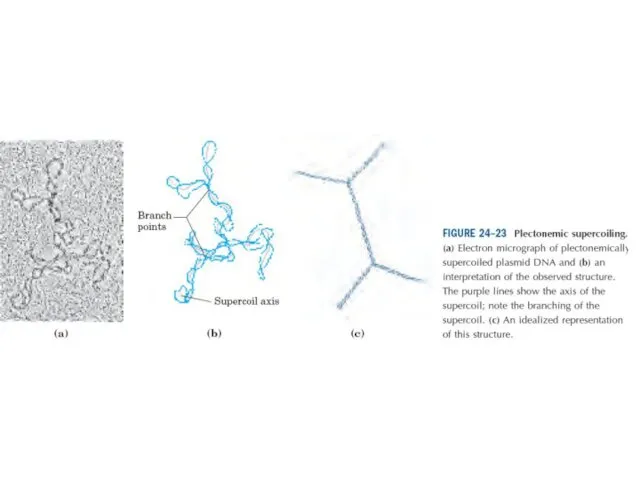
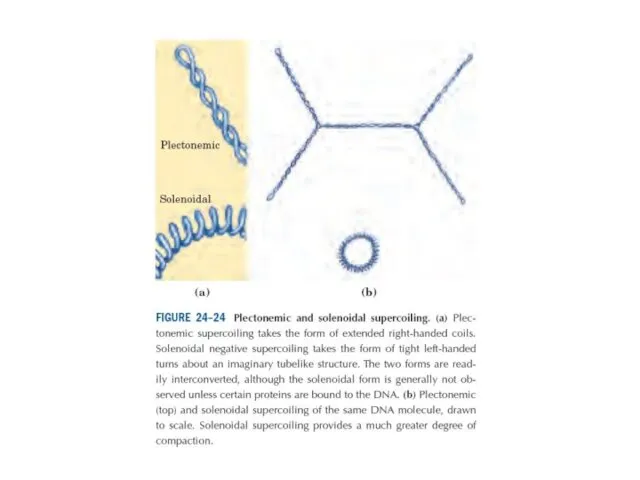

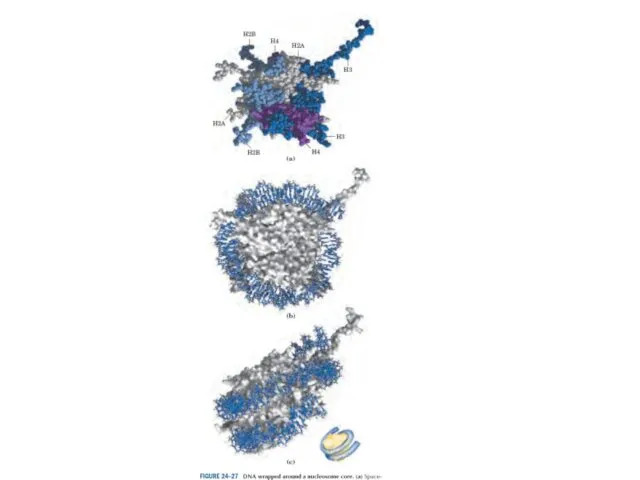
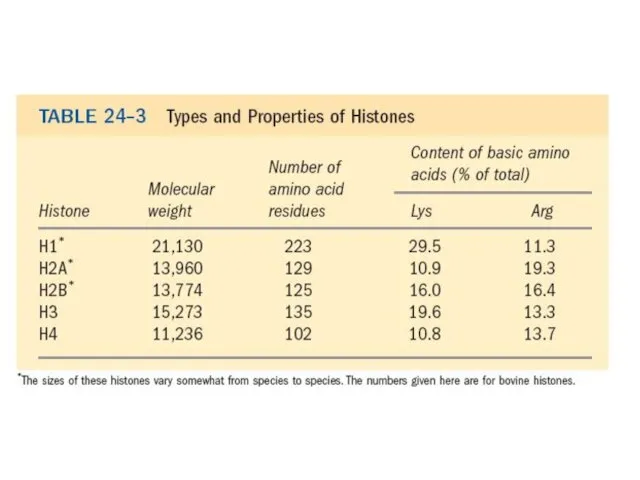
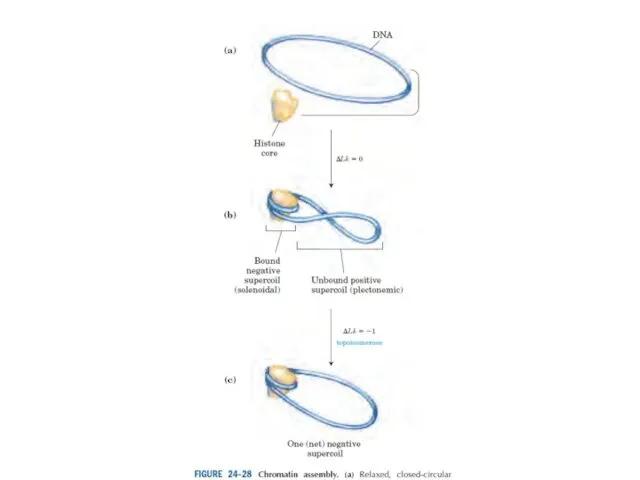
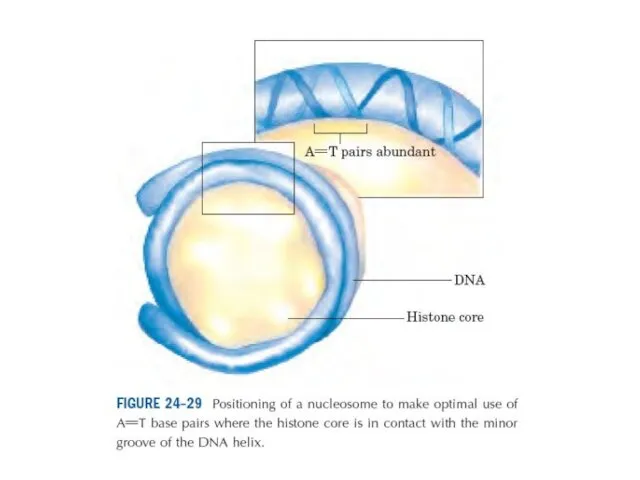
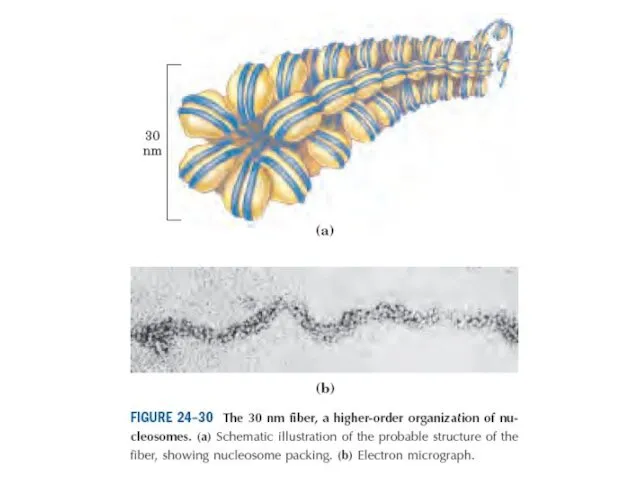

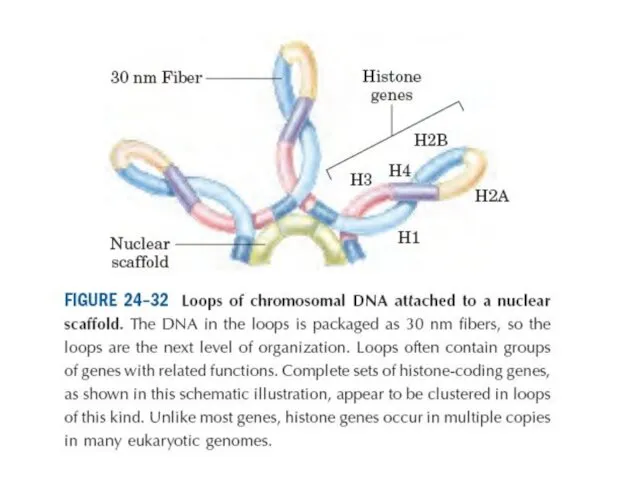
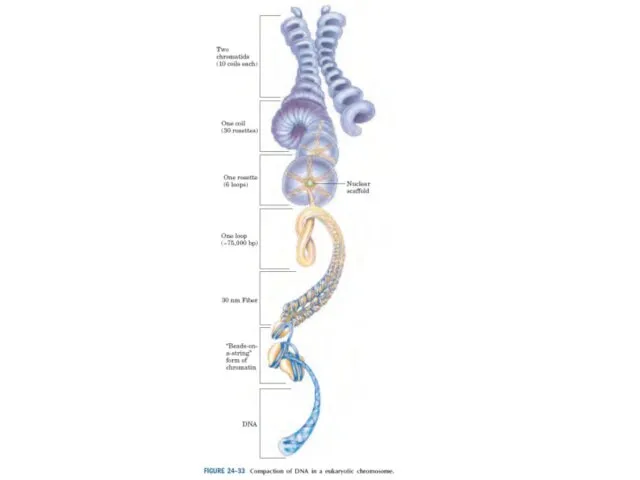

Эурариоттық жасушалардың хромосомалары негізінен хромотиннен тұрады – екі тізбекті ДНҚ кешенінен
Эурариоттық жасушалардың хромосомалары негізінен хромотиннен тұрады – екі тізбекті ДНҚ кешенінен

Диаметрі 10 нм болатын талшықтар құрылымы жоғарырақ реттіліктің құрылымының пайда болуының
Диаметрі 10 нм болатын талшықтар құрылымы жоғарырақ реттіліктің құрылымының пайда болуының
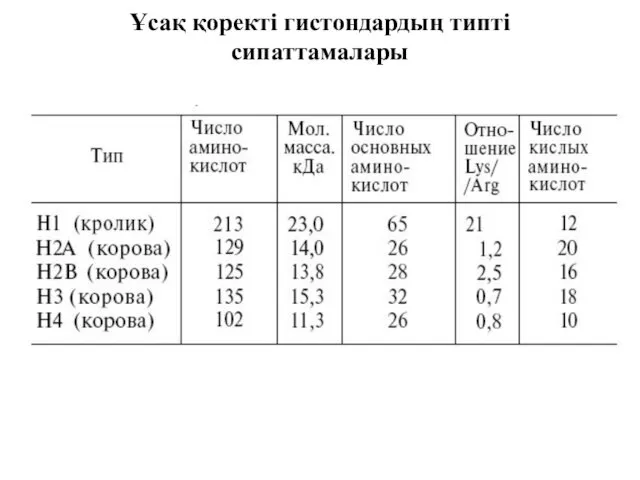
Ұсақ қоректі гистондардың типті сипаттамалары
Ұсақ қоректі гистондардың типті сипаттамалары
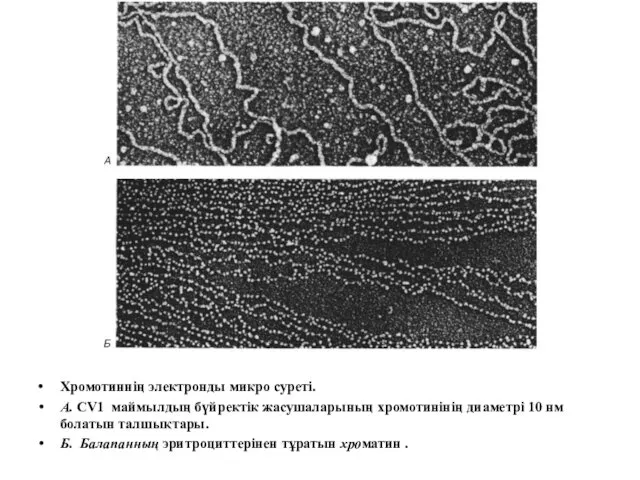
Хромотиннің электронды микро суреті.
А. CV1 маймылдың бүйректік жасушаларының хромотинінің диаметрі
Хромотиннің электронды микро суреті.
А. CV1 маймылдың бүйректік жасушаларының хромотинінің диаметрі
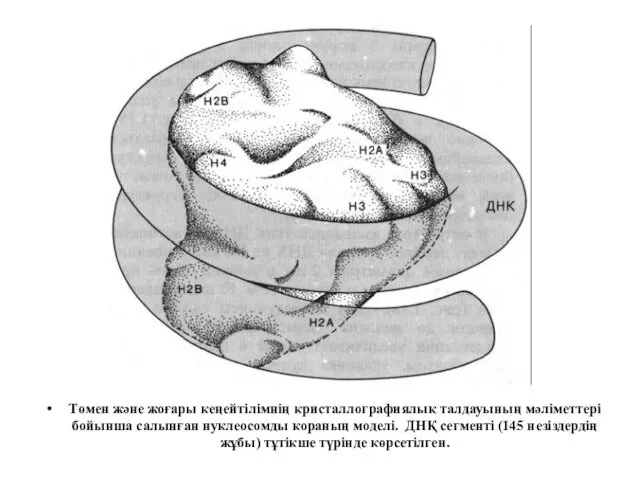
Төмен және жоғары кеңейтілімнің кристаллографиялық талдауының мәліметтері бойынша салынған нуклеосомды кораның
Төмен және жоғары кеңейтілімнің кристаллографиялық талдауының мәліметтері бойынша салынған нуклеосомды кораның
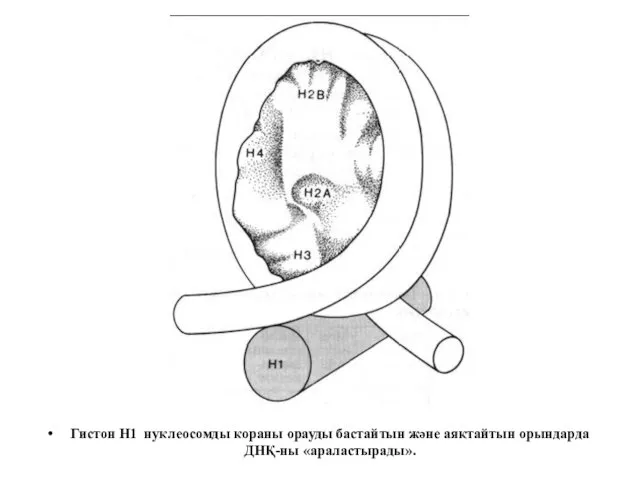
Гистон Н1 нуклеосомды кораны орауды бастайтын және аяқтайтын орындарда ДНҚ-ны «араластырады».
Гистон Н1 нуклеосомды кораны орауды бастайтын және аяқтайтын орындарда ДНҚ-ны «араластырады».
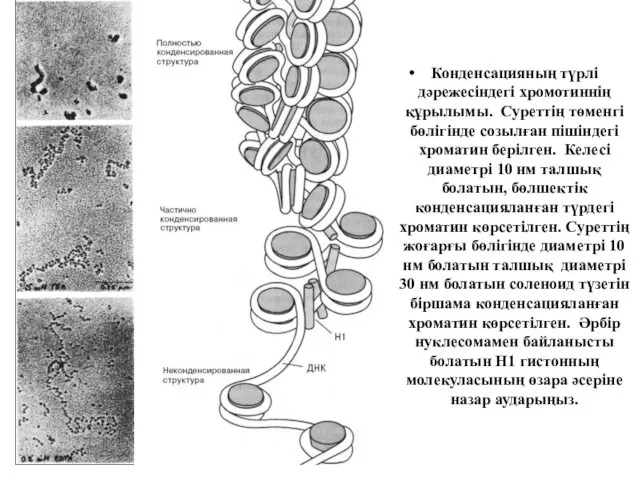
Конденсацияның түрлі дәрежесіндегі хромотиннің құрылымы. Суреттің төменгі бөлігінде созылған пішіндегі хроматин
Конденсацияның түрлі дәрежесіндегі хромотиннің құрылымы. Суреттің төменгі бөлігінде созылған пішіндегі хроматин

Гистондар ДНҚ-мен байланыса отырып, өз қызметтерін атқарады. Жоғарыда қарастырылған құрылымдық қызметтерден
Гистондар ДНҚ-мен байланыса отырып, өз қызметтерін атқарады. Жоғарыда қарастырылған құрылымдық қызметтерден
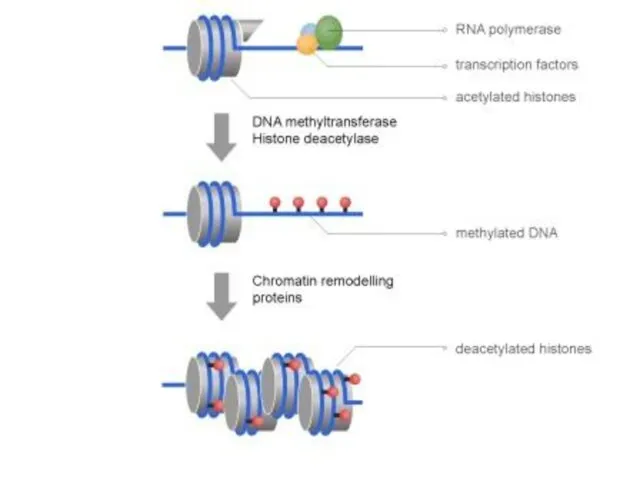

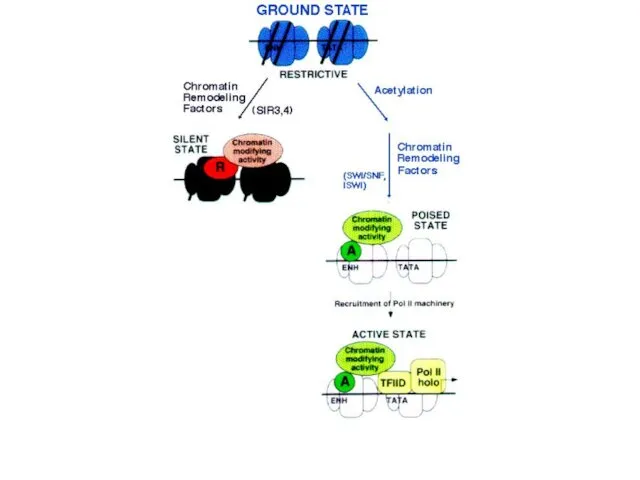
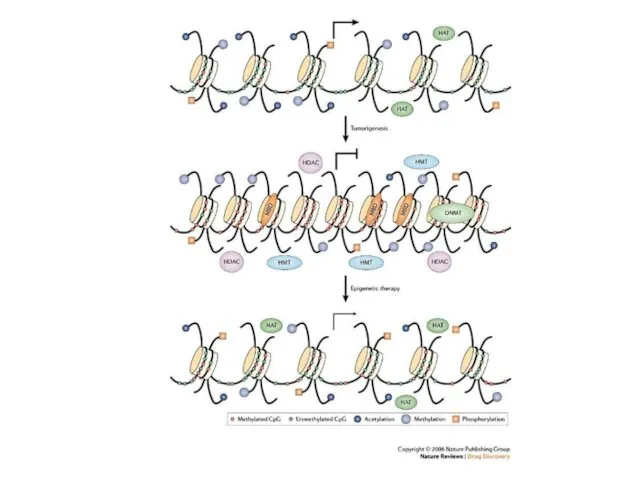
Схеманың жоғарғы бөлігінде-ДНҚ метилденбеген, гистон құйрығы ацетилденген, ол транскрипциялық факторға және
Схеманың жоғарғы бөлігінде-ДНҚ метилденбеген, гистон құйрығы ацетилденген, ол транскрипциялық факторға және
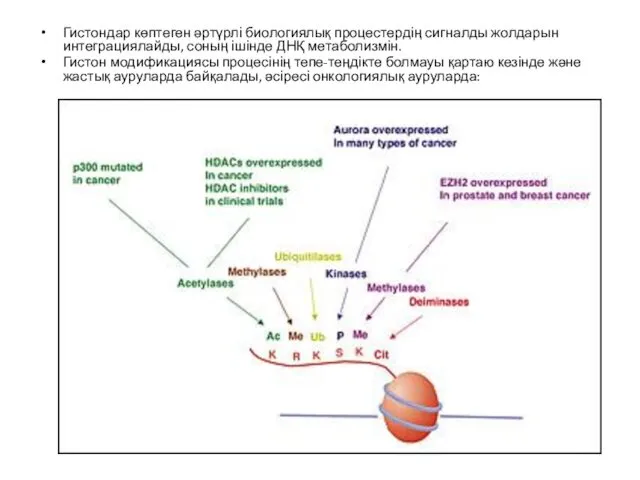
Гистондар көптеген әртүрлі биологиялық процестердің сигналды жолдарын интеграциялайды, соның ішінде ДНҚ
Гистондар көптеген әртүрлі биологиялық процестердің сигналды жолдарын интеграциялайды, соның ішінде ДНҚ

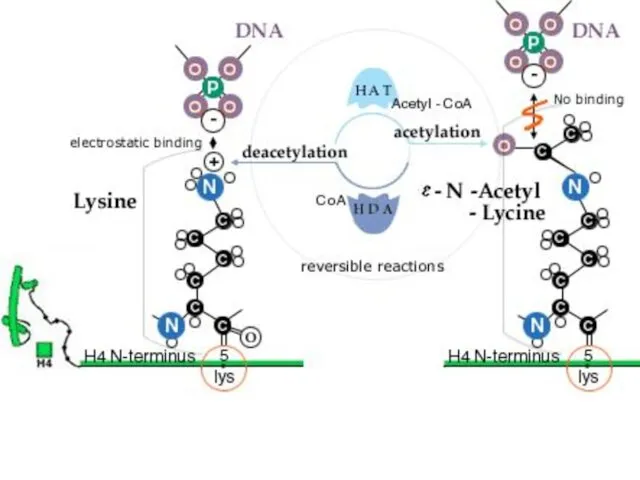
Гистондардың ацетилденуі
Гистондардың N- ұштық «құйрықтары" қалдықтарынан тұрады. Құйрықтың аминқышқылдық қатары, әсіресе
Гистондардың ацетилденуі
Гистондардың N- ұштық «құйрықтары" қалдықтарынан тұрады. Құйрықтың аминқышқылдық қатары, әсіресе

Белсенді гендер негізінен гиперацетилденген хроматинде орналасқан, белсенді еместері гипоацетилденген хроматинде орналасқан.
Белсенді гендер негізінен гиперацетилденген хроматинде орналасқан, белсенді еместері гипоацетилденген хроматинде орналасқан.

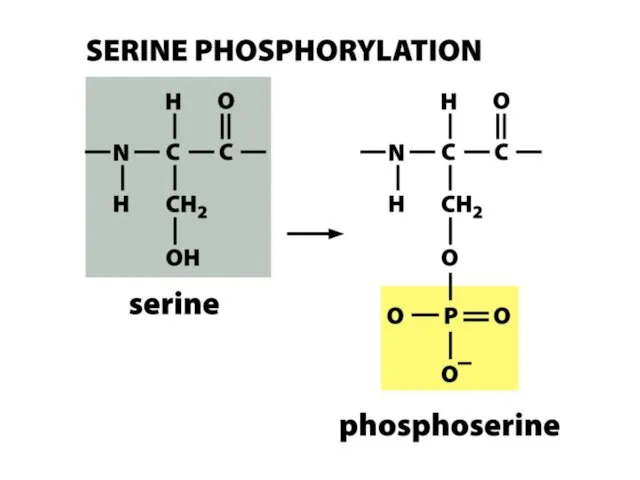
Гистондардың фосфорлануы
Гистондардың фосфорлануы сериндік қалдықтар бойынша жүзеге асады.
Н3 гистонының 10 сериннің
Гистондардың фосфорлануы
Гистондардың фосфорлануы сериндік қалдықтар бойынша жүзеге асады.
Н3 гистонының 10 сериннің

Гистондардың метилденуі
Лдену интенсивтілігі жасушалық цикл аралығында өзгеретіндігі белгілі.
Гистондардың метилденуі S
Гистондардың метилденуі
Лдену интенсивтілігі жасушалық цикл аралығында өзгеретіндігі белгілі.
Гистондардың метилденуі S

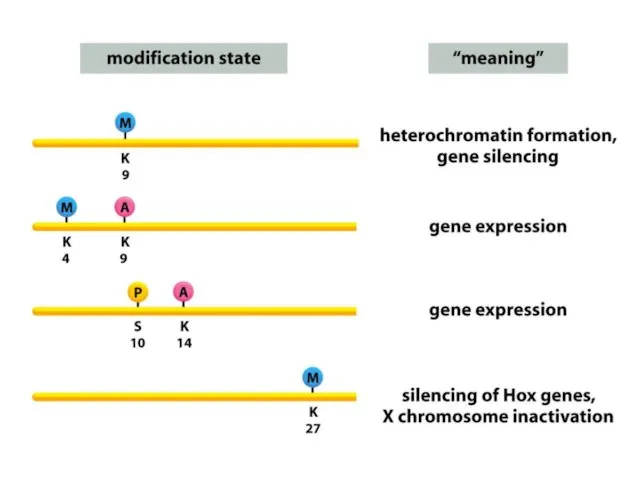
Гистондардың модификациясы гетерохроматиндегі негізгі биомолекулалық әсерлесу болып табылатын 9 лизин қалдығы
Гистондардың модификациясы гетерохроматиндегі негізгі биомолекулалық әсерлесу болып табылатын 9 лизин қалдығы

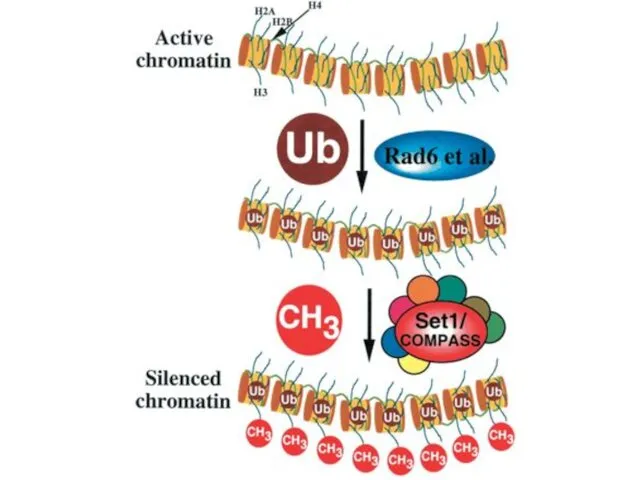
Гистондардың убиквитинирленуі
Убиквитинирлену дегеніміз үлкен емес ақуыз убиквитиннің ақуызына убиквитин-лигаз арқылы қосылу,
Гистондардың убиквитинирленуі
Убиквитинирлену дегеніміз үлкен емес ақуыз убиквитиннің ақуызына убиквитин-лигаз арқылы қосылу,

Гистондардың сумоилирленуі
Сумоилирлену убиквитинирленумен ұқсас, ол үлкен емес убиктивин тектес ақуыздардың
Гистондардың сумоилирленуі
Сумоилирлену убиквитинирленумен ұқсас, ол үлкен емес убиктивин тектес ақуыздардың

Эукариоттармен, геномдармен салыстырғанда прокариот өте ыңшамды құрылған.
Кодсыз тізбекті нуклеотидтердің саны минималды,
Эукариоттармен, геномдармен салыстырғанда прокариот өте ыңшамды құрылған.
Кодсыз тізбекті нуклеотидтердің саны минималды,

Хромосомалардағы сияқты эукариот, нуклеоидтің ДНҚ-сы ақуыздармен байланысты болатын көптеген ДНҚ-лармен байланысты.
Хромосомалардағы сияқты эукариот, нуклеоидтің ДНҚ-сы ақуыздармен байланысты болатын көптеген ДНҚ-лармен байланысты.

а – криофиксация көмегімен алыңған бактериалды жасушалардың қималарының электронды-микроскопиялық суреті. 1
а – криофиксация көмегімен алыңған бактериалды жасушалардың қималарының электронды-микроскопиялық суреті. 1

ДE. Coli ДНҚ-ның хросомды ұзындығы ∼4,6 млн. п.о. құрайды.
ДE. Coli ДНҚ-ның хросомды ұзындығы ∼4,6 млн. п.о. құрайды.

Электронды микроскопияның көмегімен жасалған бактериалды жасушаны зерттеу нуклеоидтер рибостардан еркін болатын,
Электронды микроскопияның көмегімен жасалған бактериалды жасушаны зерттеу нуклеоидтер рибостардан еркін болатын,

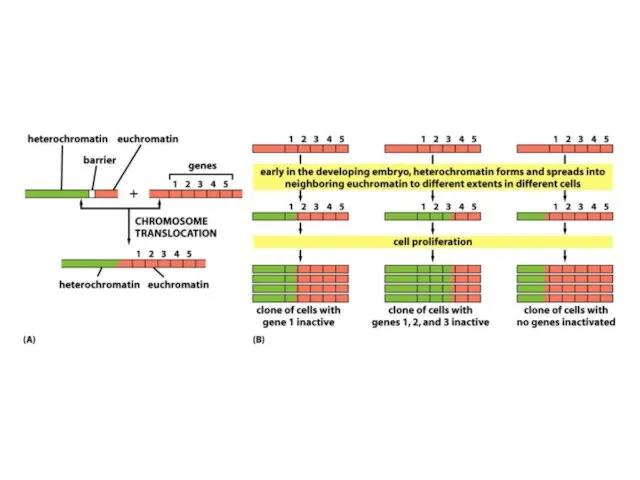
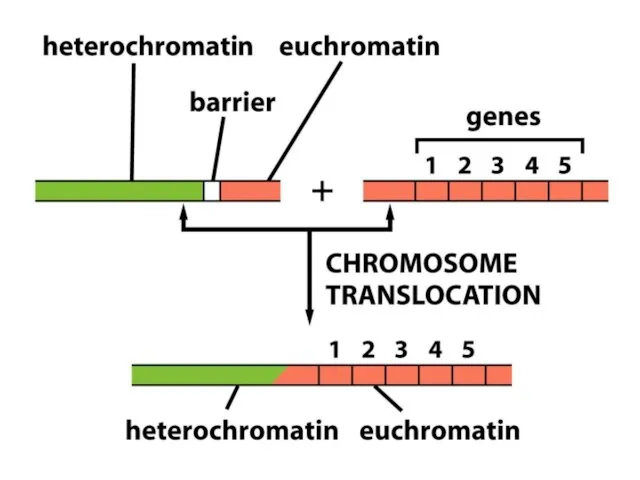
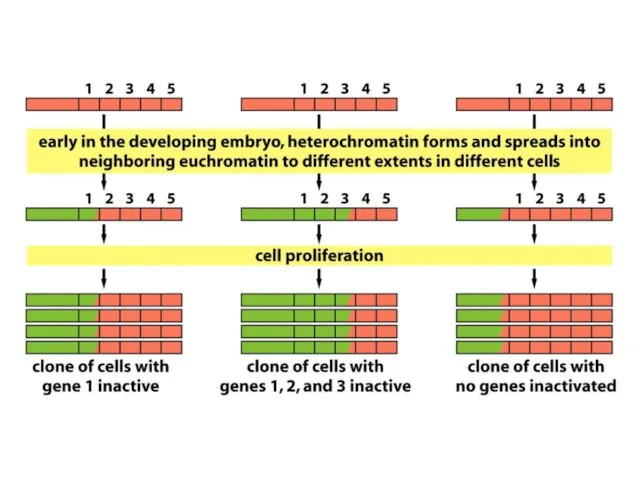
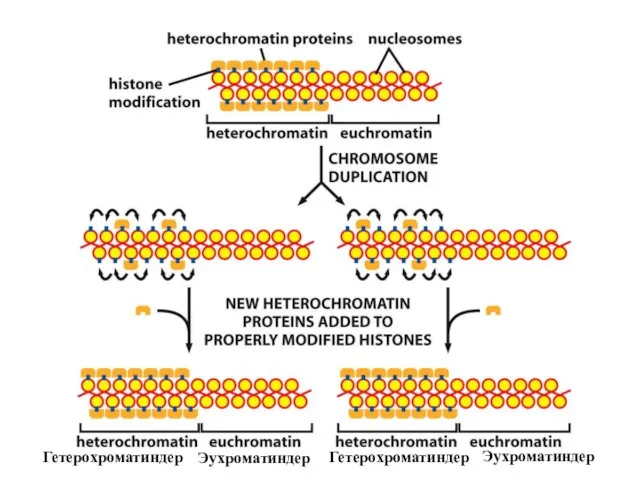
Хроматиннің құрылымының кейбір түрлері тұқым қуалауы мүмкін; яғни құрылым тікелей клеткалардан
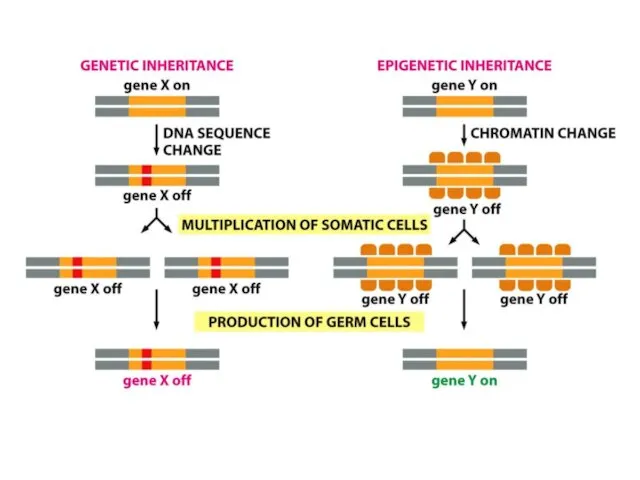

Құбылыс әсері
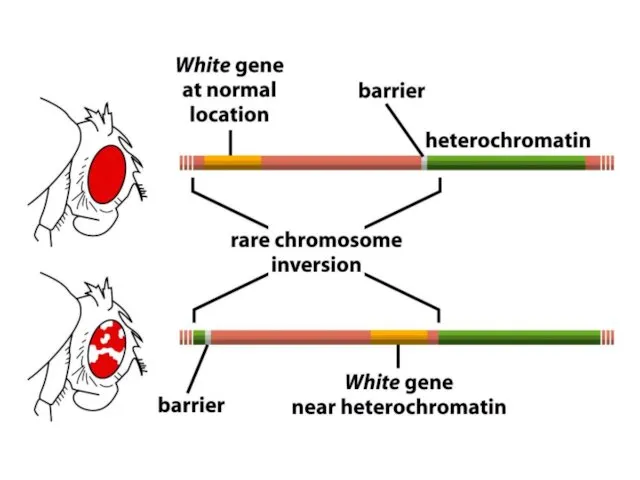
ДНҚ-ның гетерохроматин өңірлері, әдетте, транскрипцияланбайды және транскрипциялық реттеудің маңызды
Құбылыс әсері
ДНҚ-ның гетерохроматин өңірлері, әдетте, транскрипцияланбайды және транскрипциялық реттеудің маңызды

Көптеген ғалымдар нуклеосомаларды «бейтарап тұрғындар» деп санай отыра, эукариоттардағы гендердің реттелуі

Гистондардың гендердің реттелуі үшін маңызды емес деген көзқарасқа қарсы тұрудың екінші
Гистондардың гендердің реттелуі үшін маңызды емес деген көзқарасқа қарсы тұрудың екінші

Жасушалардың кейбір субпопуляциясында гендердің репрессияланған күйі тұрақты тұқымқуалауға жақын эухроматикалық аймақтардың

Дрозофила гетерохроматин аймағын эухроматин аймағына тікелей байланыстыратын, хромосомалық бұзылу жерлеріне жақын
Дрозофила гетерохроматин аймағын эухроматин аймағына тікелей байланыстыратын, хромосомалық бұзылу жерлеріне жақын

И.Ф. Жимулева зертханасында компактивтіліктің басталу орталықтарының айналасында белгілі бір тығыздауыш белоктарының
И.Ф. Жимулева зертханасында компактивтіліктің басталу орталықтарының айналасында белгілі бір тығыздауыш белоктарының


Х-хромосома инактивациясы
Гомологтық хромосомалардың және олардың аймақтарының дифференциалдық белсенділігі гендердің дифференциалды
Х-хромосома инактивациясы
Гомологтық хромосомалардың және олардың аймақтарының дифференциалдық белсенділігі гендердің дифференциалды


Лайон көрсеткіштеріне сәйкес:
1) гетеропикнотикалық Х хромосомасы бір жануардың әртүрлі жасушаларында
Лайон көрсеткіштеріне сәйкес:
1) гетеропикнотикалық Х хромосомасы бір жануардың әртүрлі жасушаларында
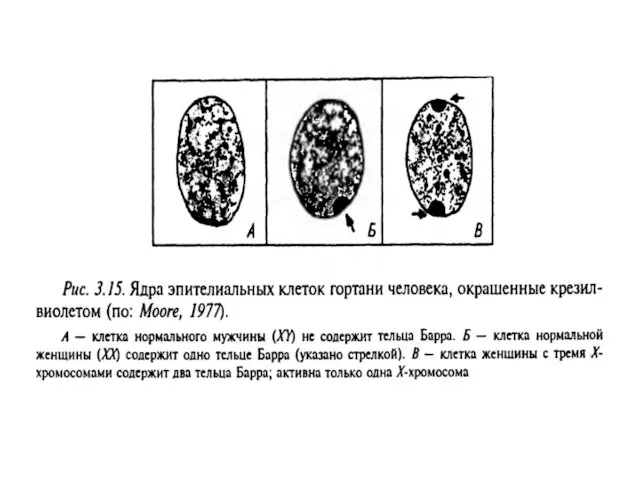

Х-хромосомалардың біреуі эмбриональдық дамудың ерте кезеңдерінде инактивтенеді.
Х-хромосомалардың екеуі де ооциттерде
Х-хромосомалардың біреуі эмбриональдық дамудың ерте кезеңдерінде инактивтенеді.
Х-хромосомалардың екеуі де ооциттерде

Адамдар мен макакалардың жыныстық хроматині 10-12 күндік бластоцистің трофобласт клеткаларында және
Адамдар мен макакалардың жыныстық хроматині 10-12 күндік бластоцистің трофобласт клеткаларында және

П. Рассел 1965 жылы. X хромосомасының инактивациялануы Xic (X-inactivation center) деп
П. Рассел 1965 жылы. X хромосомасының инактивациялануы Xic (X-inactivation center) деп

Х хромосомасын инактивациялаудың молекулалық механизмдерінің екі үлгісі ұсынылған.
1. Лайонизация басталмас бұрын,
Х хромосомасын инактивациялаудың молекулалық механизмдерінің екі үлгісі ұсынылған.
1. Лайонизация басталмас бұрын,

2. РНК-ның тұрақсыз нұсқасы лайонизация басталғанға дейін екі аллельмен де транскрибирленеді.
Х
Х

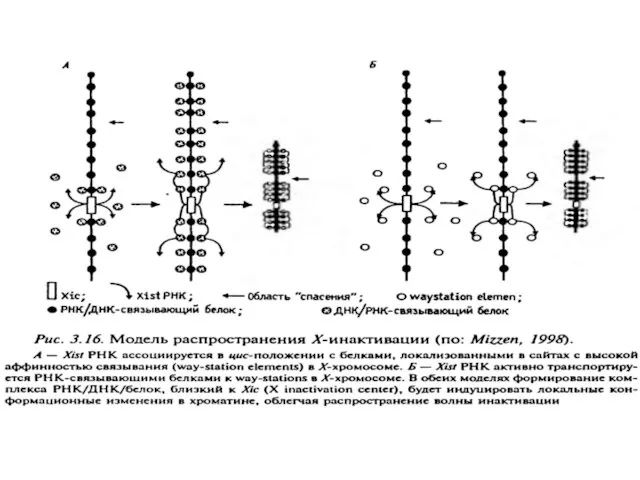
X хромосомасы бойынша инактивация қалай таралады?
Бірінші модель. Xist РНҚ хромосомада «цис-жылжыту»
X хромосомасы бойынша инактивация қалай таралады?
Бірінші модель. Xist РНҚ хромосомада «цис-жылжыту»

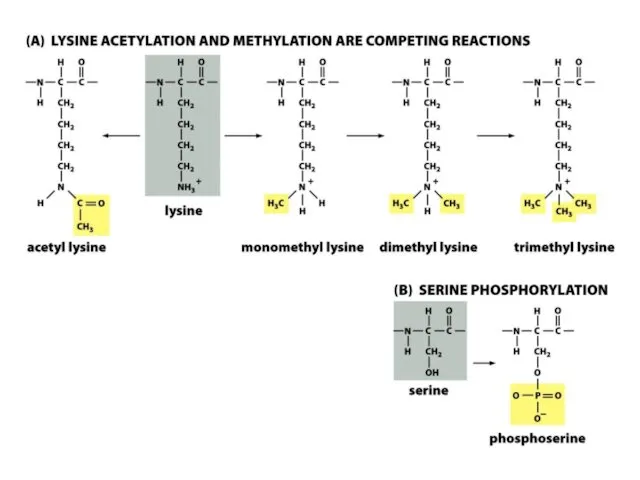
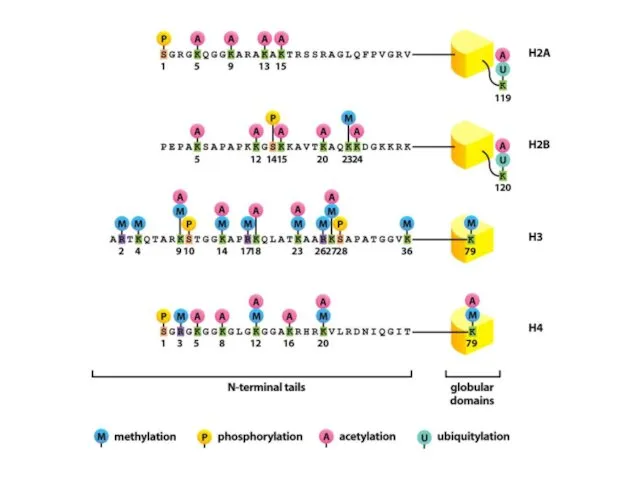
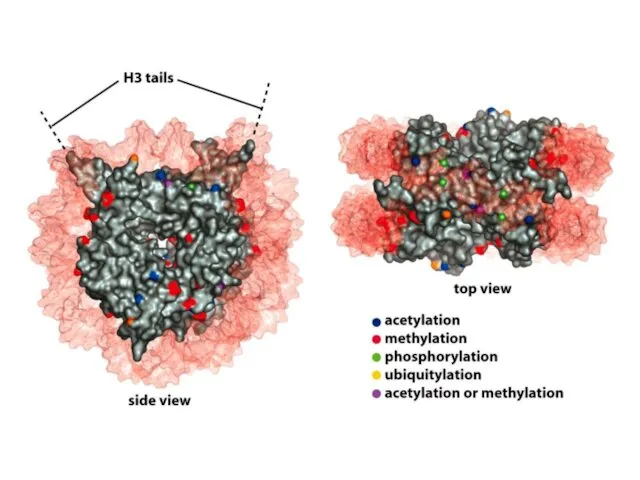
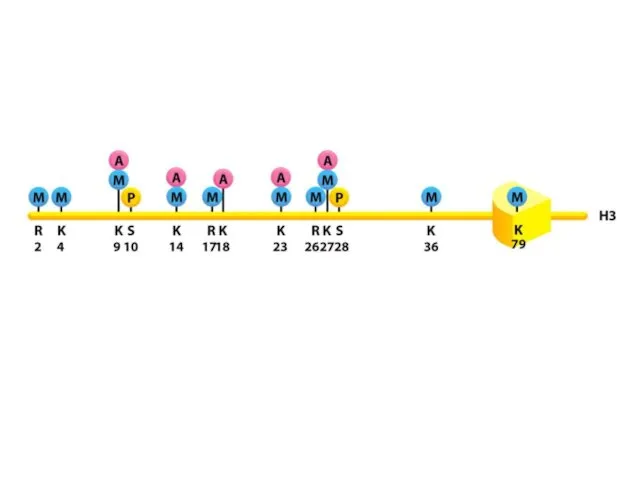
Нуклеосомалық ядроның барлық төрт аминқышқылдарының бүйірлік тізбектері, лизиндердің ацетилденуін, моно-, ди-
Нуклеосомалық ядроның барлық төрт аминқышқылдарының бүйірлік тізбектері, лизиндердің ацетилденуін, моно-, ди-


Нуклеосомадағы аминқышқылдардың модификациясы нақты ферменттермен жүзеге асырылады, бұл тек бір немесе
Нуклеосомадағы аминқышқылдардың модификациясы нақты ферменттермен жүзеге асырылады, бұл тек бір немесе

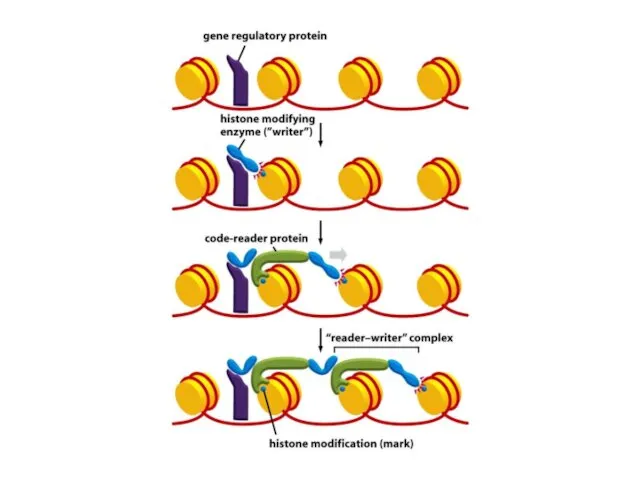
Бұл ферменттердің бастапқы «жиналуы» негізінен хромосомалардың ұзындығы бойынша кейбір ДНҚ тізбектерін
Бұл ферменттердің бастапқы «жиналуы» негізінен хромосомалардың ұзындығы бойынша кейбір ДНҚ тізбектерін

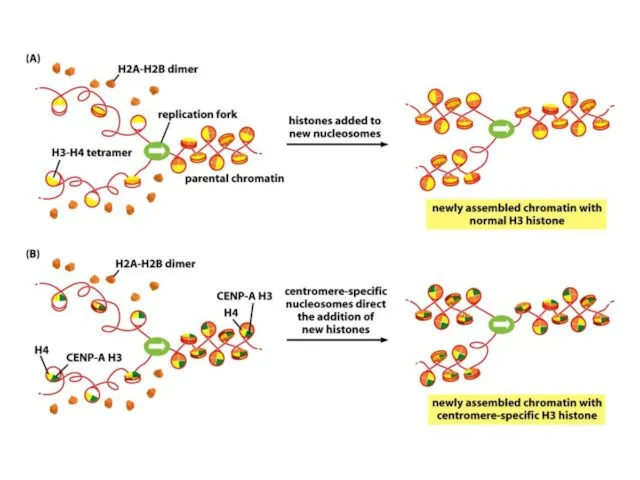
Негізгі гистондар көбінесе жасушалық циклдың S-фазасы кезінде синтезделеді және репликациялық шанышқыдан
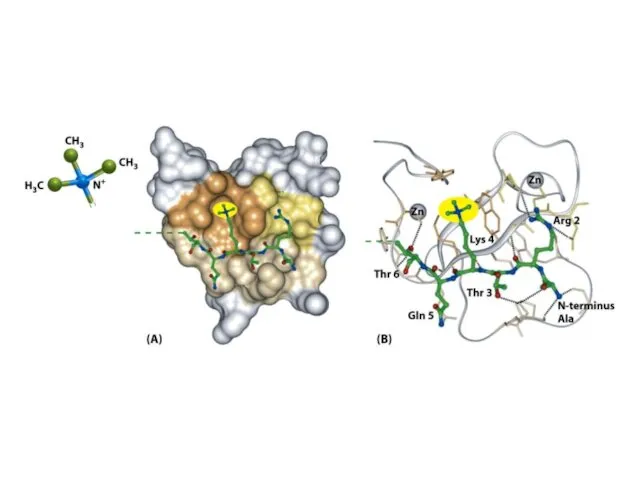
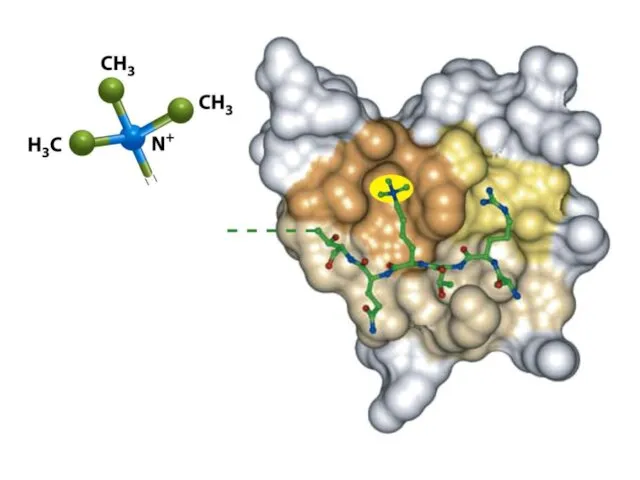
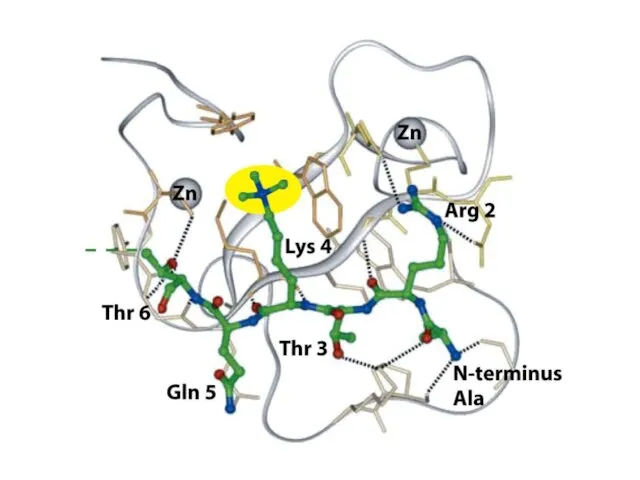

Шағын ақуыздық модульдер басқа модульдермен бірге жұмыс істеп, кодтық оқу кешені
Шағын ақуыздық модульдер басқа модульдермен бірге жұмыс істеп, кодтық оқу кешені
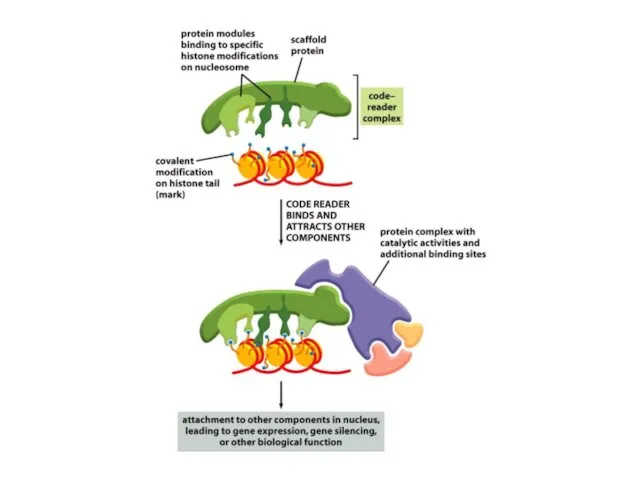

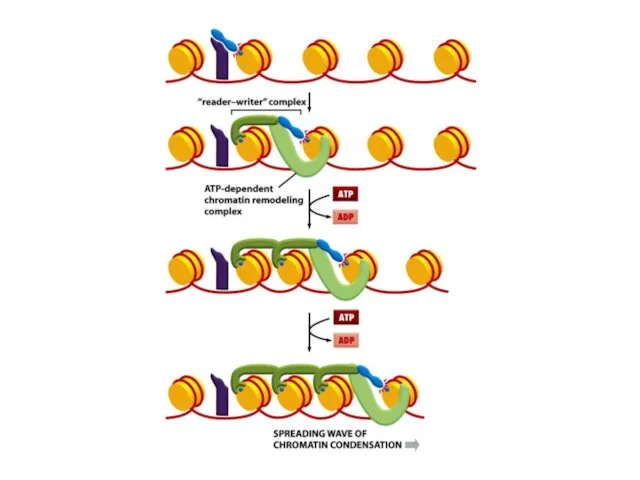
Мозаикалық әсердің құбылысы хроматиннің кейбір модифицирленген формаларының хромосомалық ДНҚ молекуласы бойында
Мозаикалық әсердің құбылысы хроматиннің кейбір модифицирленген формаларының хромосомалық ДНҚ молекуласы бойында
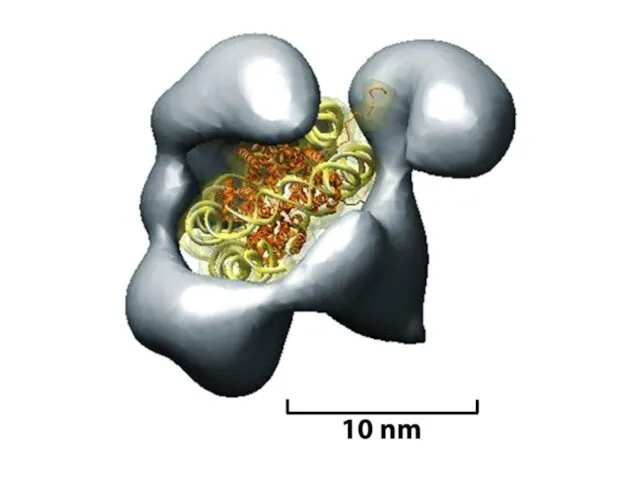

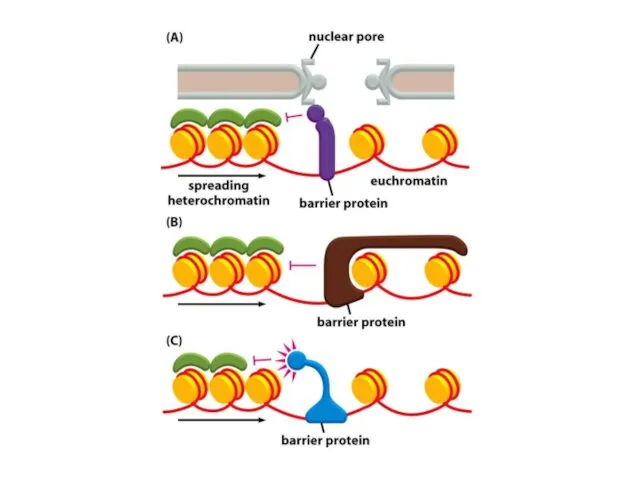
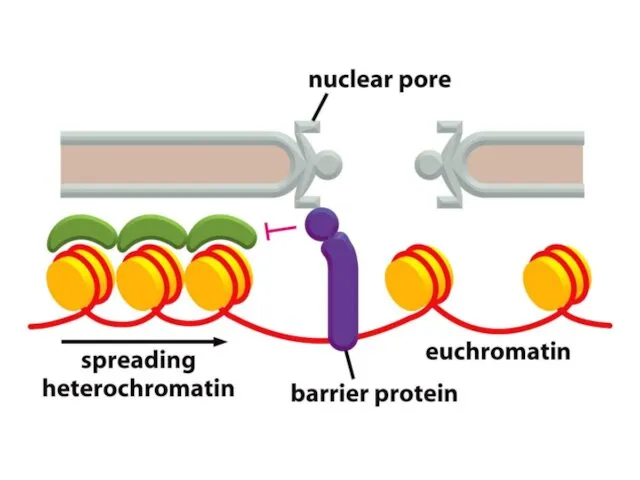
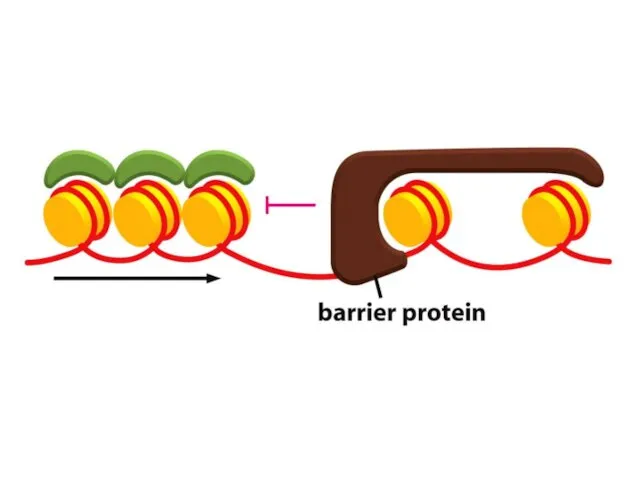
Қазіргі таңда ДНҚ тізбегінің кейбір аймақтарын хромосомаларға қосуға немесе шығаруға мүмкіндік
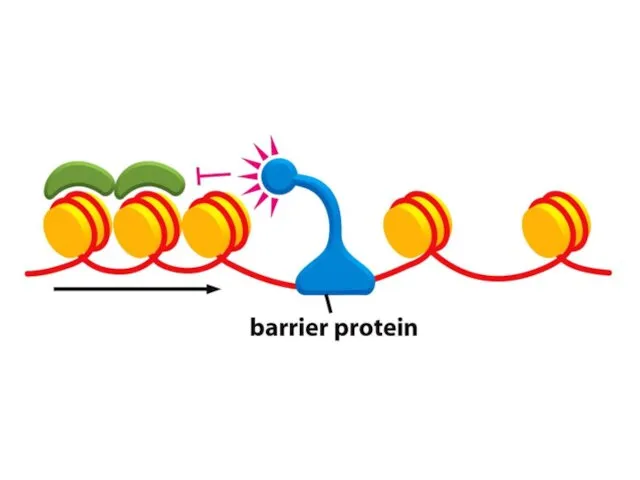
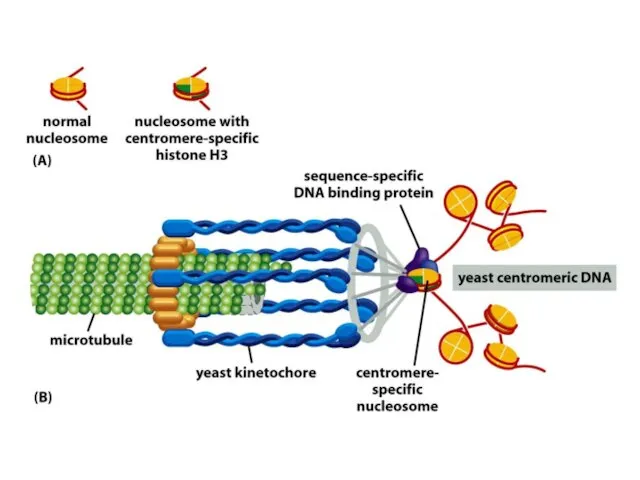

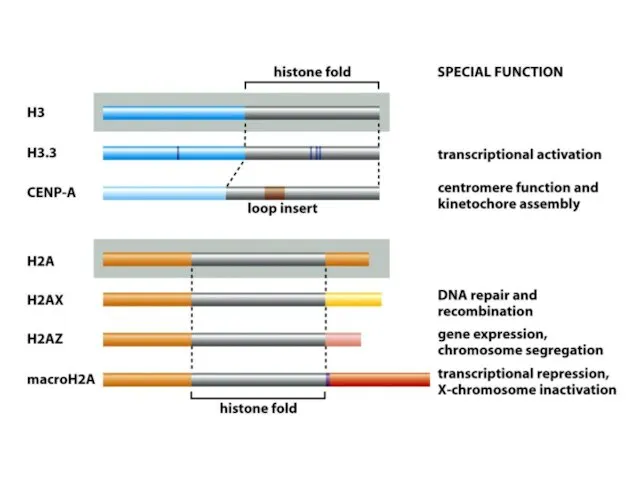
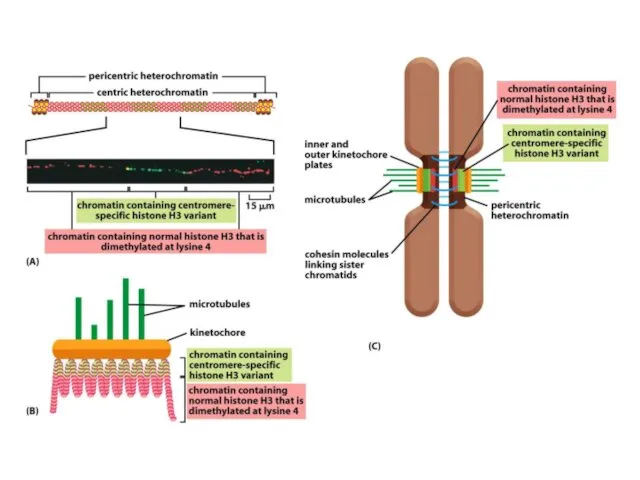
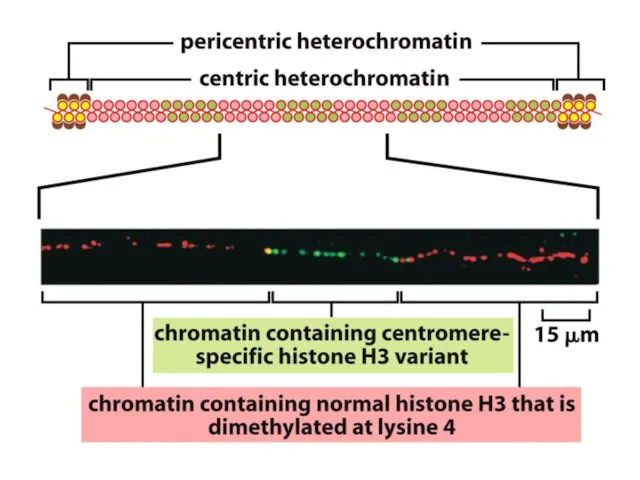
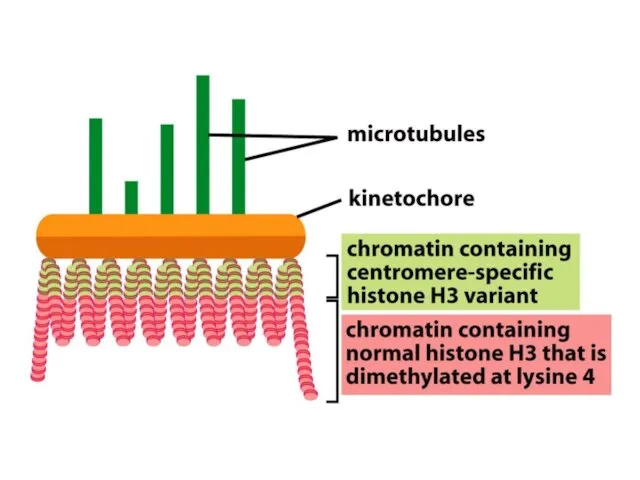
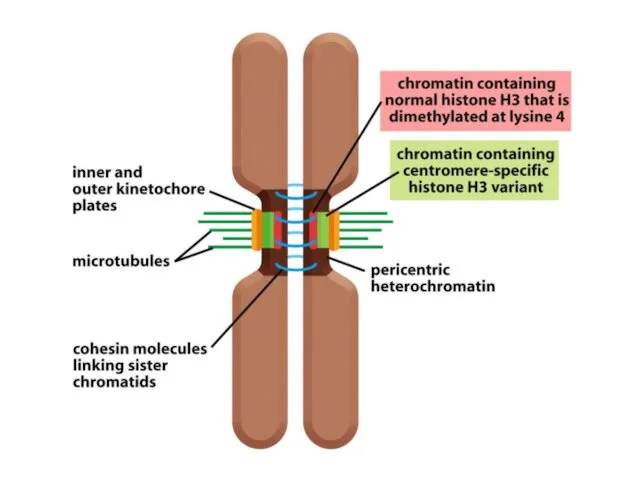
Көптеген күрделі ағзалардың, соның ішінде адамдардың әрбір центромері интерфазада сақталатын арнайы

Адамдар мен шыбындардың центромерлері жүздеген мың жұп нуклеотидтерге таралады және ДНҚ
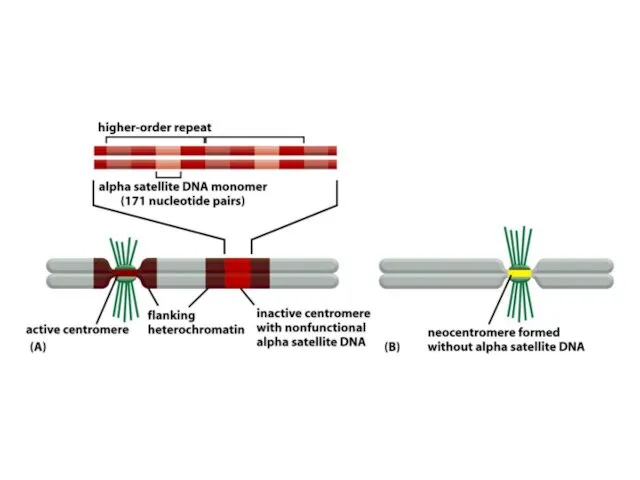
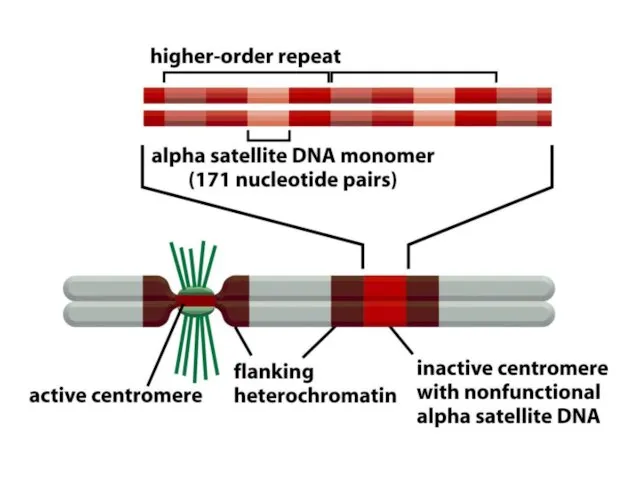
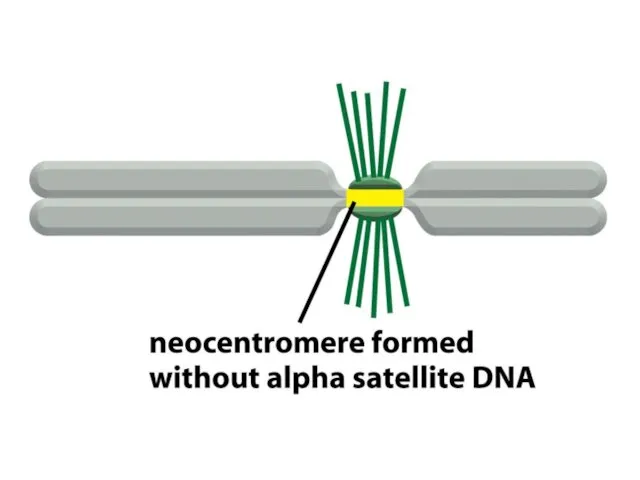

De novo центромерасының қалыптасуы бастапқы бастаушы нүктені талап етеді, оның мәні
De novo центромерасының қалыптасуы бастапқы бастаушы нүктені талап етеді, оның мәні
Гетерохроматиндер
Гетерохроматиндер
Эухроматиндер
Эухроматиндер
Гетерохроматиндер
Гетерохроматиндер
Эухроматиндер
Эухроматиндер
 Решение заданий 14 (С2) по материалам ЕГЭ профильного уровня (нахождение углов, расстояний, построение сечений)
Решение заданий 14 (С2) по материалам ЕГЭ профильного уровня (нахождение углов, расстояний, построение сечений) Балалардағы гельминтоз
Балалардағы гельминтоз Родительское собрание Мой ребёнок - левша
Родительское собрание Мой ребёнок - левша Where are the mice?
Where are the mice? Modelling and simulation
Modelling and simulation Химия р-элементов V группы. Общая характеристика элементов. Лекция 15
Химия р-элементов V группы. Общая характеристика элементов. Лекция 15 Классный час Права ребёнка
Классный час Права ребёнка 5 фактов из истории ролл мороженого
5 фактов из истории ролл мороженого Русский фольклор. Деревенский быт.
Русский фольклор. Деревенский быт. Мы памяти этой верны презентация к 70-летию победы
Мы памяти этой верны презентация к 70-летию победы Московский метрополитен
Московский метрополитен Презентация Как научить ребенка запоминать стихи
Презентация Как научить ребенка запоминать стихи Всестороннее обеспечение боя. Организация боевого обеспечения в мотострелковых подразделениях г. Астана 2015 год
Всестороннее обеспечение боя. Организация боевого обеспечения в мотострелковых подразделениях г. Астана 2015 год Биосфера - биологическая оболочка Земли
Биосфера - биологическая оболочка Земли Виды источников питания
Виды источников питания Мое право на образование
Мое право на образование Әнием, мин сине көтәрмен!
Әнием, мин сине көтәрмен! Для защиты (1)
Для защиты (1) Личный кабинет застрахованного
Личный кабинет застрахованного Бухгалтерский учет договоров аренды отдельными некредитными финансовыми организациями. Глава 13
Бухгалтерский учет договоров аренды отдельными некредитными финансовыми организациями. Глава 13 Понятие информационной системы. Модели информационных систем
Понятие информационной системы. Модели информационных систем Памяти наших героических прадедов и прабабушек посвящается…
Памяти наших героических прадедов и прабабушек посвящается… Олимпиала в Сочи 2014
Олимпиала в Сочи 2014 Компетентность личности – основа самореализации учащихся в образовательном процессе
Компетентность личности – основа самореализации учащихся в образовательном процессе Текстовые редакторы
Текстовые редакторы Скарлатина кезіндегі шаралардың стандарттары және алгоритмдері
Скарлатина кезіндегі шаралардың стандарттары және алгоритмдері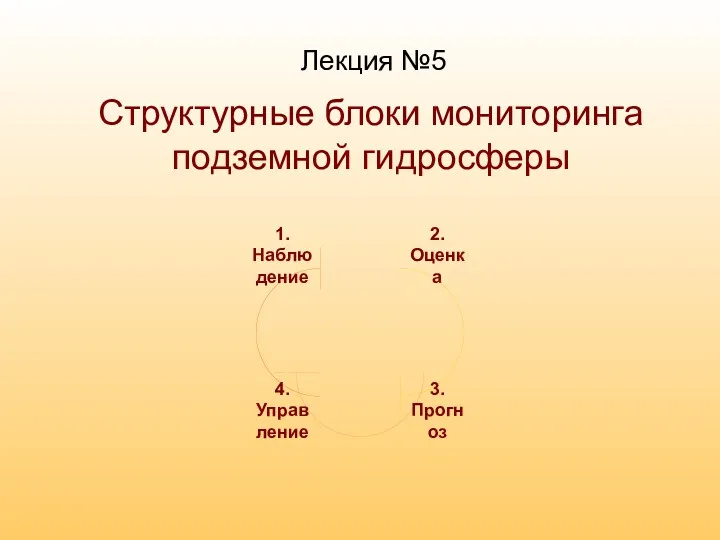 Структурные блоки мониторинга подземной гидросферы
Структурные блоки мониторинга подземной гидросферы Патологическое влечение к азартным играм
Патологическое влечение к азартным играм