- Боль, условные рефлексы

Содержание
- 2. Физиологическое значение боли Боль представляет собой неприятное ощущение, реализующееся специальной системой болевой чувствительности и высшими отделами
- 3. Психофизиология боли Боль - это не только и даже не столько сенсорная модальность, но и ощущение,
- 4. Классификация боли 1. Физиологическая и патологическая 2. Острая и хроническая 3. Поверхностная, глубокая, 4. Соматическая, висцеральная,
- 5. Классификация боли По характеру, течению, качеству и субъективным ощущениям различают боли : приступообразные, постоянные, молниеносные, тупые,
- 7. Типы боли (розовый фон) и области происхождения каждого из них (серый фон) с примерами специфических форм
- 9. Компоненты боли, активируемые вредными стимулами. Участие сенсорного, афферентного и вегетативного компонентов в оценке (когнитивный аспект) и
- 10. Структура и локализация кожных рецепторов
- 11. Точки на коже человека, воспринимающие боль и давление (внутренняя сторона предплечья). Положение болевых точек определено с
- 12. Структуры и субстраты, обусловливающие ноцицептивные боли. Вверху–последовательность стадий возникновения боли: посередине –этапы обработки информации. Внизу показано,
- 13. Характеристика алгогенных агентов Боль возникает не только в результате раздражения специальных ноцицепторов, но и вследствие избыточной
- 14. Алгогенные химические агенты а) тканевые (серотонин, гистамин, ацетилхолин, простагландины, ионы К и Н); б) плазменные (брадикинин,
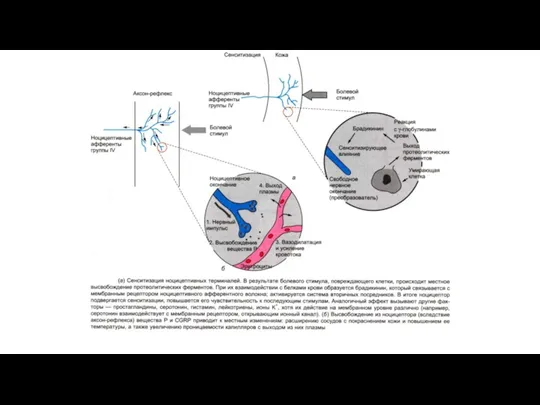
- 16. Рис. Мембранно-ионные механизмы сенсинга боли при действии химических и физических раздражителей. Ноцицепторы используют различные ионные каналы
- 17. Диаграмма, иллюстрирующая факторы, воздействующие на периферические ноцицептивные волокна, которые отвечают за их разряд ???? И сенситизацию
- 19. Диаграмма, показывающая основные взаимодействия сенсерного нейрона – сателитнат глиальной клетки в спинальном . Тройничном ганглии во
- 21. Восходящие пути болевой чувствительности. Лемнисковые и экстралемнисковые восходящие системы. В пределах спинного мозга одна из них
- 25. Схема, демонстрирующая восходящие ноцицептивные пути (слева) и системы нисходящих трактов, модулирующих ноцицептивные сигналы (справа). Из восходящих
- 27. Пути возникновения отраженной боли. Слева: некоторые из ноцицептивных афферентов от внутренних органов оканчиваются в заднем роге
- 28. Схема возникновения проецируемой боли
- 30. Основной соматосенсорный путь от лицевой поверхности и полости рта. Тригеминальные нервные тройничные окончания
- 31. Неврологические нарушения после односторонней перерезки спинного мозга (синдром Броун-Секара)
- 32. Блок-схема проводящих путей болевой чувствительности
- 33. Центральные структуры, участвующие в болевой чувствительности («болевой матрикс»). Th – таламус, Amyg – амигдала, Insula -
- 34. Структуры и механизмы интеграции боли (ноцицептивная система). Ретикулярная формация - В РФ продолговатого мозга существуют нейроны,
- 36. Антиноцицептивная система Ноцицептивная система имеет свой функциональный антипод - антиноцицептивную систему, которая контролирует деятельность структур ноцицептивной
- 37. Структура антиноцицептивнойсистемы Антиноцицептивная система представлена сегментарным и центральным уровнями контроля, а также гуморальными механизмами (системами): -
- 38. При возникновении боли обязательно происходит активация антиноцицептивной системы. 1. На уровне ствола мозга включается нисходящая аналгетическая
- 39. Опиатные механизмы обезболивания. Опиатные рецепторы мозга связываются с веществами типа морфина или его синтетическими аналогами, а
- 40. Различные области ЦНС имеют неодинаковую чувствительность эндорфинам и энкефалинам. Например, гипофиз в 40 раз чувствительнее к
- 43. Эндогенные опиаты. Вещества выделенные из мозга животных, обладают способностью соединяться с опиатными рецепторами. Они имеют структуру
- 44. Морфин – антиболевые пути
- 46. Механизм обезболивающего действия опиатов Опиаты соединяются с ноцицепторами и препятствуют соединению с ними нейротрансмиттера (субстанции P).
- 49. Адренэргические механизмы обезболивания. Норадреналин, через с α-адренорецепторы тормозит проведение ноцицептивных импульсов на всех уровнях мозга. При
- 53. АЛЬТЕРНАТИВНЫЕ механизмы обезболивания. Вазопрессин, ангиотензин, окситоцин, соматостатин, нейротензин оказывают аналгетический эффект без активации опиатной системы. Доказано,
- 55. ОСТРАЯ БОЛЬ "ВОРОТНАЯ" ТЕОРИЯ Р. МЕЛЬЗАКА И П. УОЛЛА. Роль ворот, пропускающих ноцицептивные импульсы вверх, играют
- 56. ХРОНИЧЕСКАЯ БОЛЬ При длительном повреждении тканей (воспаление, переломы, опухоли и т.д.) постоянная болевая информация, вызывая резкую
- 57. Фантомные боли Фантомные боли (боли в ампутированных конечностях) объясняются в основном дефицитом афферентной информации, в результате
- 58. Отраженная боль. Ее возникновение связано с тем, что афференты внутренних органов и кожи связаны с одними
- 61. Положение зон Геда внутренних органов и их соотношение с дерматомами
- 63. Механизм отраженных болей
- 66. Нарушения функций организма при боли. Нарушение сна, сосредоточенности, полового влечения, повышенная раздражительность. При хронической интенсивной боли
- 67. Нарушения функций организма при боли. Биохимические изменения проявляются в виде повышения потребления кислорода, распада гликогена, гипергликемии,
- 68. Болевая рецепция полости рта. Выраженной болевой чувствительностью обладает часть слизистой оболочки на вестибулярной поверхности нижней челюсти
- 72. Виды обезболивания. Наркоз (ингаляционный и внутривенный Проводниковая анестезия. Обезболивание охлаждением. Электрообезболивание. Обезболивание иглоукалыванием. Аудиоанестезия. Гипноз.
- 74. Главные фармакологические, физические и психологические методы облегчения боли
- 75. Физиология высшей нервной деятельности
- 76. Цель лекции: Дать характеристику условно-рефлекторной деятельности человека Мотивация: Данный раздел физиологии представляет интерес для клинической практики
- 77. План лекции: 1. Общая характеристика высшей нервной деятельности животных и человека 2. Методы исследования головного мозга
- 78. ВЕХИ ИСТОРИИ 1637 — Р. Декарт (R. Descartes, Франция) дал первое описание двигательного рефлекса. 1863 —
- 83. СОЗНАНИЕ Сознание - свойство мозга - высшее проявление функции отражения действительности. Сознание - способность субъективного отражения
- 88. Смена уровней поведения в эволюции Врожден-ные Приобре-тенные
- 89. Уровни активации психической деятельности Эмоции Внимание Спокойное бодрствование Сон Уровни активации нервной системы Поведение Аффект Бодрствование
- 90. Таксисы предполагают избирательные реакции приближения к источнику света ( фототаксис), химических веществ ( хемотаксис) и т.д.
- 91. Безусловные рефлексы Врожденные, свойственные всем особям данного вида Имеют готовую генетически детерминированную рефлекторную дугу Созревают постепенно
- 92. Витальные инстинкты Пищевой Питьевой Оборонительный (активный -ястреб и пассивный - кролик) Регулирования цикла «сон-бодрствование» Экономии энергии
- 93. Ролевые инстинкты Половые - выбор партнера Родительские - разделение ролей отца и матери Территориальные - охрана
- 94. Инстинкты саморазвития Исследовательский Новизны Свободы Имитационный (подражательный) Игровой
- 95. Отличия условного и безусловного рефлексов: условные безусловные приобретенные врожденные временные постоянные индивидуальные видовые вырабатываются на осуществляются
- 96. Выработка условного рефлекса по И.П.Павлову Фистула для сбора слюны Звонок (условный раздражитель) Пища (безусловный раздражитель)
- 98. Инструментальный условный рефлекс Педаль Кормушка
- 101. Основные характеристики условного рефлекса (по И.П.Павлову) 1) Приобретаемость условных рефлексов (врожденность безусловных рефлексов) 2) Индивидуальность условного
- 103. Образование временной связи по пути «кора-кора» по И.П.Павлову
- 105. Образование временной связи путем конвергенции на нейроне по П.К.Анохину
- 107. Классификация условных рефлексов По происхождению - натуральные и искусственные По характеру безусловного подкрепления - пищевые, оборонительные,
- 108. Условия выработки условных рефлексов Условие времени - предварительность или одновременность действия условного и безусловного раздражителей Условие
- 109. Механизм замыкания временной связи Теория двух корковых очагов И.П.Павлова - проторение пути Теория корково-подкорковых связей Теория
- 110. а) наличные совпадающие короткоотставленные отставленные подкрепление через 2-4 с через 20-30 с б) следовые короткоотставленные поздние
- 111. Правила выработки условного рефлекса: ∙ вырабатывается на базе безусловного ∙ необходимо неоднократное сочетание индифферентного и безусловного
- 114. Схемы образования условного рефлекса по И. П. Павлову (А) и Э. А. Асратяну (Б) К —
- 115. Дуга условного рефлекса с двусторонней связью А, Б — корковые представительства мигательного и пищевого рефлексов; В,
- 116. Взаимодействие синаптических входов с гетерохимической организацией для стимулов биологической (а) и сенсорных модальностей. Различные свойства входов
- 117. Передача условной связи, сложившейся в процессе сочетаний, с входного нейрона (А) на «выходной» (Б) по одному
- 118. Аксошипиковый транзиторный синапс 1 — аксон, 2 — синаптические везикулы, 3 — шипиковый аппарат, 4 —
- 119. Основные типы нейронных объединений в лобной коре А — нейронная «ловушка»; Б — вертикальный модуль
- 121. Представление об иммунохимическом механизме формирования памяти (по Ашмарину) КБ — катионные белки, открывающие путь комплексам синаптического
- 126. Безусловное торможение: внешнее торможение (протекает по типы постоянного и гаснущего тормоза), запредельное торможение (развивается при большой
- 127. Внешнее торможение
- 128. Угасательное торможение звонок пища звонок звонок
- 129. Запаздывающее торможение звонок пища пища пища звонок звонок
- 130. Дифференцировочное торможение
- 131. Сигнальное торможение
- 132. Образование угасательного торможения А — условный пищевой рефлекс; Б — стадия частичного торможения пищевой реакции вследствие
- 134. Связи нервных клеток, объясняющие происхождение тормозных условных нейронных ответов 1 — пирамидная клетка глубоких слоев коры,
- 139. Скачать презентацию

Физиологическое значение боли
Боль представляет собой неприятное ощущение, реализующееся специальной системой
Физиологическое значение боли
Боль представляет собой неприятное ощущение, реализующееся специальной системой

Психофизиология боли
Боль - это не только и даже не столько сенсорная
Психофизиология боли
Боль - это не только и даже не столько сенсорная

Классификация боли
1. Физиологическая и патологическая
2. Острая и хроническая
3. Поверхностная, глубокая,
4. Соматическая,
Классификация боли
1. Физиологическая и патологическая
2. Острая и хроническая
3. Поверхностная, глубокая,
4. Соматическая,

Классификация боли
По характеру, течению, качеству и субъективным ощущениям различают боли :
Классификация боли
По характеру, течению, качеству и субъективным ощущениям различают боли :
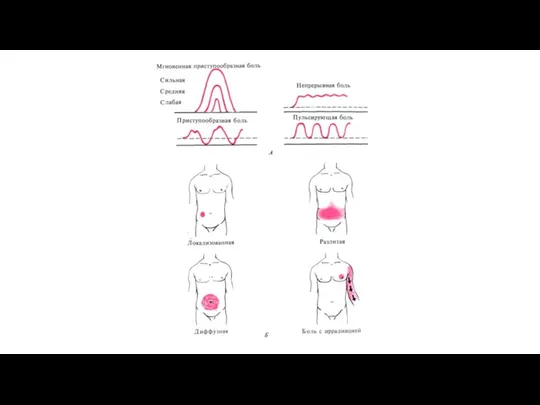
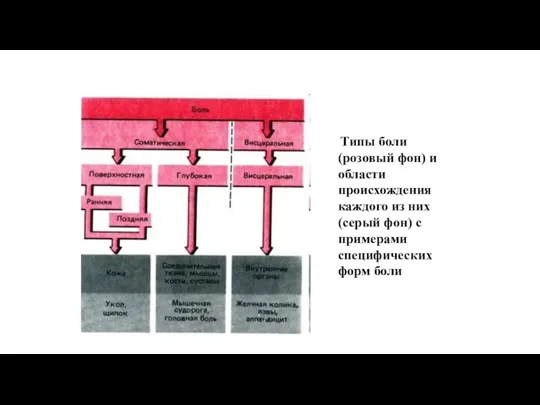
Типы боли (розовый фон) и области происхождения каждого из них
Типы боли (розовый фон) и области происхождения каждого из них
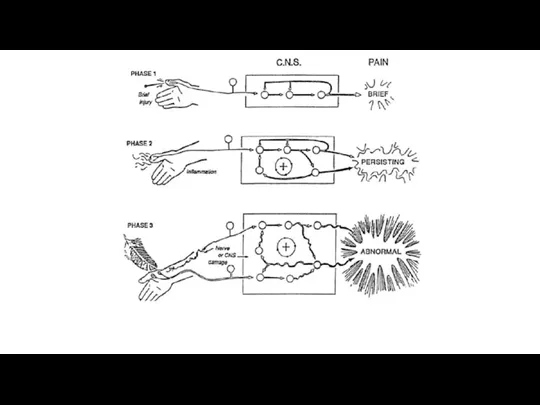
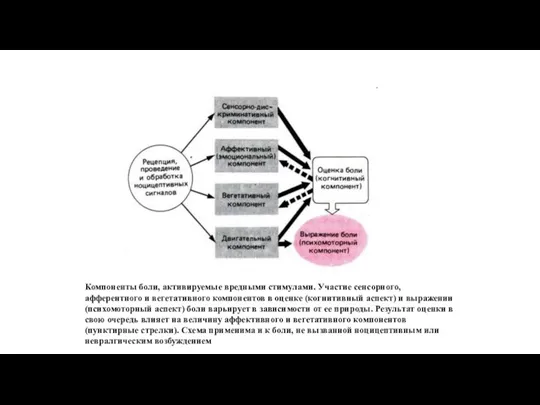
Компоненты боли, активируемые вредными стимулами. Участие сенсорного, афферентного и вегетативного компонентов
Компоненты боли, активируемые вредными стимулами. Участие сенсорного, афферентного и вегетативного компонентов
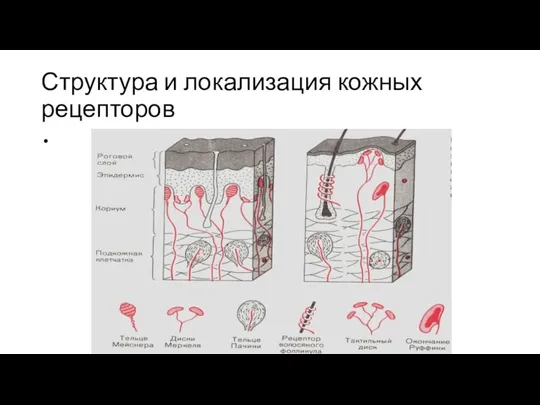
Структура и локализация кожных рецепторов
Структура и локализация кожных рецепторов

Точки на коже человека, воспринимающие боль и давление (внутренняя сторона предплечья).
Точки на коже человека, воспринимающие боль и давление (внутренняя сторона предплечья).
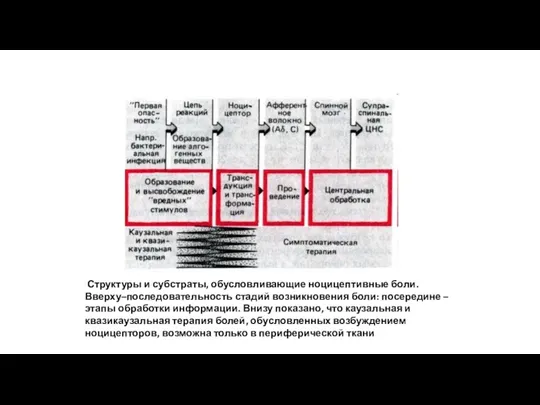
Структуры и субстраты, обусловливающие ноцицептивные боли. Вверху–последовательность стадий возникновения боли:
Структуры и субстраты, обусловливающие ноцицептивные боли. Вверху–последовательность стадий возникновения боли:

Характеристика алгогенных агентов
Боль возникает не только в результате раздражения специальных
Характеристика алгогенных агентов
Боль возникает не только в результате раздражения специальных

Алгогенные химические агенты
а) тканевые (серотонин, гистамин, ацетилхолин, простагландины, ионы К и
Алгогенные химические агенты
а) тканевые (серотонин, гистамин, ацетилхолин, простагландины, ионы К и
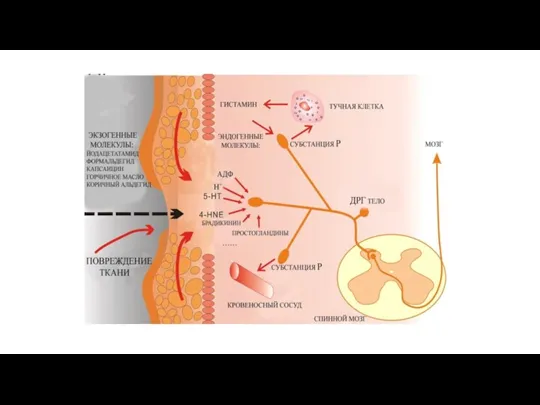
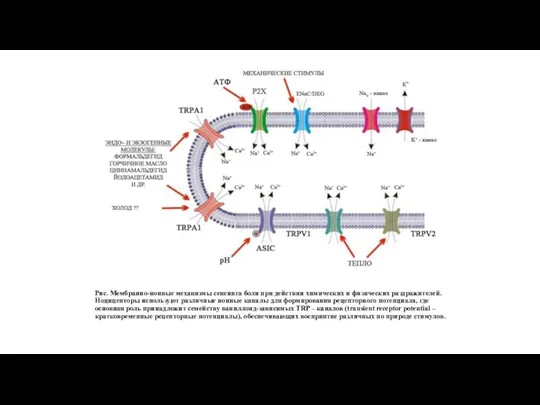
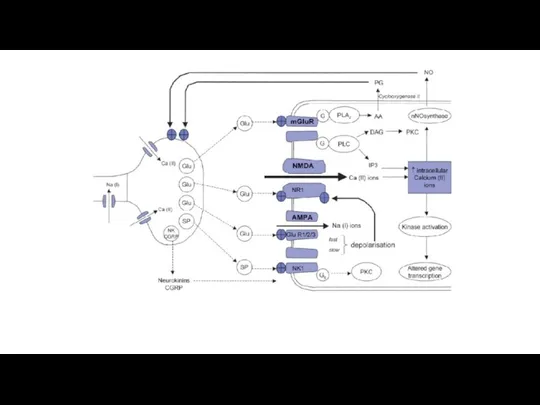
Рис. Мембранно-ионные механизмы сенсинга боли при действии химических и физических раздражителей.
Рис. Мембранно-ионные механизмы сенсинга боли при действии химических и физических раздражителей.
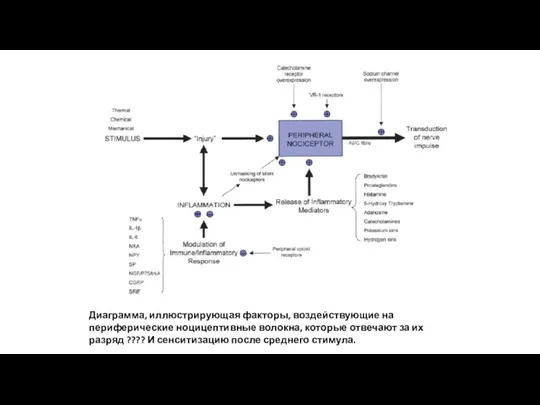
Диаграмма, иллюстрирующая факторы, воздействующие на периферические ноцицептивные волокна, которые отвечают за
Диаграмма, иллюстрирующая факторы, воздействующие на периферические ноцицептивные волокна, которые отвечают за
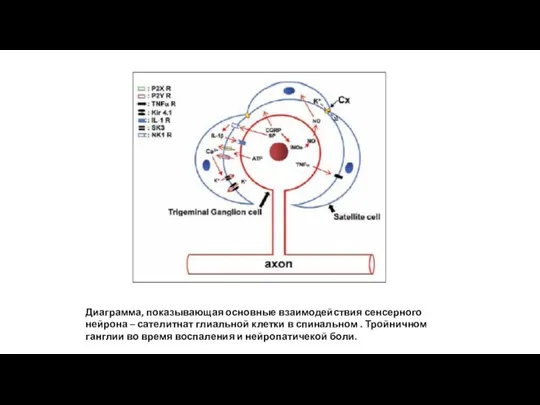
Диаграмма, показывающая основные взаимодействия сенсерного нейрона – сателитнат глиальной клетки в
Диаграмма, показывающая основные взаимодействия сенсерного нейрона – сателитнат глиальной клетки в

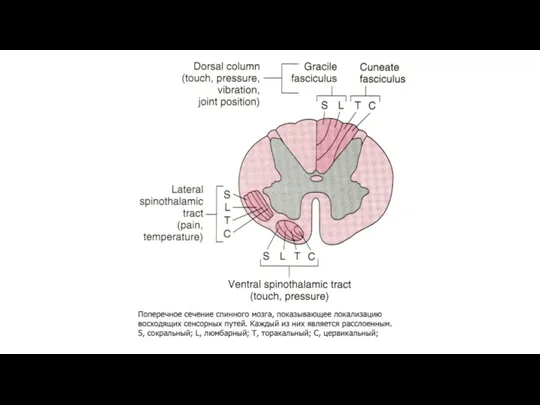
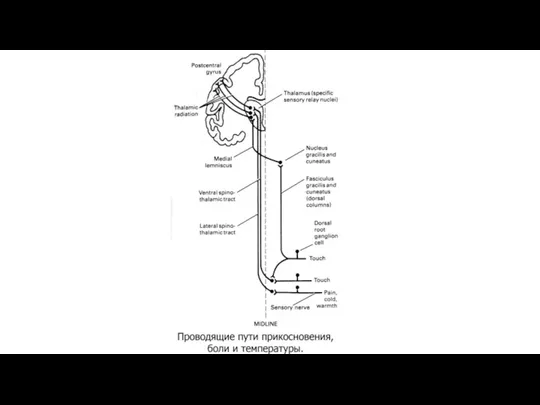
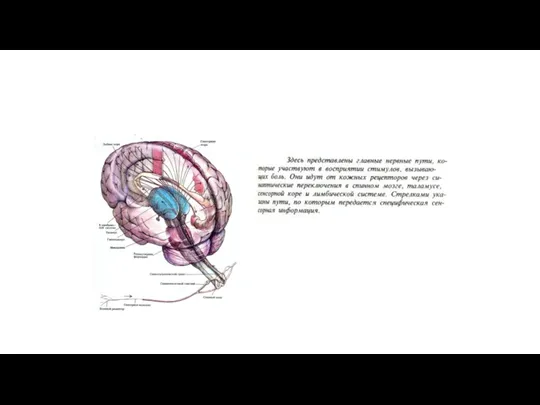
Восходящие пути болевой чувствительности.
Лемнисковые и экстралемнисковые восходящие системы.
В
Восходящие пути болевой чувствительности.
Лемнисковые и экстралемнисковые восходящие системы.
В

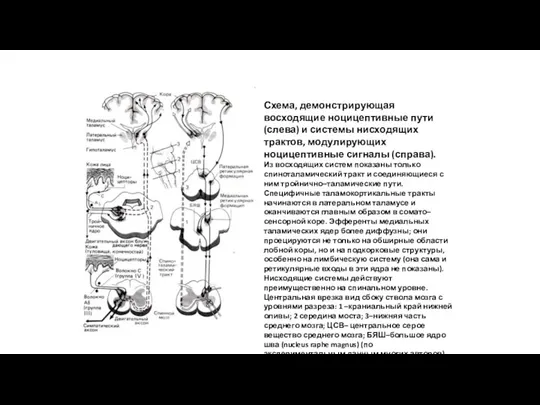
Схема, демонстрирующая восходящие ноцицептивные пути (слева) и системы нисходящих трактов, модулирующих
Схема, демонстрирующая восходящие ноцицептивные пути (слева) и системы нисходящих трактов, модулирующих
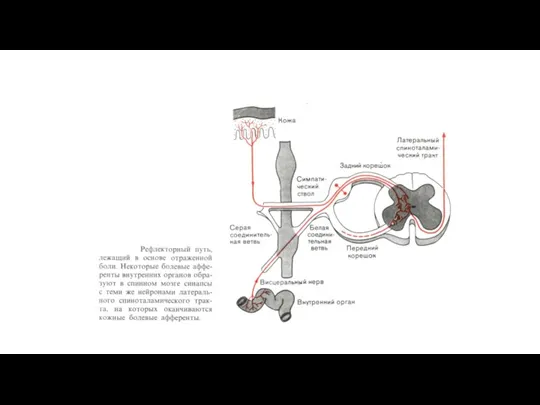
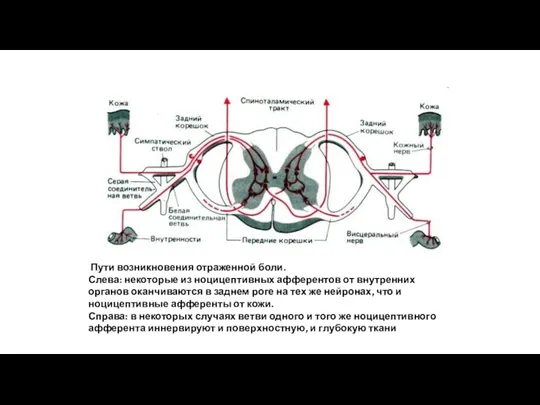
Пути возникновения отраженной боли.
Слева: некоторые из ноцицептивных афферентов от
Пути возникновения отраженной боли.
Слева: некоторые из ноцицептивных афферентов от
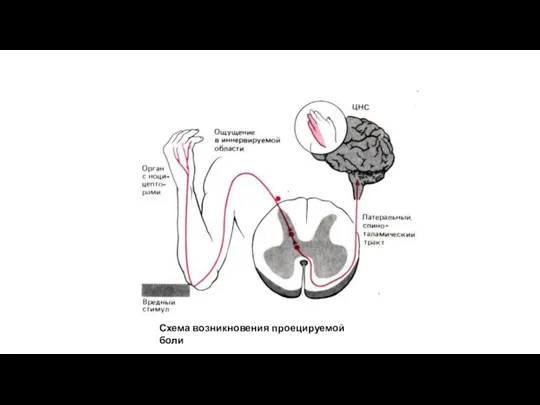
Схема возникновения проецируемой боли
Схема возникновения проецируемой боли
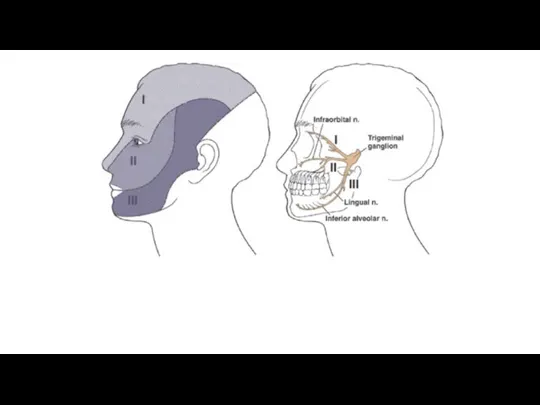
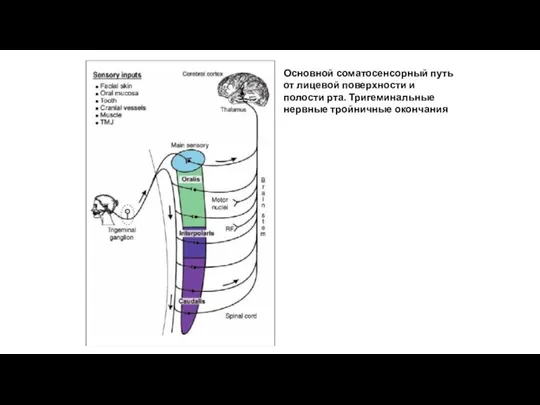
Основной соматосенсорный путь от лицевой поверхности и полости рта. Тригеминальные нервные
Основной соматосенсорный путь от лицевой поверхности и полости рта. Тригеминальные нервные
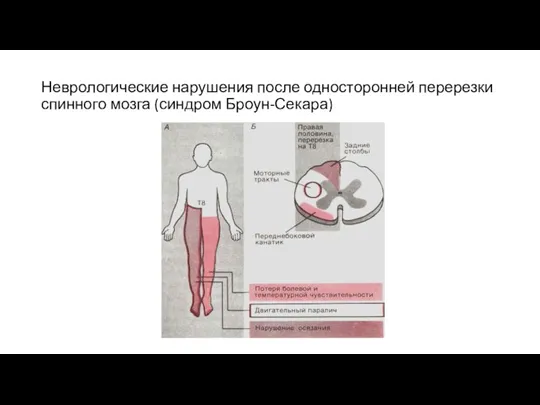
Неврологические нарушения после односторонней перерезки спинного мозга (синдром Броун-Секара)
Неврологические нарушения после односторонней перерезки спинного мозга (синдром Броун-Секара)
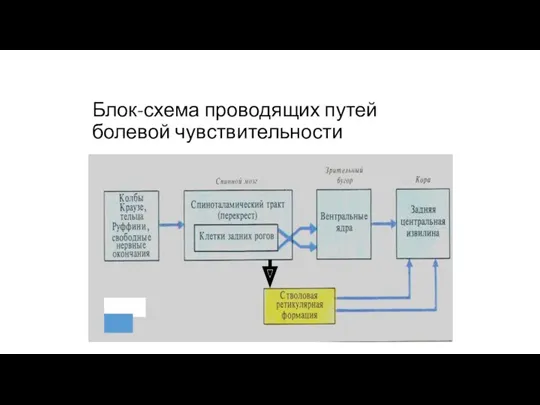
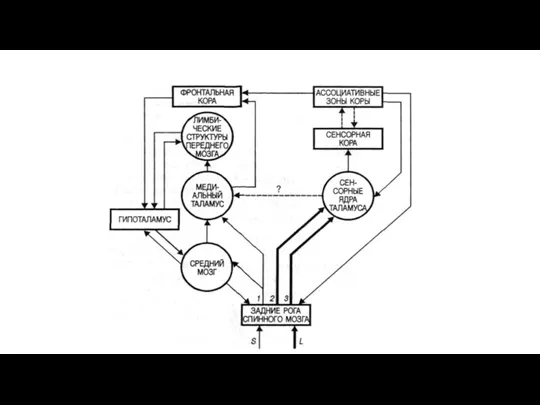
Блок-схема проводящих путей болевой чувствительности
Блок-схема проводящих путей болевой чувствительности
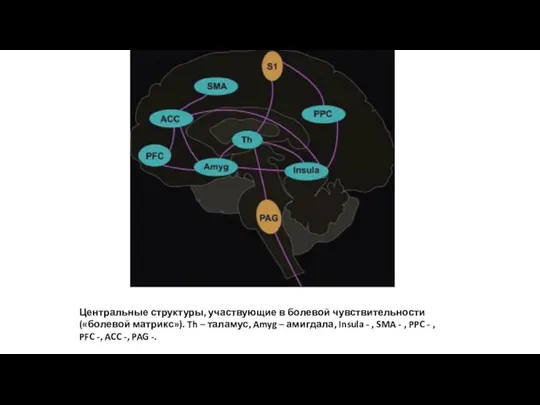
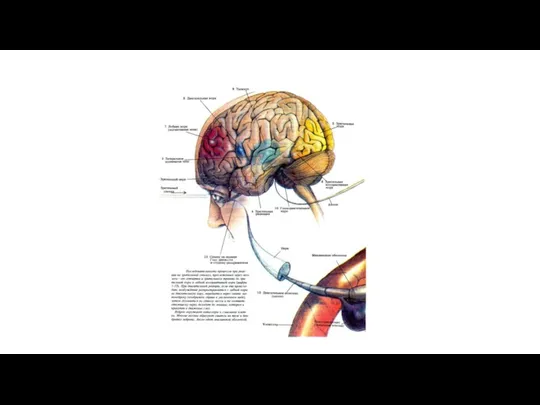
Центральные структуры, участвующие в болевой чувствительности («болевой матрикс»). Th – таламус,
Центральные структуры, участвующие в болевой чувствительности («болевой матрикс»). Th – таламус,

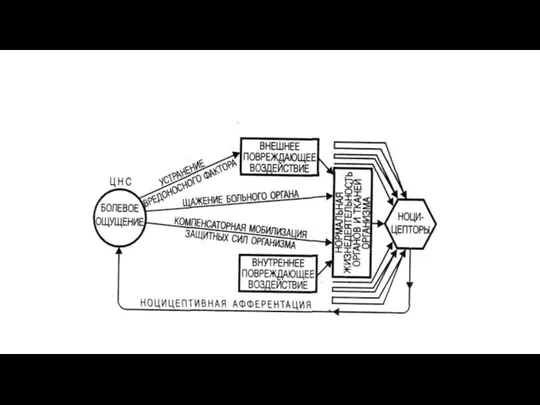
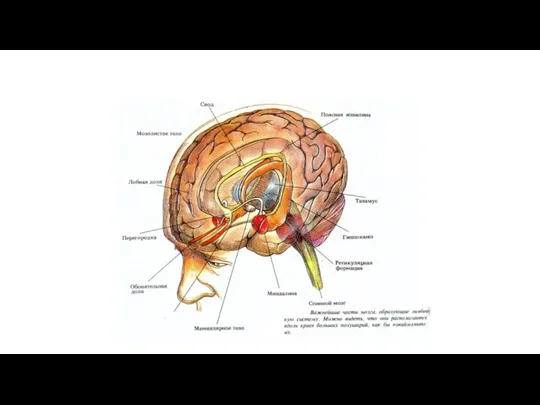
Структуры и механизмы интеграции боли (ноцицептивная система).
Ретикулярная формация - В
Структуры и механизмы интеграции боли (ноцицептивная система).
Ретикулярная формация - В
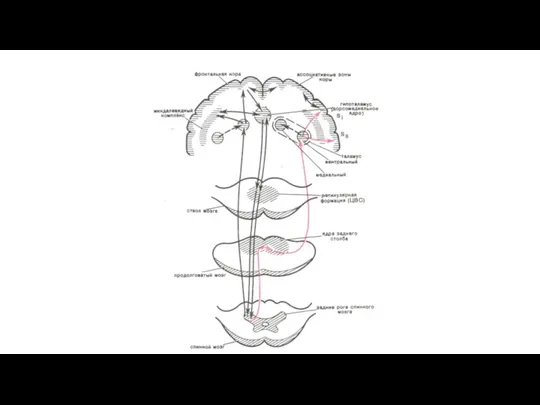

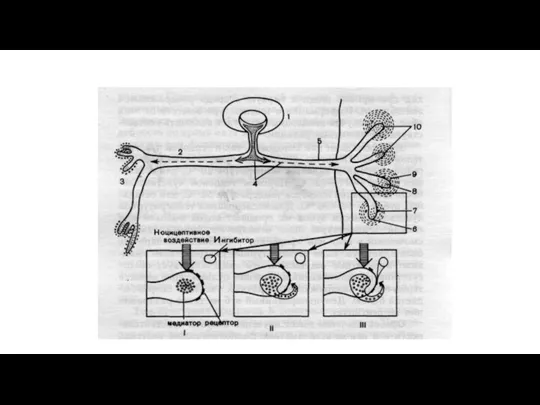
Антиноцицептивная система
Ноцицептивная система имеет свой функциональный антипод - антиноцицептивную систему, которая
Антиноцицептивная система
Ноцицептивная система имеет свой функциональный антипод - антиноцицептивную систему, которая

Структура антиноцицептивнойсистемы
Антиноцицептивная система представлена сегментарным и центральным уровнями контроля, а также
Структура антиноцицептивнойсистемы
Антиноцицептивная система представлена сегментарным и центральным уровнями контроля, а также

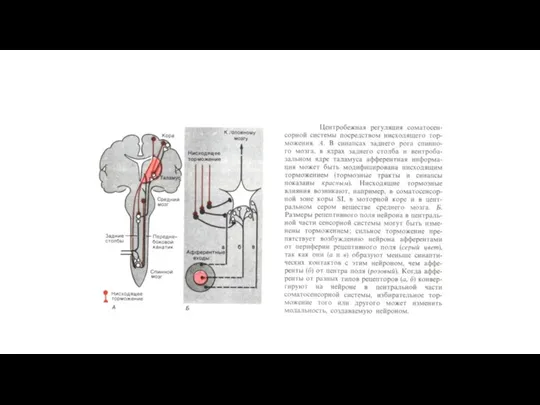
При возникновении боли обязательно происходит активация антиноцицептивной системы.
1. На уровне
При возникновении боли обязательно происходит активация антиноцицептивной системы.
1. На уровне

Опиатные механизмы обезболивания.
Опиатные рецепторы мозга связываются с веществами типа морфина или
Опиатные механизмы обезболивания.
Опиатные рецепторы мозга связываются с веществами типа морфина или

Различные области ЦНС имеют неодинаковую чувствительность эндорфинам и энкефалинам. Например, гипофиз
Различные области ЦНС имеют неодинаковую чувствительность эндорфинам и энкефалинам. Например, гипофиз
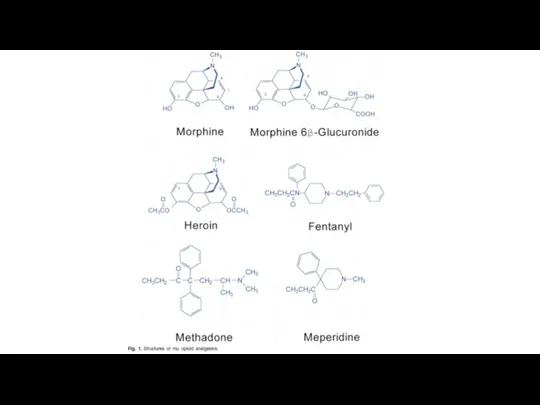
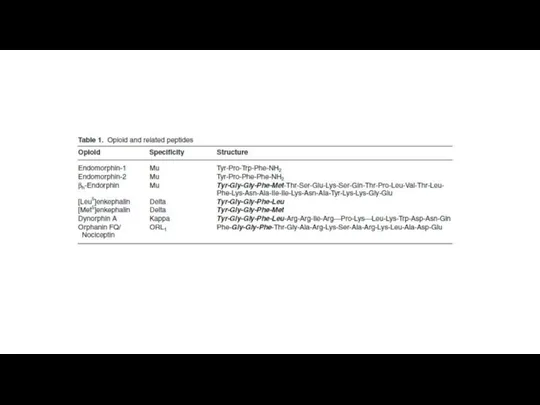

Эндогенные опиаты.
Вещества выделенные из мозга животных, обладают способностью соединяться с опиатными
Эндогенные опиаты.
Вещества выделенные из мозга животных, обладают способностью соединяться с опиатными

Морфин – антиболевые пути
Морфин – антиболевые пути
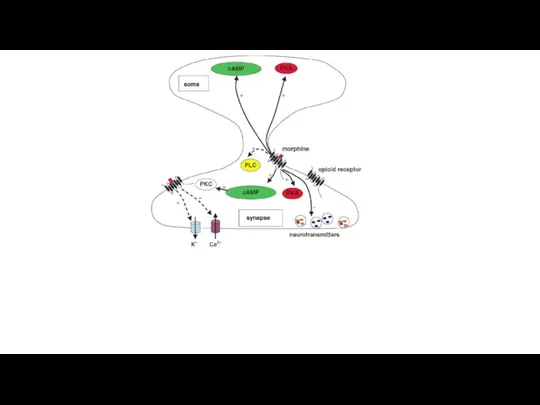

Механизм обезболивающего действия опиатов
Опиаты соединяются с ноцицепторами и препятствуют соединению
Механизм обезболивающего действия опиатов
Опиаты соединяются с ноцицепторами и препятствуют соединению
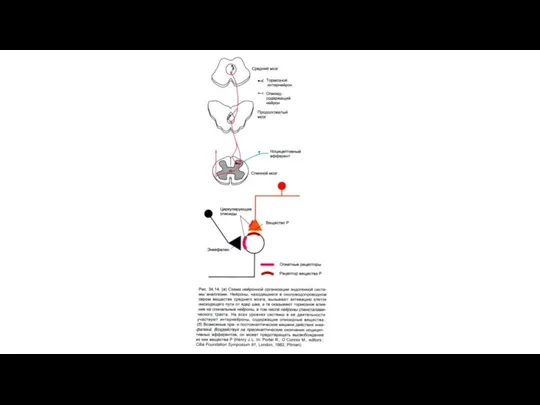

Адренэргические механизмы обезболивания.
Норадреналин, через с α-адренорецепторы тормозит проведение ноцицептивных
Адренэргические механизмы обезболивания.
Норадреналин, через с α-адренорецепторы тормозит проведение ноцицептивных

АЛЬТЕРНАТИВНЫЕ механизмы обезболивания.
Вазопрессин, ангиотензин, окситоцин, соматостатин, нейротензин оказывают аналгетический эффект
АЛЬТЕРНАТИВНЫЕ механизмы обезболивания.
Вазопрессин, ангиотензин, окситоцин, соматостатин, нейротензин оказывают аналгетический эффект

ОСТРАЯ БОЛЬ
"ВОРОТНАЯ" ТЕОРИЯ Р. МЕЛЬЗАКА И
П. УОЛЛА.
Роль ворот,
ОСТРАЯ БОЛЬ
"ВОРОТНАЯ" ТЕОРИЯ Р. МЕЛЬЗАКА И
П. УОЛЛА.
Роль ворот,

ХРОНИЧЕСКАЯ БОЛЬ
При длительном повреждении тканей (воспаление, переломы, опухоли и т.д.) постоянная
ХРОНИЧЕСКАЯ БОЛЬ
При длительном повреждении тканей (воспаление, переломы, опухоли и т.д.) постоянная

Фантомные боли
Фантомные боли (боли в ампутированных конечностях) объясняются в основном
Фантомные боли
Фантомные боли (боли в ампутированных конечностях) объясняются в основном

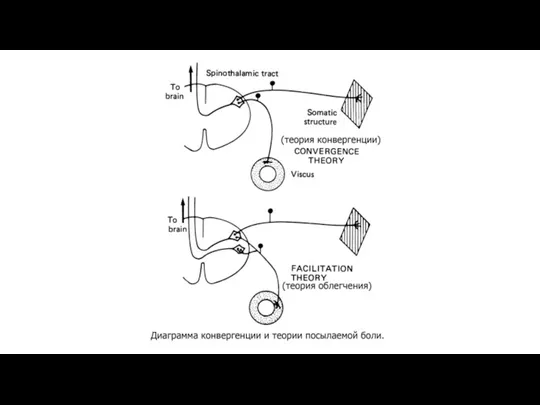
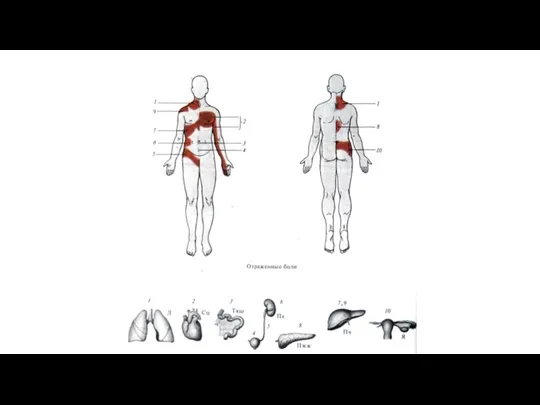
Отраженная боль.
Ее возникновение связано с тем, что афференты внутренних органов и
Отраженная боль.
Ее возникновение связано с тем, что афференты внутренних органов и
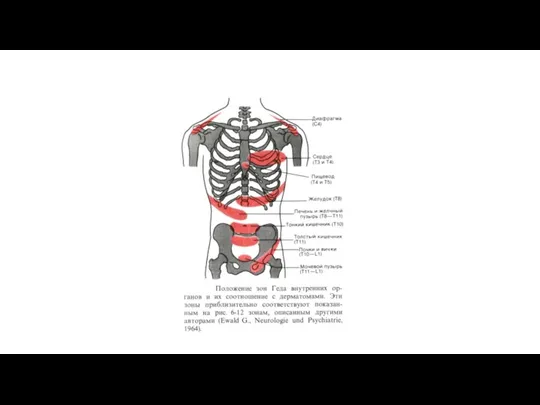
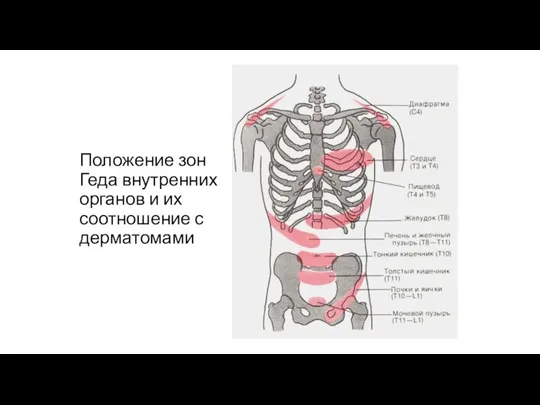
Положение зон Геда внутренних органов и их соотношение с дерматомами
Положение зон Геда внутренних органов и их соотношение с дерматомами
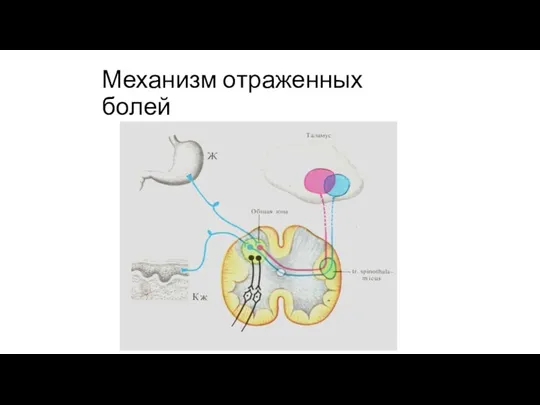
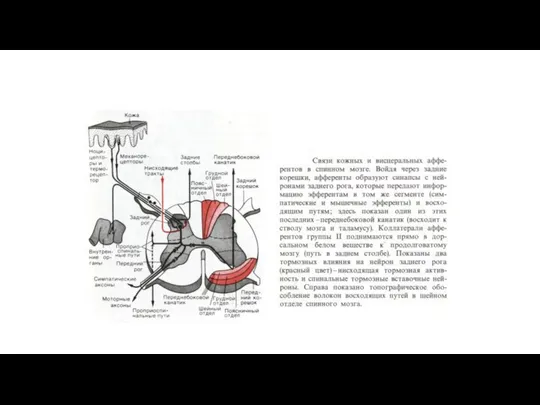
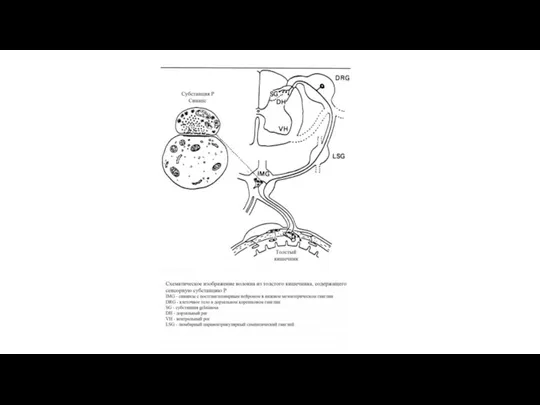
Механизм отраженных болей
Механизм отраженных болей
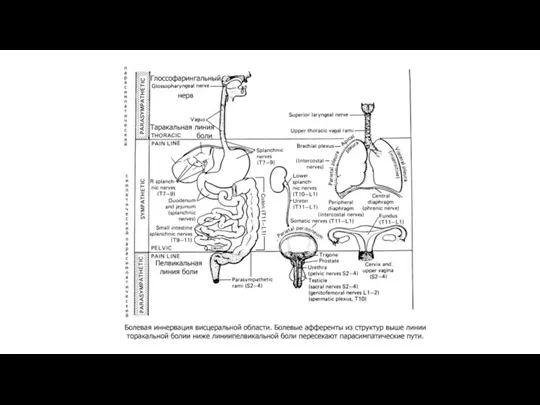

Нарушения функций организма при боли.
Нарушение сна, сосредоточенности, полового влечения,
Нарушения функций организма при боли.
Нарушение сна, сосредоточенности, полового влечения,

Нарушения функций организма при боли.
Биохимические изменения проявляются в виде повышения потребления
Нарушения функций организма при боли.
Биохимические изменения проявляются в виде повышения потребления

Болевая рецепция полости рта.
Выраженной болевой чувствительностью обладает часть слизистой оболочки на
Болевая рецепция полости рта.
Выраженной болевой чувствительностью обладает часть слизистой оболочки на

Виды обезболивания.
Наркоз (ингаляционный и внутривенный
Проводниковая анестезия.
Обезболивание охлаждением.
Электрообезболивание.
Обезболивание иглоукалыванием.
Виды обезболивания.
Наркоз (ингаляционный и внутривенный
Проводниковая анестезия.
Обезболивание охлаждением.
Электрообезболивание.
Обезболивание иглоукалыванием.
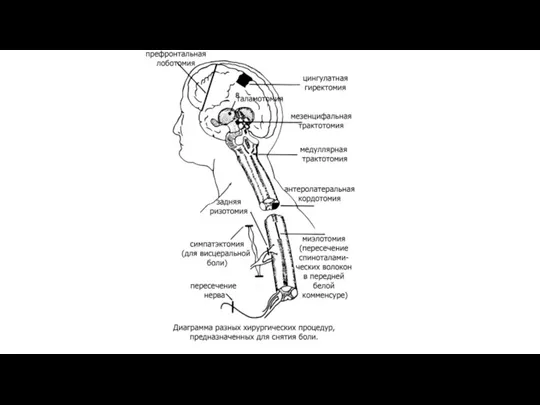
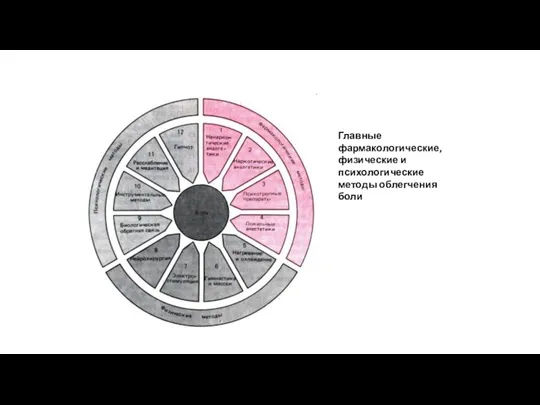
Главные фармакологические, физические и психологические методы облегчения боли
Главные фармакологические, физические и психологические методы облегчения боли
Физиология
высшей
нервной
деятельности
Физиология
высшей
нервной
деятельности

Цель лекции:
Дать характеристику условно-рефлекторной деятельности человека
Мотивация:
Данный раздел физиологии представляет интерес для
Цель лекции:
Дать характеристику условно-рефлекторной деятельности человека
Мотивация:
Данный раздел физиологии представляет интерес для

План лекции:
1. Общая характеристика высшей нервной деятельности животных и человека
2. Методы
План лекции:
1. Общая характеристика высшей нервной деятельности животных и человека
2. Методы

ВЕХИ ИСТОРИИ
1637 — Р. Декарт (R. Descartes, Франция) дал первое
ВЕХИ ИСТОРИИ
1637 — Р. Декарт (R. Descartes, Франция) дал первое





СОЗНАНИЕ
Сознание - свойство мозга - высшее проявление функции отражения действительности.
Сознание -
СОЗНАНИЕ
Сознание - свойство мозга - высшее проявление функции отражения действительности.
Сознание -

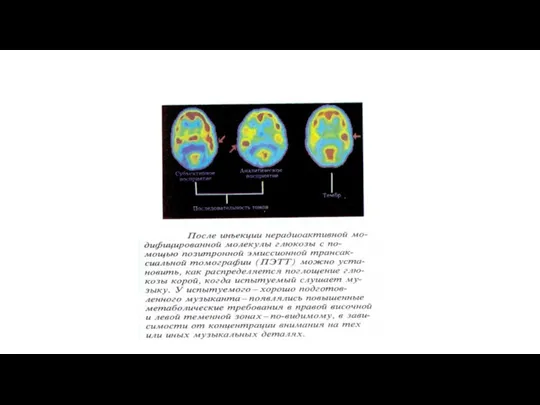
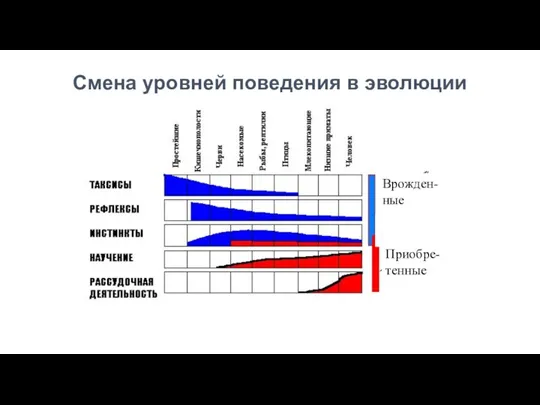
Смена уровней поведения в эволюции
Врожден-ные
Приобре-тенные
Смена уровней поведения в эволюции
Врожден-ные
Приобре-тенные
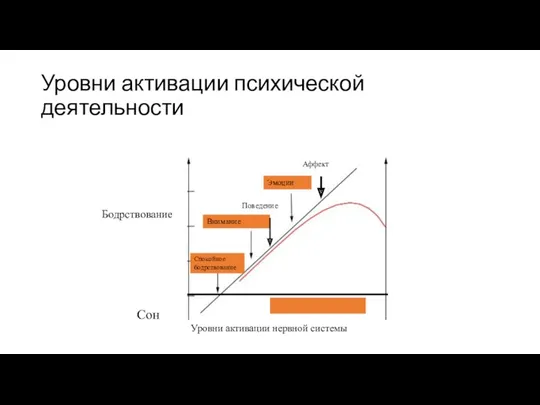
Уровни активации психической деятельности
Эмоции
Внимание
Спокойное бодрствование
Сон
Уровни активации нервной системы
Поведение
Аффект
Бодрствование
Уровни активации психической деятельности
Эмоции
Внимание
Спокойное бодрствование
Сон
Уровни активации нервной системы
Поведение
Аффект
Бодрствование

Таксисы предполагают избирательные реакции приближения к источнику света ( фототаксис), химических
Таксисы предполагают избирательные реакции приближения к источнику света ( фототаксис), химических

Безусловные рефлексы
Врожденные, свойственные всем особям данного вида
Имеют готовую генетически детерминированную
Безусловные рефлексы
Врожденные, свойственные всем особям данного вида
Имеют готовую генетически детерминированную

Витальные инстинкты
Пищевой
Питьевой
Оборонительный (активный -ястреб и пассивный - кролик)
Регулирования цикла «сон-бодрствование»
Экономии энергии
Витальные инстинкты
Пищевой
Питьевой
Оборонительный (активный -ястреб и пассивный - кролик)
Регулирования цикла «сон-бодрствование»
Экономии энергии

Ролевые инстинкты
Половые - выбор партнера
Родительские - разделение ролей отца и матери
Территориальные
Ролевые инстинкты
Половые - выбор партнера
Родительские - разделение ролей отца и матери
Территориальные

Инстинкты саморазвития
Исследовательский
Новизны
Свободы
Имитационный (подражательный)
Игровой
Инстинкты саморазвития
Исследовательский
Новизны
Свободы
Имитационный (подражательный)
Игровой

Отличия условного и безусловного рефлексов:
условные безусловные
приобретенные врожденные
временные постоянные
индивидуальные видовые
вырабатываются на осуществляются только
любой раздражитель при действии адекватного
Отличия условного и безусловного рефлексов:
условные безусловные
приобретенные врожденные
временные постоянные
индивидуальные видовые
вырабатываются на осуществляются только
любой раздражитель при действии адекватного
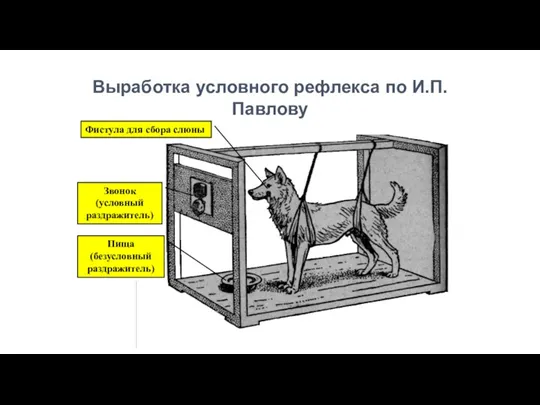
Выработка условного рефлекса по И.П.Павлову
Фистула для сбора слюны
Звонок
(условный раздражитель)
Пища
(безусловный
раздражитель)
Выработка условного рефлекса по И.П.Павлову
Фистула для сбора слюны
Звонок
(условный раздражитель)
Пища
(безусловный
раздражитель)

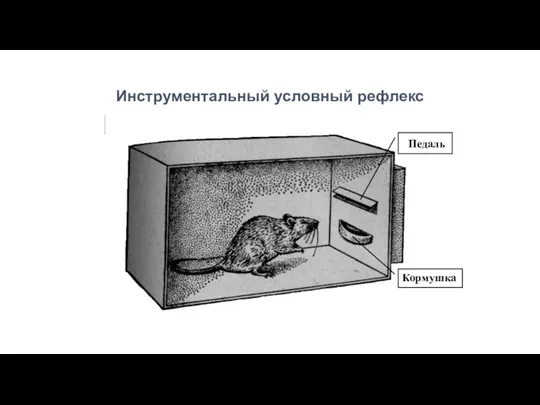
Инструментальный условный рефлекс
Педаль
Кормушка
Инструментальный условный рефлекс
Педаль
Кормушка



Основные характеристики
условного рефлекса (по И.П.Павлову)
1) Приобретаемость условных рефлексов (врожденность безусловных
Основные характеристики
условного рефлекса (по И.П.Павлову)
1) Приобретаемость условных рефлексов (врожденность безусловных


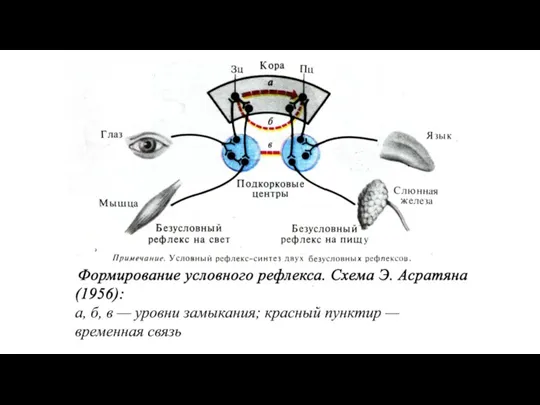
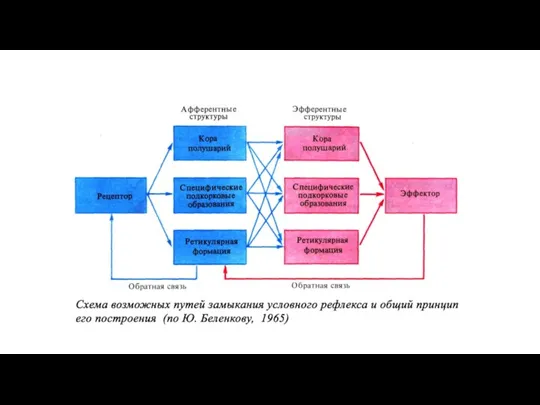
Образование временной связи по пути «кора-кора» по И.П.Павлову
Образование временной связи по пути «кора-кора» по И.П.Павлову
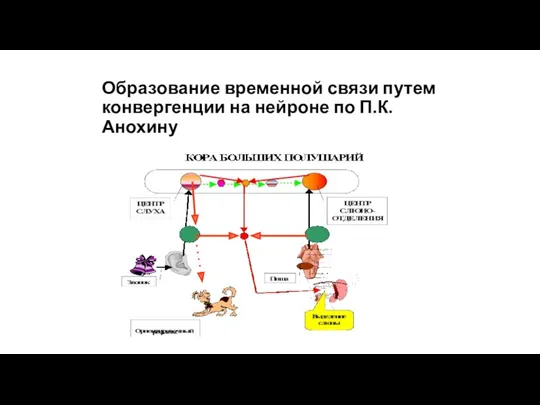
Образование временной связи путем конвергенции на нейроне по П.К.Анохину
Образование временной связи путем конвергенции на нейроне по П.К.Анохину
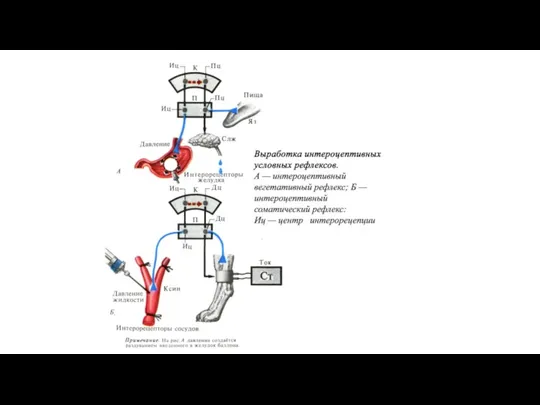

Классификация условных рефлексов
По происхождению - натуральные и искусственные
По характеру безусловного подкрепления
Классификация условных рефлексов
По происхождению - натуральные и искусственные
По характеру безусловного подкрепления

Условия выработки
условных рефлексов
Условие времени - предварительность или одновременность действия условного
Условия выработки
условных рефлексов
Условие времени - предварительность или одновременность действия условного

Механизм замыкания временной связи
Теория двух корковых очагов И.П.Павлова - проторение пути
Теория
Механизм замыкания временной связи
Теория двух корковых очагов И.П.Павлова - проторение пути
Теория
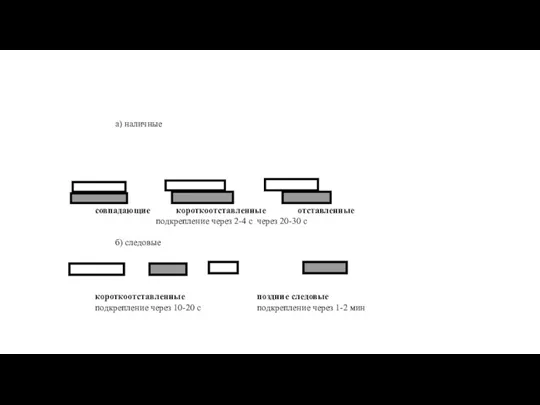
а) наличные
совпадающие короткоотставленные отставленные
подкрепление через 2-4 с через 20-30 с
б) следовые
короткоотставленные поздние следовые
подкрепление через 10-20
а) наличные
совпадающие короткоотставленные отставленные
подкрепление через 2-4 с через 20-30 с
б) следовые
короткоотставленные поздние следовые
подкрепление через 10-20

Правила выработки условного рефлекса:
∙ вырабатывается на базе безусловного
∙ необходимо неоднократное сочетание индифферентного и
Правила выработки условного рефлекса:
∙ вырабатывается на базе безусловного
∙ необходимо неоднократное сочетание индифферентного и

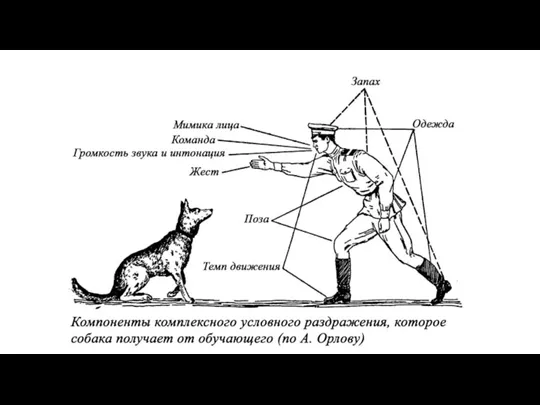

Схемы образования условного рефлекса по И. П. Павлову (А) и Э.
Схемы образования условного рефлекса по И. П. Павлову (А) и Э.
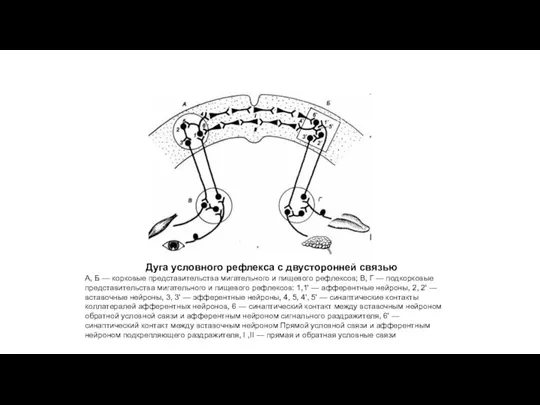
Дуга условного рефлекса с двусторонней связью
А, Б — корковые
Дуга условного рефлекса с двусторонней связью
А, Б — корковые

Взаимодействие синаптических входов с гетерохимической организацией для стимулов биологической (а) и
Взаимодействие синаптических входов с гетерохимической организацией для стимулов биологической (а) и
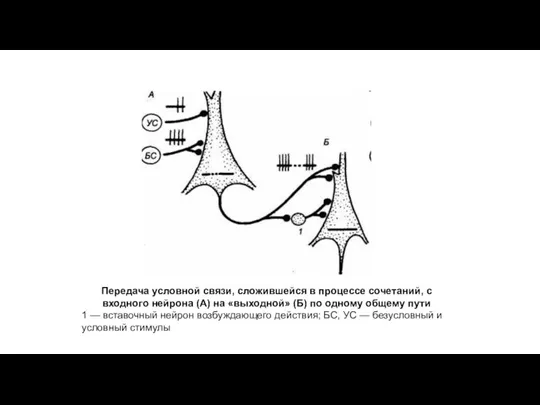
Передача условной связи, сложившейся в процессе сочетаний, с входного нейрона (А)
Передача условной связи, сложившейся в процессе сочетаний, с входного нейрона (А)
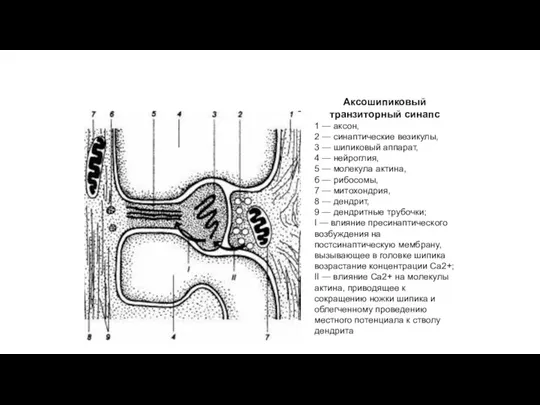
Аксошипиковый транзиторный синапс
1 — аксон,
2 — синаптические везикулы,
3 —
Аксошипиковый транзиторный синапс
1 — аксон,
2 — синаптические везикулы,
3 —
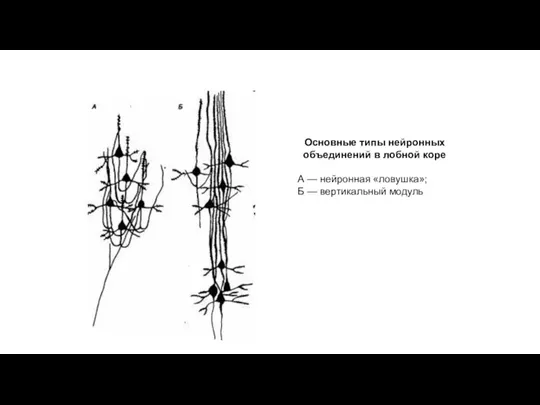
Основные типы нейронных объединений в лобной коре
А — нейронная «ловушка»;
Б
Основные типы нейронных объединений в лобной коре
А — нейронная «ловушка»;
Б

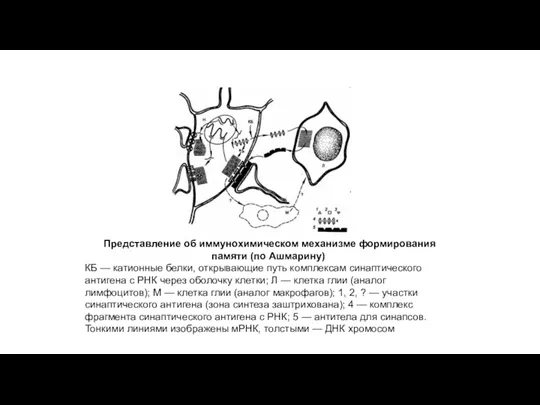
Представление об иммунохимическом механизме формирования памяти (по Ашмарину)
КБ — катионные
Представление об иммунохимическом механизме формирования памяти (по Ашмарину)
КБ — катионные

Безусловное торможение:
внешнее торможение (протекает по типы постоянного и гаснущего тормоза),
запредельное торможение
Безусловное торможение:
внешнее торможение (протекает по типы постоянного и гаснущего тормоза),
запредельное торможение
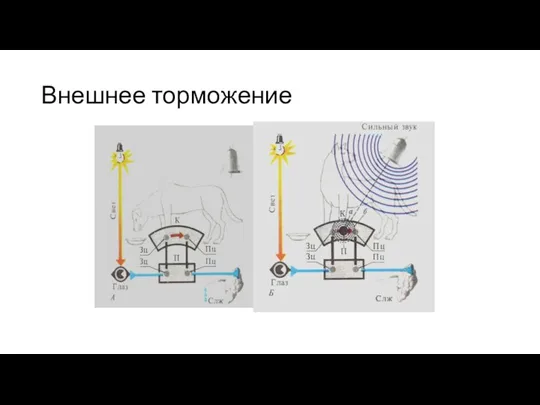
Внешнее торможение
Внешнее торможение

Угасательное торможение
звонок
пища
звонок
звонок
Угасательное торможение
звонок
пища
звонок
звонок
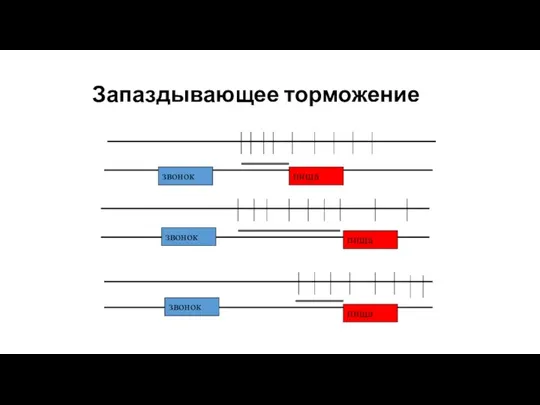
Запаздывающее торможение
звонок
пища
пища
пища
звонок
звонок
Запаздывающее торможение
звонок
пища
пища
пища
звонок
звонок

Дифференцировочное торможение
Дифференцировочное торможение
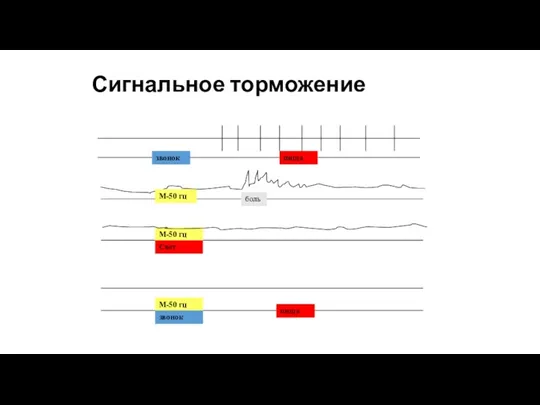
Сигнальное торможение
Сигнальное торможение

Образование угасательного торможения
А — условный пищевой рефлекс;
Б — стадия
Образование угасательного торможения
А — условный пищевой рефлекс;
Б — стадия

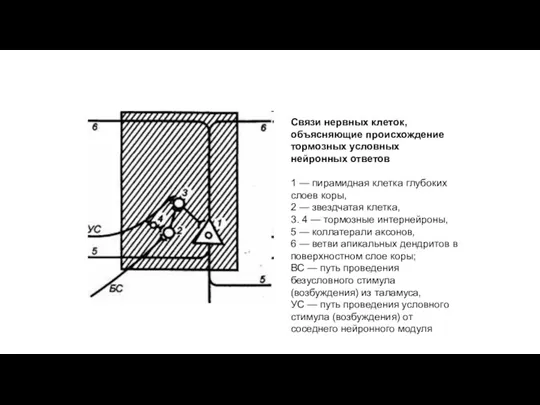
Связи нервных клеток, объясняющие происхождение тормозных условных нейронных ответов
1 — пирамидная
Связи нервных клеток, объясняющие происхождение тормозных условных нейронных ответов
1 — пирамидная


 Нарушения речи и особенности их коррекции у детей с интеллектуальной недостаточностью
Нарушения речи и особенности их коррекции у детей с интеллектуальной недостаточностью Лікарські засоби, що впливають на серцево-судинну систему та функцію нирок
Лікарські засоби, що впливають на серцево-судинну систему та функцію нирок Геморрагические лихорадки
Геморрагические лихорадки Гемолитическая болезнь плода и новорожденного. Повышение непрямого билирубина
Гемолитическая болезнь плода и новорожденного. Повышение непрямого билирубина Особенности фармакокинетики лекарственных веществ, вводимых роженице в родах и их влияние на плод
Особенности фармакокинетики лекарственных веществ, вводимых роженице в родах и их влияние на плод Инвазия. Формы инвазии
Инвазия. Формы инвазии Расспрос больных с заболеваниями органов дыхания. Общий осмотр. Осмотр и пальпация грудной клетки. Перкуссия легких
Расспрос больных с заболеваниями органов дыхания. Общий осмотр. Осмотр и пальпация грудной клетки. Перкуссия легких Әлеуметтік диагностика
Әлеуметтік диагностика Энергетическая ценность пищевых продуктов
Энергетическая ценность пищевых продуктов Функционирование учреждения здравоохранения в условиях чрезвычайных ситуаций
Функционирование учреждения здравоохранения в условиях чрезвычайных ситуаций Клиническая психология
Клиническая психология Предупреждение желудочно-кишечных болезней
Предупреждение желудочно-кишечных болезней Пролиферативное воспаление
Пролиферативное воспаление Переломи і опіки
Переломи і опіки ОП нарушения кровообращения
ОП нарушения кровообращения 24-hour patterns of central and peripheral systolic blood pressure in the very elderly with HFpEF
24-hour patterns of central and peripheral systolic blood pressure in the very elderly with HFpEF Периферическая нервная система. Спинномозговые нервы
Периферическая нервная система. Спинномозговые нервы Медицинская служба Вооруженных сил РФ в ЧС
Медицинская служба Вооруженных сил РФ в ЧС Рожистое воспаление
Рожистое воспаление Гипогликемия и гипергликемия у новорожденных
Гипогликемия и гипергликемия у новорожденных Первая помощь при термотравмах и электротравмах
Первая помощь при термотравмах и электротравмах Переливание крови. История. Изогемагглютинация, группы крови. Механизм действия перелитой крови
Переливание крови. История. Изогемагглютинация, группы крови. Механизм действия перелитой крови Проблема инвалидности
Проблема инвалидности Эффект применения Флуфеназина в лечении синдрома Туретта
Эффект применения Флуфеназина в лечении синдрома Туретта Синдром пангипопитуитаризма
Синдром пангипопитуитаризма Вторичный метаболизм
Вторичный метаболизм Клинико-психологическое сопровождение пожилых людей страдающих инволлюционным паранойдом
Клинико-психологическое сопровождение пожилых людей страдающих инволлюционным паранойдом Ультразвуковые маркеры хромосомной патологии
Ультразвуковые маркеры хромосомной патологии