- Биохимия зубов. Биохимия межклеточного матрикса

Содержание
- 2. ВВЕДЕНИЕ В БИОХИМИЮ ПОЛОСТИ РТА Гомеостаз полости рта во многом определяется структурно-функциональным состоянием тканей и микроорганизмов
- 3. ВИДЫ ТКАНЕЙ ПОЛОСТИ РТА
- 4. ОБЩИЙ ПЛАН СТРУКТУРНОЙ ОРГАНИЗАЦИИ ЗУБА
- 5. МЕЖКЛЕТОЧНЫЙ МАТРИКС Это комплекс органических и неорганических компонентов, заполняющий пространство между клетками. Для каждого вида тканей
- 6. Основными компонентами межклеточного матрикса являются: Различные виды коллагена, придающие тканям прочность; Неколлагеновые белки, преимущественно выполняющие функцию
- 7. ТИПЫ КОЛЛАГЕНОВ
- 8. ТИПЫ КОЛЛАГЕНОВ ПОЛОСТИ РТА Все типы коллагенов в зависимости от структуры делятся на: Фибриллообразующие Ассоциированные с
- 9. Структурной единицей коллагенового волокна является молекула тропоколлагена. Молекула тропоколлагена имеет толщину 1,5нм и длину 300нм. Каждая
- 10. ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА Синтез и созревание коллагена - сложный многоэтапный процесс, начинающийся в клетке,
- 11. Синтез полипептидных цепей коллагена Полипептидные цепи коллагена синтезируются на рибосомах, связанных с мембранами эндоплазматического ретикулума (ЭР),
- 12. Гидроксилазы пролина и лизина содержат в активном центре атом железа Fe 2+. Для сохранения атома железа
- 13. Образование проколлагена и его секреция в межклеточное пространство После гидроксилирования и гликозилирования каждая про-α-цепь соединяется водородными
- 14. СХЕМА СИНТЕЗА КОЛЛАГЕНА
- 15. Катаболизм коллагена Коллаген не постоянная молекула, со временем он разрушается. Его относят к медленно обменивающимся белкам;
- 16. Регуляция обмена коллагена Синтез коллагена регулируется разными способами. Сам коллаген и N-npo-пептиды после своего отщепления тормозят
- 17. НАРУШЕНИЯ ОБМЕНА КОЛЛАГЕНА Любые нарушения синтеза коллагена в первую очередь проявляется изменением со стороны зубочелюстной системы
- 18. Снижение процессов распада коллагена ведёт к фиброзу органов и тканей (в основном печени и лёгких). А
- 19. НЕКОЛЛАГЕНОВЫЕ БЕЛКИ ЭЛАСТИН Эластин – уникальный белок, главным свойством которого является возможность растягиваться в несколько раз
- 20. СТРУКТУРА ЭЛАСТИНА
- 21. СИНТЕЗ И РАСПАД ЭЛАСТИНА Синтез эластина начинается в фибробластах с образованием предшественника эластина – тропоэластина. Тропоэластин
- 22. НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА При снижении образования десмозинов (или их отсутствии) поперечные сшивкиобразуются в недостаточном количестве или
- 23. ГЛИКОЗАМИНОГЛИКАНЫ И ПРОТЕОГЛИКАНЫ Гликозаминогликаны (мукополисахариды) - линейные отрицательно заряженные гетерополисахариды, которые обнаруживаются в слизистых секретах (мукозах)
- 24. ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ они являются структурными компонентами межклеточного матрикса; протеогликаны и гликозаминогликаны специфически взаимодействуют с
- 25. СТРУКТУРА РАЗЛИЧНЫХ КЛАССОВ ГЛИКОЗАМИНОГЛИКАНОВ Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополисахаридов. Они построены из повторяющихся дисахаридных
- 26. СИНТЕЗ И РАЗРУШЕНИЕ ГЛИКОЗАМИНОГЛИКАНОВ 1.Синтез гликозаминогликанов Полисахаридные цепи гликозаминогликанов практически всегда связаны с белком, который называется
- 27. 2. Распад гликозаминогликанов Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2 ) многих из них
- 28. СТРОЕНИЕ И ВИДЫ ПРОТЕОГЛИКАНОВ Основной протеогликан хрящевого матрикса называется агрекан, он составляет 10% по весу исходной
- 29. Катаболизм агрекана изучен в настоящее время недостаточно. Имеются данные о наличии в хрящевом межклеточном матриксе фермента
- 30. НЕКОЛЛАГЕНОВЫЕ БЕЛКИ СОЕДИНИТЕЛЬНОЙ ТКАНИ Фибронектин Фибронектин – это гликопротеин экстрацеллюлярного матрикса, который синтезируется большинством клеток соединительной
- 31. СТРУКТУРА ФИБРОНЕКТИНА Рецептор фибронектина - интегрин, встроен в клеточную мембрану. Внутри клетки интегрин взаимодействует с актиновыми
- 32. ЛАМИНИН Ламинин – это гликопротеин, наиболее распространенный в базальных мембранах. Состоит из 3 полипептидных цепей, которые
- 33. НИДОГЕН Нидоген – этосульфатированный гликопротеин базальных мембран. Он состоит из одной полипептидной цепи, скрученной в 3
- 35. Скачать презентацию

ВВЕДЕНИЕ В БИОХИМИЮ ПОЛОСТИ РТА
Гомеостаз полости рта во многом определяется структурно-функциональным
ВВЕДЕНИЕ В БИОХИМИЮ ПОЛОСТИ РТА
Гомеостаз полости рта во многом определяется структурно-функциональным

ВИДЫ ТКАНЕЙ ПОЛОСТИ
РТА
ВИДЫ ТКАНЕЙ ПОЛОСТИ
РТА
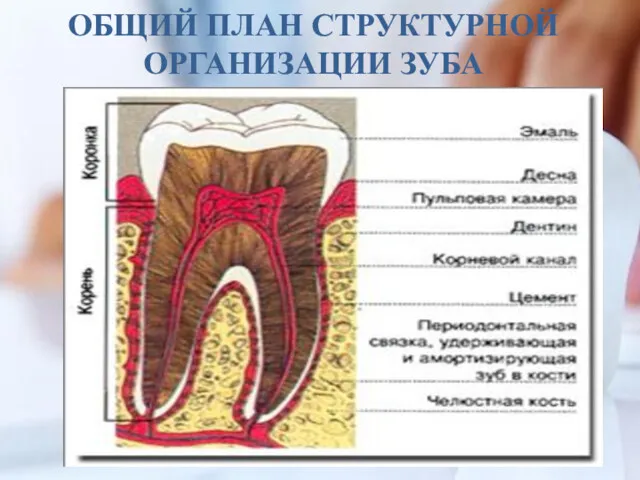
ОБЩИЙ ПЛАН СТРУКТУРНОЙ ОРГАНИЗАЦИИ ЗУБА
ОБЩИЙ ПЛАН СТРУКТУРНОЙ ОРГАНИЗАЦИИ ЗУБА

МЕЖКЛЕТОЧНЫЙ МАТРИКС
Это комплекс органических и неорганических компонентов, заполняющий пространство между клетками.
МЕЖКЛЕТОЧНЫЙ МАТРИКС
Это комплекс органических и неорганических компонентов, заполняющий пространство между клетками.

Основными компонентами межклеточного матрикса являются:
Различные виды коллагена, придающие тканям прочность;
Неколлагеновые белки,
Основными компонентами межклеточного матрикса являются:
Различные виды коллагена, придающие тканям прочность;
Неколлагеновые белки,

ТИПЫ КОЛЛАГЕНОВ
ТИПЫ КОЛЛАГЕНОВ

ТИПЫ КОЛЛАГЕНОВ ПОЛОСТИ РТА
Все типы коллагенов в зависимости от структуры делятся
ТИПЫ КОЛЛАГЕНОВ ПОЛОСТИ РТА
Все типы коллагенов в зависимости от структуры делятся

Структурной единицей коллагенового волокна является молекула тропоколлагена.
Молекула тропоколлагена имеет толщину 1,5нм
Структурной единицей коллагенового волокна является молекула тропоколлагена.
Молекула тропоколлагена имеет толщину 1,5нм

ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Синтез и созревание коллагена - сложный многоэтапный
ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Синтез и созревание коллагена - сложный многоэтапный

Синтез полипептидных цепей коллагена
Полипептидные цепи коллагена синтезируются на рибосомах, связанных с
Синтез полипептидных цепей коллагена
Полипептидные цепи коллагена синтезируются на рибосомах, связанных с

Гидроксилазы пролина и лизина содержат в активном центре атом железа Fe
Гидроксилазы пролина и лизина содержат в активном центре атом железа Fe

Образование проколлагена и его секреция в межклеточное пространство
После гидроксилирования и гликозилирования
Образование проколлагена и его секреция в межклеточное пространство
После гидроксилирования и гликозилирования
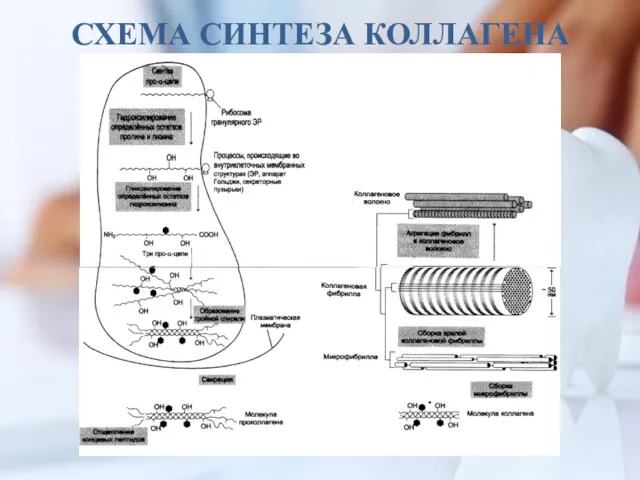
СХЕМА СИНТЕЗА КОЛЛАГЕНА
СХЕМА СИНТЕЗА КОЛЛАГЕНА

Катаболизм коллагена
Коллаген не постоянная молекула, со временем он разрушается. Его относят
Катаболизм коллагена
Коллаген не постоянная молекула, со временем он разрушается. Его относят

Регуляция обмена коллагена
Синтез коллагена регулируется разными способами.
Сам коллаген и N-npo-пептиды
Регуляция обмена коллагена
Синтез коллагена регулируется разными способами.
Сам коллаген и N-npo-пептиды

НАРУШЕНИЯ ОБМЕНА КОЛЛАГЕНА
Любые нарушения синтеза коллагена в первую очередь проявляется изменением
НАРУШЕНИЯ ОБМЕНА КОЛЛАГЕНА
Любые нарушения синтеза коллагена в первую очередь проявляется изменением

Снижение процессов распада коллагена ведёт к фиброзу органов и тканей (в
Снижение процессов распада коллагена ведёт к фиброзу органов и тканей (в

НЕКОЛЛАГЕНОВЫЕ БЕЛКИ
ЭЛАСТИН
Эластин – уникальный белок, главным свойством которого является возможность
НЕКОЛЛАГЕНОВЫЕ БЕЛКИ
ЭЛАСТИН
Эластин – уникальный белок, главным свойством которого является возможность
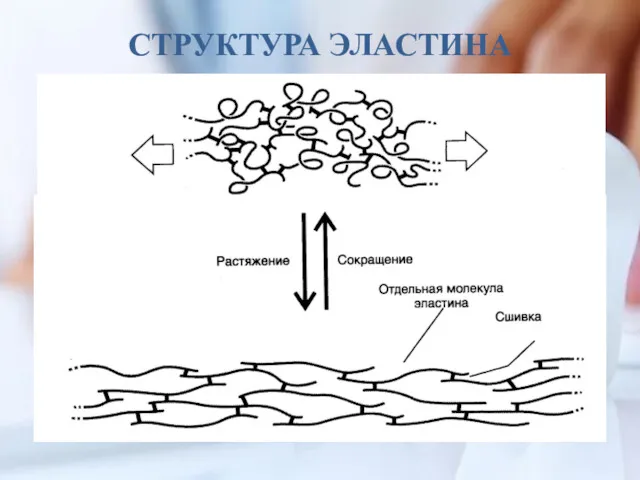
СТРУКТУРА ЭЛАСТИНА
СТРУКТУРА ЭЛАСТИНА

СИНТЕЗ И РАСПАД ЭЛАСТИНА
Синтез эластина начинается в фибробластах с образованием предшественника
СИНТЕЗ И РАСПАД ЭЛАСТИНА
Синтез эластина начинается в фибробластах с образованием предшественника

НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА
При снижении образования десмозинов (или их отсутствии) поперечные сшивкиобразуются
НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА
При снижении образования десмозинов (или их отсутствии) поперечные сшивкиобразуются

ГЛИКОЗАМИНОГЛИКАНЫ И ПРОТЕОГЛИКАНЫ
Гликозаминогликаны (мукополисахариды) - линейные отрицательно заряженные гетерополисахариды, которые обнаруживаются
ГЛИКОЗАМИНОГЛИКАНЫ И ПРОТЕОГЛИКАНЫ
Гликозаминогликаны (мукополисахариды) - линейные отрицательно заряженные гетерополисахариды, которые обнаруживаются

ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ
они являются структурными компонентами межклеточного матрикса;
протеогликаны и гликозаминогликаны
ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ
они являются структурными компонентами межклеточного матрикса;
протеогликаны и гликозаминогликаны

СТРУКТУРА РАЗЛИЧНЫХ КЛАССОВ ГЛИКОЗАМИНОГЛИКАНОВ
Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополисахаридов. Они
СТРУКТУРА РАЗЛИЧНЫХ КЛАССОВ ГЛИКОЗАМИНОГЛИКАНОВ
Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополисахаридов. Они

СИНТЕЗ И РАЗРУШЕНИЕ ГЛИКОЗАМИНОГЛИКАНОВ
1.Синтез гликозаминогликанов
Полисахаридные цепи гликозаминогликанов практически всегда связаны с
СИНТЕЗ И РАЗРУШЕНИЕ ГЛИКОЗАМИНОГЛИКАНОВ
1.Синтез гликозаминогликанов
Полисахаридные цепи гликозаминогликанов практически всегда связаны с

2. Распад гликозаминогликанов
Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2
2. Распад гликозаминогликанов
Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2

СТРОЕНИЕ И ВИДЫ ПРОТЕОГЛИКАНОВ
Основной протеогликан хрящевого матрикса называется агрекан, он составляет
СТРОЕНИЕ И ВИДЫ ПРОТЕОГЛИКАНОВ
Основной протеогликан хрящевого матрикса называется агрекан, он составляет
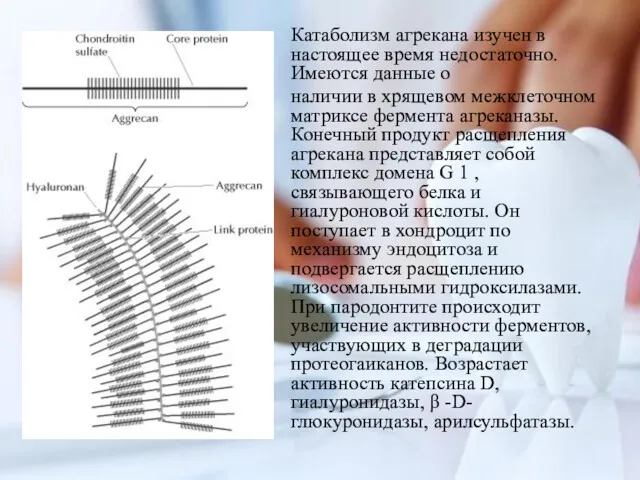
Катаболизм агрекана изучен в настоящее время недостаточно. Имеются данные о
наличии в
Катаболизм агрекана изучен в настоящее время недостаточно. Имеются данные о
наличии в

НЕКОЛЛАГЕНОВЫЕ БЕЛКИ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Фибронектин
Фибронектин – это гликопротеин экстрацеллюлярного матрикса, который синтезируется
НЕКОЛЛАГЕНОВЫЕ БЕЛКИ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Фибронектин
Фибронектин – это гликопротеин экстрацеллюлярного матрикса, который синтезируется
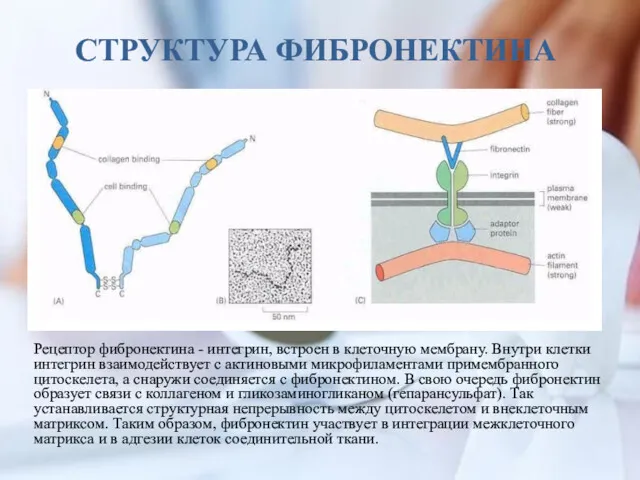
СТРУКТУРА ФИБРОНЕКТИНА
Рецептор фибронектина - интегрин, встроен в клеточную мембрану. Внутри клетки
СТРУКТУРА ФИБРОНЕКТИНА
Рецептор фибронектина - интегрин, встроен в клеточную мембрану. Внутри клетки
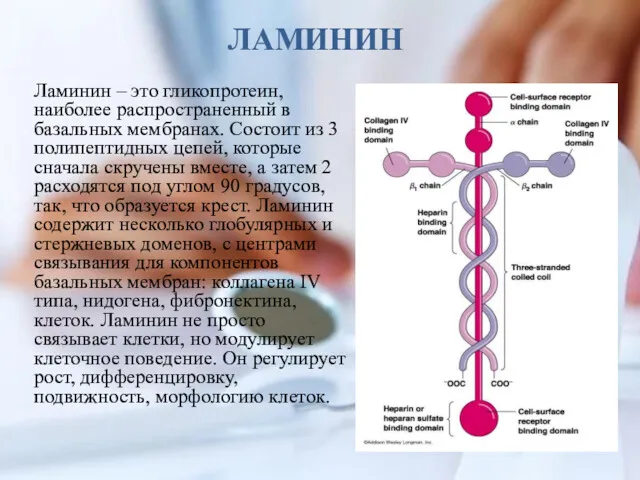
ЛАМИНИН
Ламинин – это гликопротеин, наиболее распространенный в базальных мембранах. Состоит из
ЛАМИНИН
Ламинин – это гликопротеин, наиболее распространенный в базальных мембранах. Состоит из

НИДОГЕН
Нидоген – этосульфатированный гликопротеин базальных мембран. Он состоит из одной полипептидной
НИДОГЕН
Нидоген – этосульфатированный гликопротеин базальных мембран. Он состоит из одной полипептидной
 Әлеуметтану ғылым ретінде. Әлеуметтану және медицина
Әлеуметтану ғылым ретінде. Әлеуметтану және медицина Воспитание личности заикающегося
Воспитание личности заикающегося Кровезаменители. Кровезамещающие жидкости
Кровезаменители. Кровезамещающие жидкости Физиология почки
Физиология почки Средства, влияющие на функции ЦНС. Психотропы и растительные гипнотики
Средства, влияющие на функции ЦНС. Психотропы и растительные гипнотики Укрепление здоровья населения. Современные проблемы профилактики
Укрепление здоровья населения. Современные проблемы профилактики Equipment and instruments of dental clinic(The Passive Voice)
Equipment and instruments of dental clinic(The Passive Voice) Секрет здоровья и долголетия
Секрет здоровья и долголетия Дамудың туа біткен ақауларының түрлері
Дамудың туа біткен ақауларының түрлері Определение газового состава крови. Исследование кислотно-щелочного равновесия
Определение газового состава крови. Исследование кислотно-щелочного равновесия Обследование больных с патологией сердечно-сосудистой системы. Расспрос, осмотр, пальпация сердечной области. (Лекция 6)
Обследование больных с патологией сердечно-сосудистой системы. Расспрос, осмотр, пальпация сердечной области. (Лекция 6) Детский реабилитационно-восстановительный центр. Базовая программа реабилитации
Детский реабилитационно-восстановительный центр. Базовая программа реабилитации Фізіологічне спостереження за розвитком плоду, перебігом вагітності. Біофізичний профіль плода
Фізіологічне спостереження за розвитком плоду, перебігом вагітності. Біофізичний профіль плода Переломы нижней челюсти. Клиническая картина, диагностика и лечение
Переломы нижней челюсти. Клиническая картина, диагностика и лечение Жүйкелік анорексия
Жүйкелік анорексия Заболевания глотки. Методы исследования
Заболевания глотки. Методы исследования Патологическая анатоми заболеваний почек
Патологическая анатоми заболеваний почек Захворювання печінки, жовчних шляхів, підшлункової залози. Стоматити. Гельмінтози
Захворювання печінки, жовчних шляхів, підшлункової залози. Стоматити. Гельмінтози Изготовление порошков с веществами списка А и Б, тритурации
Изготовление порошков с веществами списка А и Б, тритурации Закаливание детей в ДОУ
Закаливание детей в ДОУ Неотложные состояния в педиатрии
Неотложные состояния в педиатрии Виды кровотечений. Методы остановки кровотечений
Виды кровотечений. Методы остановки кровотечений Фізіологічні особливості організму дітей середнього і старшого шкільного віку та їх адаптпція до фізичних навантажень
Фізіологічні особливості організму дітей середнього і старшого шкільного віку та їх адаптпція до фізичних навантажень Классификация аномалий развития женских половых органов, диагностика
Классификация аномалий развития женских половых органов, диагностика Раневая инфекция
Раневая инфекция Крок для студентов медицинских вузов
Крок для студентов медицинских вузов Тұқым қуалайтын ауруларды емдеудің негізгі принциптері
Тұқым қуалайтын ауруларды емдеудің негізгі принциптері Миофасциальный болевой синдром лица
Миофасциальный болевой синдром лица