- Физиология выделительной системы

Содержание
- 2. Функции почек: Выделительная функция (избыток воды, неорганических и органических веществ, продукты азотистого обмена и чужеродные вещества).
- 3. Регуляция кислотно-основного состояния (путем экскреции Н+, нелетучих кислот и оснований). Инкреторная функция (ренин, активная форма витамина
- 4. Регуляция уровня АД (секреция ренина, экскреция Na+ и воды, изменение ОЦК). Регуляция эритропоэза (секреция эритропоэтина). Регуляция
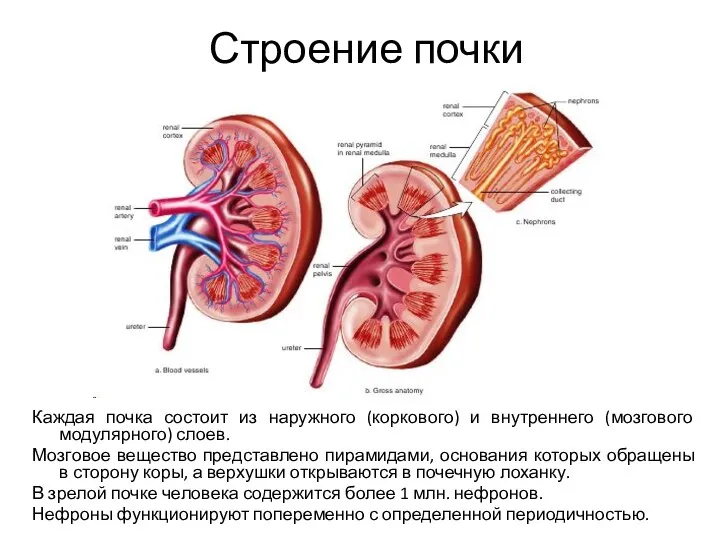
- 5. Строение почки Каждая почка состоит из наружного (коркового) и внутреннего (мозгового модулярного) слоев. Мозговое вещество представлено
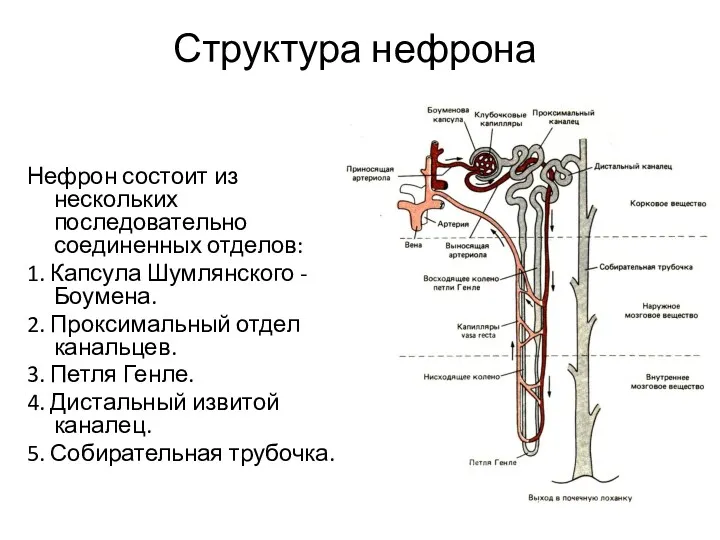
- 6. Структура нефрона Нефрон состоит из нескольких последовательно соединенных отделов: 1. Капсула Шумлянского - Боумена. 2. Проксимальный
- 7. Различают 3 типа нефронов: суперфициальные (поверхностные) (20-30%), участвуют в фильтрации мочи; интракортикальные (корковые) (60-70%), выполняя главную
- 8. Кровоснабжение почек мл/мин на 1 г ткани Через сосуды почки в 1 мин проходит около 1/4
- 9. За минуту проходит ≈ 1200 мл крови или ≈ 400мл/100г в минуту, что превышает кровоток в
- 10. Почечный кровоток делится на 2 типа: корковый – примерно 80-90 %; мозговой – ≈ 10-20 %,
- 11. Механизмы поддержания почечного кровотока: Изменение тонуса сосудов почки при изменении давления в почечной артерии. Изменение соотношения
- 12. Оценка коркового кровотока Проводится путём определения коэффициента очищения, или клиренса, таких веществ как парааминогиппуровая кислота (ПАГ),
- 13. Физиология мочеобразования
- 14. Механизм образования мочи 1. Фильтрация 2. Реабсорбция 3. Секреция
- 15. Процессы, участвующие в образовании мочи: В результате фильтрации образуется первичная моча В результате реабсорбции и секреции
- 16. Строение клубочкового фильтра Гломерулярный фильтр имеет 3 слоя: 1. Эндотелий капилляров имеет поры диаметром 50-100 нм,
- 17. Базальная мембрана – трехслойная структура, толщиной до 300 нМ и является основным барьером для процессов фильтрации.
- 18. Подоциты – эпителиальные клетки висцерального листка капсулы 3. Отростки подоцитов имеют щелевые диафрагмы пронизанные порами, диаметр
- 19. Состав ультрафильтрата (первичная моча) Близок по концентрации веществ в плазме крови (кроме белка). Вещества с молекулярной
- 20. В норме, за сутки, фильтруется до 5 г белка – в основном альбумины, реабсорбируется более 99
- 21. Зависимость между размерами молекул веществ и и прохождением их через фильтрационный барьер
- 22. Фильтрационное давление 1. Гидростатическое давление в клубочке (ГДкл) = 70 мм рт.ст. 2. Онкотическое давление в
- 23. Скорость клубочковой фильтрации Определяется по величине клиренса. C = M×V/П, где С - клиренс, М –
- 24. Клиренс – это скорость, с которой объём плазмы полностью очищается от конкретного вещества в единицу времени.
- 25. В клинической практике измерение СКФ проводится с помощью эндогенного индикатора, который находится в плазме крови –
- 26. Обычно СКФ составляет 85-135 мл/мин на каждые 1,73 м2 поверхности тела. Клиренс (С) может быть определён
- 27. ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ОБЪЁМ ФИЛЬТРАТА Фильтрационное давление – движущая сила перехода жидкости из капилляров почечного клубочка в
- 28. Регуляция скорости клубочковой фильтрации (СКФ) осуществляется с помощью механизмов регуляции коркового кровотока (через вазомоторные влияния –
- 29. Регуляция скорости клубочковой фильтрации (СКФ) Симпатические влияния – снижают величину клубочкового кровотока и как следствие, объём
- 30. Мочевина – образуется в результате катаболизма белка. За сутки, из 100 гр. белка освобождается ≈ 16
- 31. Канальцевая реабсорбция это процесс обратного всасывания воды и ряда растворенных в ней веществ из просвета канальцев
- 32. Вещества, содержащиеся в моче, по степени их реабсорбции, делятся на: Беспороговые (непороговые) – выделяются с мочой
- 33. Избирательность реабсорбции Физиологически ценные, жизненно важные вещества реабсорбируются полностью: витамины, аминокислоты, низкомолекулярные белки. Реабсорбируется большая часть
- 34. Реабсорбция подразделяется на: Облигатную (обязательную), нерегулируемую – реабсорбируется фактически та же плазма, которая профильтровалась в капсулу
- 35. Относительный объём проксимальной (облигатной) реабсорбции – около 2/3 (65 %) от скорости клубочковой фильтрации «правило двух
- 36. Облигатная реабсорбция веществ из проксимальных канальцев происходит за счёт первичной реабсорбции Na+, за счёт активного транспорта
- 37. Факультативная реабсорбция – происходит в основном в дистальных извитых канальцах и собирательных трубочках реабсорбируется 9% общего
- 38. Основная масса молекул реабсорбируется в проксимальном извитом отделе нефрона. В петле Генле, дистальном канальце и собирательных
- 39. Механизмы канальцевой реабсорбции Канальцевая реабсорбция может быть активной или пассивной.
- 40. Пассивный транспорт осуществляется по принципу как облегчённой диффузии (реабсорбция Н2О, СО2, хлориды). Так и по электрохимическому
- 41. Механизм канальцевой реабсорбции Обеспечивается: активным и пассивным транспортом. Виды активного транспорта: 1. Первично-активный – против электрохимического
- 42. Реабсорбция глюкозы Глюкоза полностью реабсорбируется в проксимальном канальце путем вторично-активного транспорта с Na+. Гликозурия происходит когда
- 43. Реабсорбция аминокислот (АК) АК полностью реабсорбируются в проксимальном канальце путем вторично-активного транспорта с Na+. Почки не
- 44. В проксимальном отделе нефрона на апикальной мембране эпителия (подобно эпителию тонкого кишечника ЖКТ) имеются белки переносчики,
- 45. Внутри клеток эпителия концентрация АК увеличивается в несколько раз по сравнению с плазмой крови: так туарина
- 46. Реабсорбция аминокислот Аминокислоты (АК) – реабсорбируются более чем на 99% в проксимальном отделе нефрона: валин >
- 47. Реабсорбция белков Белки реабсорбируются полностью в проксимальном канальце путем пиноцитоза. В дальнейшем они распадаются в клетках
- 48. Механизм эндоцитоза белков За счёт наличия на щёточной кайме рецепторного мегалин-кубилинового комплекса связывающего белки и транспортирующего
- 49. Реабсорбция белков Большие пептиды (инсулин, альбумины и другие) поступают в клетки эпителия проксимальных канальцев за счёт
- 50. Белок в моче В ультрафильтрате обнаруживается ряд белков такие как альбумины (до 4г/сут), лизоцим, обломки иммуноглобулинов,
- 51. Реабсорбция мочевины Мочевина – это неполярное низкомолекулярное соединение, поэтому легко проникает через клеточные мембраны. Реабсорбция мочевины
- 52. Реабсорбция слабых кислот и оснований Реабсорция зависит от pH внутриканальцевой жидкости. Слабые основания и кислоты реабсорбируются
- 53. Реабсорбция воды по всему протяжению почечных канальцев происходит только пассивно. Из 170 л отфильтрованной воды в
- 54. Аквапорины (AQP) — семейство мембранных белков-пор для воды. Идентифицировано 10 аквапоринов. AQP3, AQP7 и AQP9 дополнительно
- 55. Аквапорины в почке. Аквапорин 1 (AQP1) экспрессируется в апикальной и базолатеральной мембране клеток проксимального канальца нефрона
- 56. Реабсорбция воды Реабсорбция воды в проксимальных отделах нефрона постоянна и происходит вслед за активным транспортом Na+.
- 60. Реабсорбция электролитов 1. Реабсорбция Na+ идет в 2 этапа: а. выкачивание Na+ через базальную мембрану клеток
- 61. ПРОТИВОТОЧНО-МНОЖИТЕЛЬНЫЙ МЕХАНИЗМ КОНЦЕНТРИРОВАНИЯ МОЧИ
- 62. Эта система объединяет в себе 3 взаимодополняющих звена, каждое из которых состоит из двух параллельных трубок
- 63. Первое звено: Определяющую роль в работе противоточного механизма играет восходящее колено петли Генле. Из него реабсорбируется
- 64. Нисходящее колено проницаемо для воды, которая уходит в гиперосмотический интерстиций. Чем больше Na+ реабсорбируется в восходящем
- 65. Второе звено работает по тому же принципу. Но стенка собирательной трубочки становится проницаемой для воды только
- 66. Третье звено: В нисходящем прямом капилляре кровь отдает воду из-за нарастающего осмотического давления интерстиции, сгущается и
- 67. Канальцевая секреция Канальцевая секреция - это транспорт веществ из крови в просвет канальцев. Канальцевая секреция представляет
- 68. Регуляция водно-солевого баланса Повышение осмотического давления крови возбуждение осморецепторов (в области супраоптического ядра гипоталамуса, а также
- 69. Инсулин. Недостаток этого гормона приводит к гипергликемии, глюкозурии, увеличению осмотического давления мочи и увеличению диуреза. Тироксин
- 70. Регуляция ионного состава крови
- 71. Na+ Альдостерон усиливает реабсорбцию Na+ в дистальных отделах канальцев и в собирательных трубочках. Секреция альдостерона увеличивается
- 72. .Различные лекарственные средства (диуретики), подавляя реабсорбцию Na+, увеличивают экскрецию Na+ и воды, тем самым уменьшая в
- 73. K+ Альдостерон усиливает секрецию K+ в дистальном отделе канальцев и собирательных трубочках. Инсулин уменьшает выделение K+,
- 74. Реабсорбция кальция Концентрация кальция в плазме крови ≈ 2,5 мМ, значительная часть его связана с белками.
- 75. Реабсорбция кальция Реабсорбция ионов Са2+ в канальце осуществляется за счёт: - Пассивного межклеточного механизма, - Трансклеточного
- 76. Ca2+ Паратгормон увеличивает реабсорбцию Ca2+ и высвобождение Ca 2+ из костей, что приводит к повышению его
- 77. Cl - Альдостерон участвует в регуляции уровня Cl- в плазме крови. При увеличении реабсорбции Na+ возрастает
- 78. Реабсорбция Pi продолжается и дистальнее проксимального канальца, так что в итоге фракционное выведение составляет 10-20%. По
- 79. Реабсорбция фосфата в проксимальном канальце Неорганический фосфат (Pi) – содержится в плазме в виде HPO42- и
- 80. Паратиреоидный гормон (ПТГ) (паращитовидные железы), действие которого опосредуется через цАМФ или IP3/DAГ влияет на количество NaPi-3-обменника
- 81. Поддержание кислотно-щелочного равновесия
- 82. Основания с большей скоростью экскретируются с кислой мочой, а кислоты - с щелочной мочой.
- 83. 2. За счет работы карбоангидразной системы Секреция иона Н+ в обмен на ион Nа+ приводит к
- 84. 3. Выведение Н+ с фосфатами. Секретируемые Н+ в просвете канальца связываются также с Na2HPO4, вытесняя из
- 85. 4. В результате дезаминирования АК в почках образуется аммиак и выход его в просвет канальца. Н+
- 86. Нервная регуляция мочеобразования. Морфологические основы. Симпатическая эфферентная иннервация почек осуществляется из Th5-L3 сегментов спинного мозга (по
- 87. Физиологические основы регуляции фильтрации Возбуждение симпатической нервной системы уменьшает фильтрацию, за счёт спазма сосудов почечных клубочков
- 88. Регуляция величины АД
- 89. В регуляции АД участвует юкстагломерулярный аппарат (ЮГА) почки, который отвечает за выработку ренина
- 90. Часть начального отдела дистального извитого канальца очень плотно прилегает к корню клубочка и имеет особое строение
- 91. Клетки ЮГА иннервируются симпатическими волокнами и содержат β- и α-ADR. Активация β-ADR усиливает секрецию ренина, а
- 92. Юкстагломерулярный комплекс работает по принципу обратной отрицательной связи: 1. при снижении скорости клубочковой фильтрации уменьшается количество
- 93. Регуляция выработки ренина При понижении давления в приносящей артериоле в гранулярных клетках ЮГА вырабатывается ренин. Продукция
- 94. Возбуждение β-адренорецепторов гранулярных клеток приводит к усилению секреции ренина, а α-адренорецепторов - торможению. Простагландины, арахидоновая кислота
- 95. Реакция почек на снижение АД
- 96. Реакция почек на увеличение АД (внутрисистемная и внесистемная)
- 97. ↑ АД в почке ↑ клубочково-капиллярного гидростатического давления ↑ скорость клубочковой фильтрации ↑ скорость движения жидкости
- 98. Реакция почек на увеличение АД
- 99. Нервная регуляция мочеобразования
- 100. Влияние СНС: При сужении приносящих артериол → ↓ фильтрация. Сужение выносящих артериол →↑ фильтрации. Стимуляция симпатических
- 101. При болевых раздражениях диурез рефлекторно уменьшается вплоть до анурии. Механизм: - сужение почечных сосудов при стимуляции
- 102. Раздражение ПСНС вызывает усиление реабсорбции глюкозы и секреции органических кислот. Условно-рефлекторно может происходить уменьшение или увеличение
- 103. Непочечные органы выделения:
- 104. Дыхательная система выводит: углекислый газ и воду, пары эфира и хлороформа, сивушные масла, алкоголь, ацетон при
- 105. Печень и ЖКТ выводят: воду, продукты обмена гемоглобина и холестерина, лекарственные препараты (антибиотики), соли тяжелых металлов,
- 106. Потовые железы удаляют: воду, мочевину, мочевую кислоту, креатинин, молочную кислоту, соли щелочных металлов, летучие жирные кислоты,
- 107. свободные жирные и неомыляемые кислоты, продукты обмена половых гормонов. Сальные железы удаляют:
- 108. Инструментальные методы исследования. Обзорный рентгеновский снимок определит положение, величину и форму почек, а также рентгеноконтрастные камни.
- 109. Транслюмбальная ангиография (норма) осуществляют путем введения контрастного вещества в аорту посредством ее пункции в поясничной области
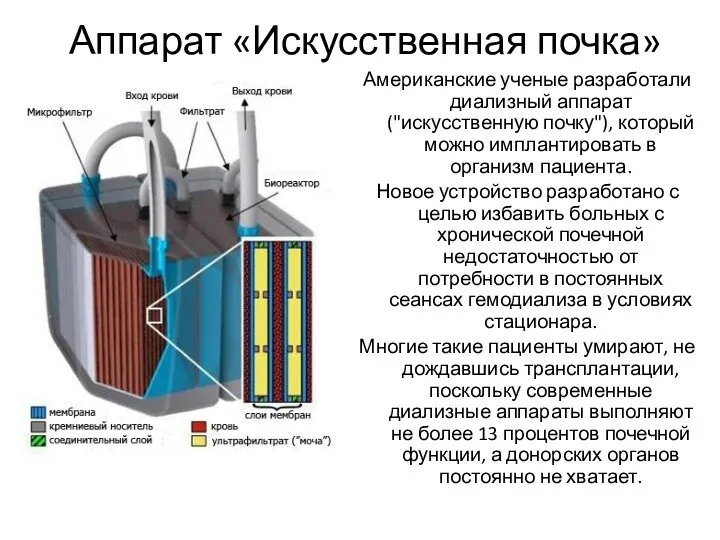
- 110. Аппарат «Искусственная почка»
- 111. Аппарат «Искусственная почка» Американские ученые разработали диализный аппарат ("искусственную почку"), который можно имплантировать в организм пациента.
- 112. ФИЗИОЛОГИЯ МОЧЕВЫДЕЛЕНИЯ
- 113. Морфологические основы В процессе мочеобразования моча, из собирательных трубочек по выводным протокам поступает в чашечки, а
- 114. Мускулатура мочевого пузыря – трёхслойная: наружный и внутренний слои – продольные, средний слой – циркулярный (самый
- 115. Позыв к мочеиспусканию возникает при 150-200 мл. Наполнение до 300 мл вызывает резкое усиление афферентной импульсации
- 116. Эфферентные волокна – идут от пирамидных нейронов КБП к различным структурам головного мозга и к центрам
- 117. Иннервация мочевого пузыря
- 118. Парасимпатическая система оказывает стимулирующее влияние на сокращение гладких мышц мочевого пузыря. Медиатр АЦХ – рецепторы М-холинорецепторы.
- 119. Повышение активности симпатической системы – создаёт условия для наполнения мочевого пузыря. Повышение активности парасимпатической системы –
- 120. Мочевыделение Физиологические основы.
- 121. Центры контроля мочеиспускания находятся: – коркового контроля – в первой сенсорной зоне на медиальной поверхности полушарий.
- 122. Эфференты парасимпатической системы вызывают сокращение гладких мышц мочевого пузыря и его опорожнение. В начале мочеиспускания давление
- 123. Таким образом: корковые нейроны и средний мозг оказывают тормозящее влияние на центры мочеиспускания спинного мозга. Задний
- 125. Скачать презентацию

Функции почек:
Выделительная функция (избыток воды, неорганических и органических веществ, продукты азотистого
Функции почек:
Выделительная функция (избыток воды, неорганических и органических веществ, продукты азотистого

Регуляция кислотно-основного состояния (путем экскреции Н+, нелетучих кислот и оснований).
Инкреторная
Регуляция кислотно-основного состояния (путем экскреции Н+, нелетучих кислот и оснований).
Инкреторная

Регуляция уровня АД (секреция ренина, экскреция Na+ и воды, изменение ОЦК).
Регуляция уровня АД (секреция ренина, экскреция Na+ и воды, изменение ОЦК).
Строение почки
Каждая почка состоит из наружного (коркового) и внутреннего (мозгового модулярного)
Строение почки
Каждая почка состоит из наружного (коркового) и внутреннего (мозгового модулярного)
Структура нефрона
Нефрон состоит из нескольких последовательно соединенных отделов:
1. Капсула Шумлянского -
Структура нефрона
Нефрон состоит из нескольких последовательно соединенных отделов:
1. Капсула Шумлянского -
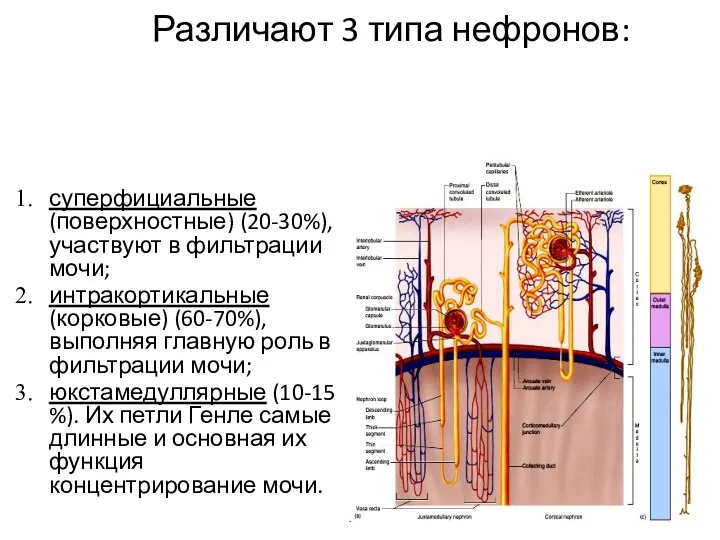
Различают 3 типа нефронов:
суперфициальные (поверхностные) (20-30%), участвуют в фильтрации мочи;
интракортикальные (корковые)
Различают 3 типа нефронов:
суперфициальные (поверхностные) (20-30%), участвуют в фильтрации мочи;
интракортикальные (корковые)
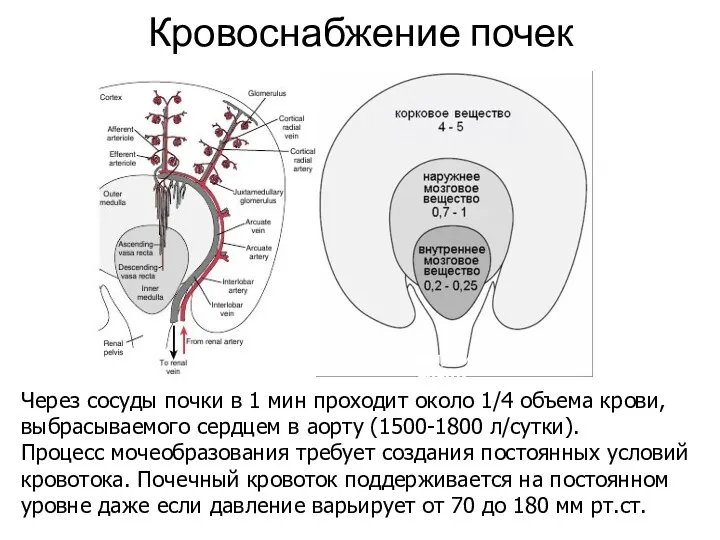
Кровоснабжение почек
мл/мин на 1 г ткани
Через сосуды почки в
Кровоснабжение почек
мл/мин на 1 г ткани
Через сосуды почки в

За минуту проходит ≈ 1200 мл крови или ≈ 400мл/100г в
За минуту проходит ≈ 1200 мл крови или ≈ 400мл/100г в

Почечный кровоток делится на 2 типа: корковый – примерно 80-90 %;
Почечный кровоток делится на 2 типа: корковый – примерно 80-90 %;
Механизмы поддержания почечного кровотока:
Изменение тонуса сосудов почки при изменении давления в
Механизмы поддержания почечного кровотока:
Изменение тонуса сосудов почки при изменении давления в

Оценка коркового кровотока
Проводится путём определения коэффициента очищения, или клиренса, таких
Оценка коркового кровотока Проводится путём определения коэффициента очищения, или клиренса, таких
Физиология мочеобразования
Физиология мочеобразования

Механизм образования мочи
1. Фильтрация
2. Реабсорбция
3. Секреция
Механизм образования мочи
1. Фильтрация
2. Реабсорбция
3. Секреция
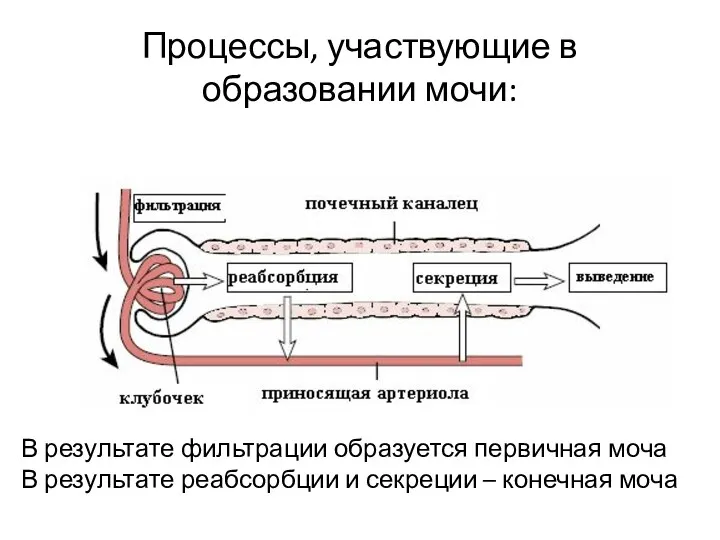
Процессы, участвующие в образовании мочи:
В результате фильтрации образуется первичная моча
В
Процессы, участвующие в образовании мочи:
В результате фильтрации образуется первичная моча
В
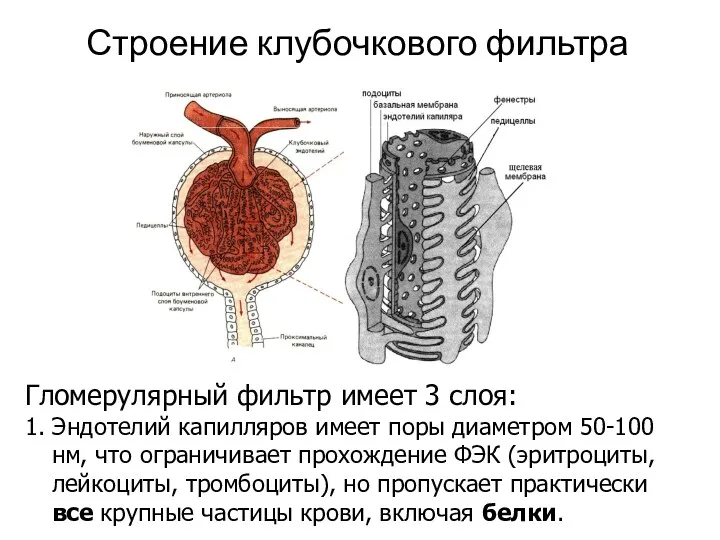
Строение клубочкового фильтра
Гломерулярный фильтр имеет 3 слоя:
1. Эндотелий капилляров имеет
Строение клубочкового фильтра
Гломерулярный фильтр имеет 3 слоя:
1. Эндотелий капилляров имеет
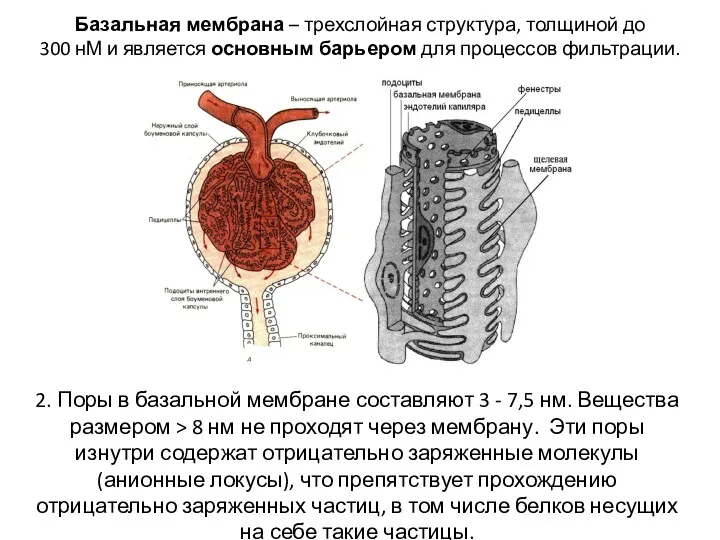
Базальная мембрана – трехслойная структура, толщиной до
300 нМ и является
Базальная мембрана – трехслойная структура, толщиной до 300 нМ и является
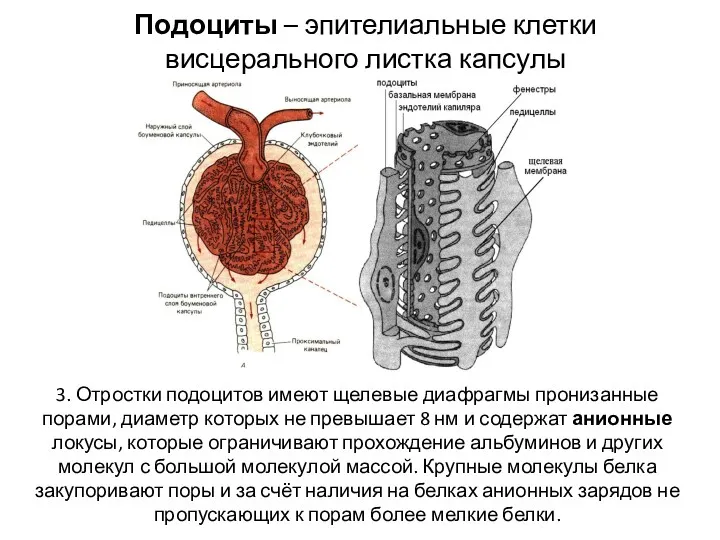
Подоциты – эпителиальные клетки висцерального листка капсулы
3. Отростки подоцитов имеют
Подоциты – эпителиальные клетки висцерального листка капсулы
3. Отростки подоцитов имеют

Состав ультрафильтрата (первичная моча)
Близок по концентрации веществ в плазме крови (кроме
Состав ультрафильтрата (первичная моча) Близок по концентрации веществ в плазме крови (кроме

В норме, за сутки, фильтруется до 5 г белка – в
В норме, за сутки, фильтруется до 5 г белка – в
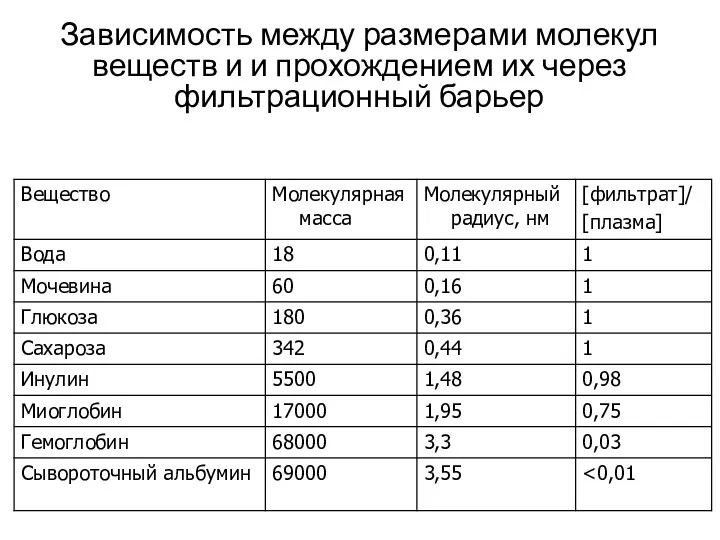
Зависимость между размерами молекул веществ и и прохождением их через фильтрационный
Зависимость между размерами молекул веществ и и прохождением их через фильтрационный
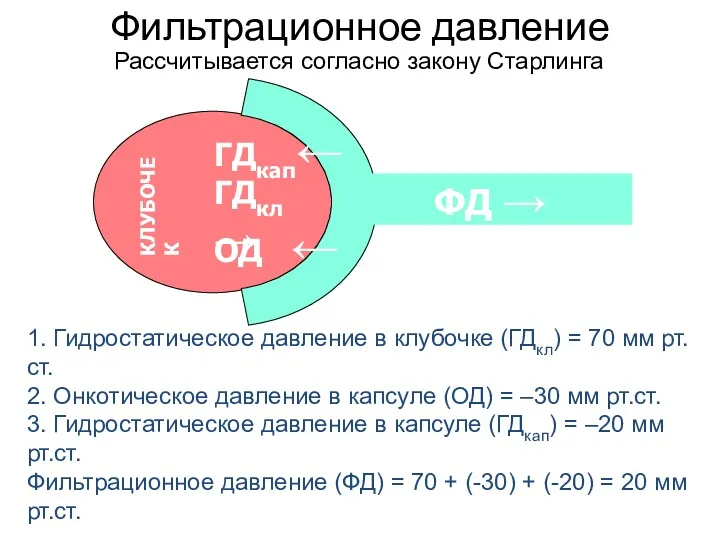
Фильтрационное давление
1. Гидростатическое давление в клубочке (ГДкл) = 70 мм рт.ст.
2.
Фильтрационное давление
1. Гидростатическое давление в клубочке (ГДкл) = 70 мм рт.ст. 2.
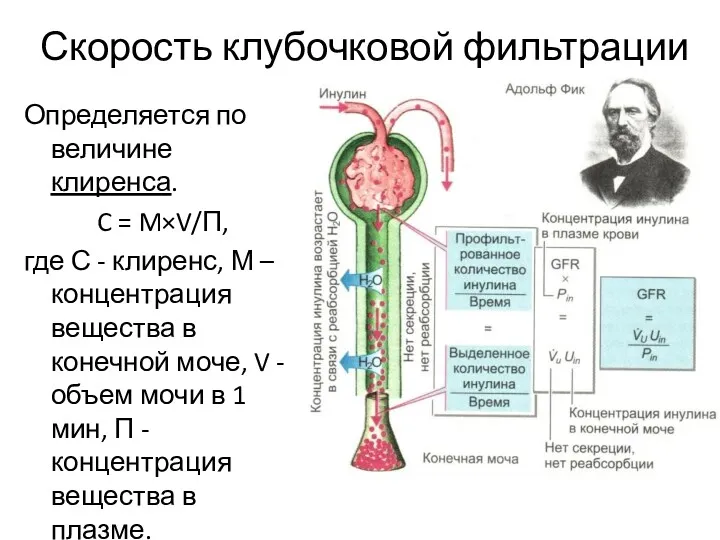
Скорость клубочковой фильтрации
Определяется по величине клиренса.
C = M×V/П,
где С -
Скорость клубочковой фильтрации
Определяется по величине клиренса.
C = M×V/П,
где С -

Клиренс – это скорость, с которой объём плазмы полностью очищается от
Клиренс – это скорость, с которой объём плазмы полностью очищается от
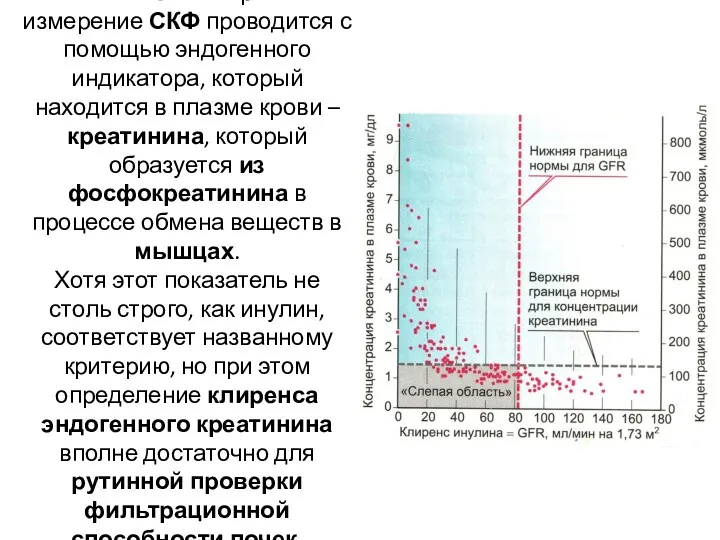
В клинической практике измерение СКФ проводится с помощью эндогенного индикатора, который
В клинической практике измерение СКФ проводится с помощью эндогенного индикатора, который
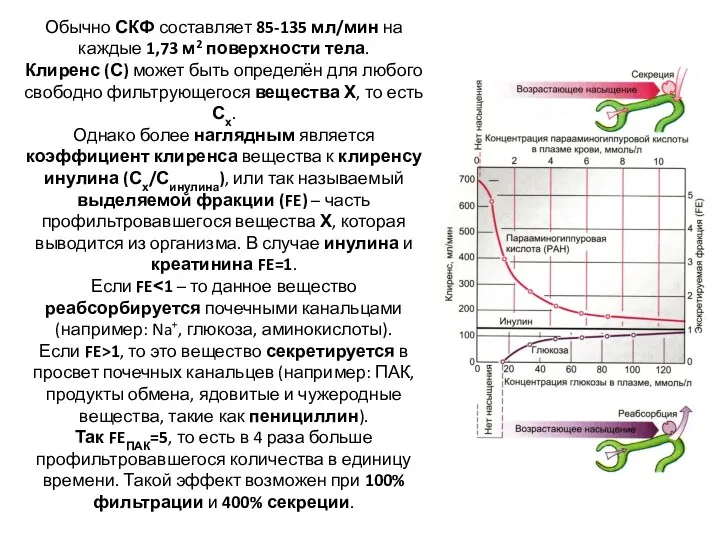
Обычно СКФ составляет 85-135 мл/мин на каждые 1,73 м2 поверхности тела.
Обычно СКФ составляет 85-135 мл/мин на каждые 1,73 м2 поверхности тела.

ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ОБЪЁМ ФИЛЬТРАТА
Фильтрационное давление – движущая сила перехода жидкости
ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ОБЪЁМ ФИЛЬТРАТА
Фильтрационное давление – движущая сила перехода жидкости
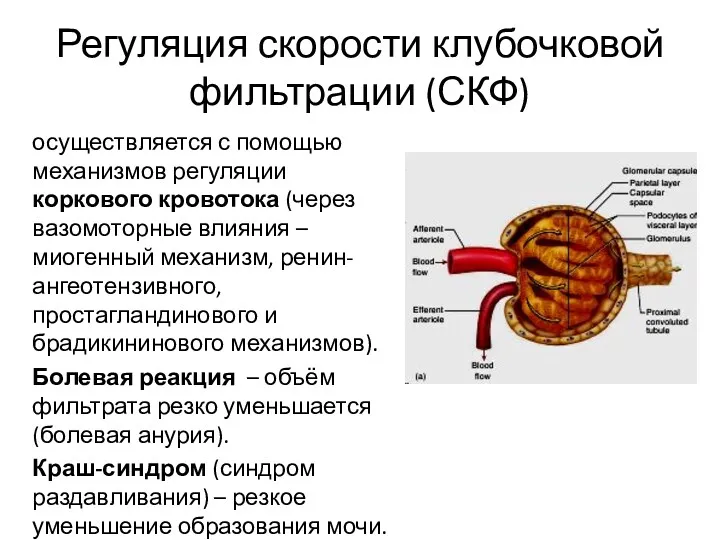
Регуляция скорости клубочковой фильтрации (СКФ)
осуществляется с помощью механизмов регуляции коркового кровотока
Регуляция скорости клубочковой фильтрации (СКФ)
осуществляется с помощью механизмов регуляции коркового кровотока
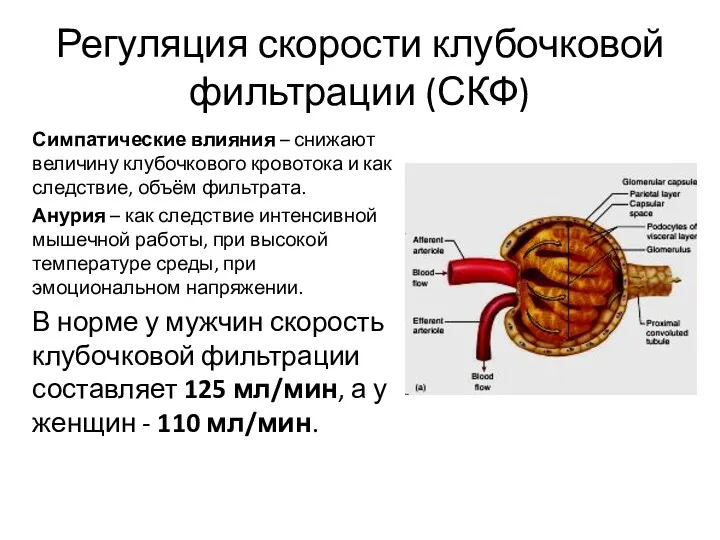
Регуляция скорости клубочковой фильтрации (СКФ)
Симпатические влияния – снижают величину клубочкового кровотока
Регуляция скорости клубочковой фильтрации (СКФ)
Симпатические влияния – снижают величину клубочкового кровотока

Мочевина – образуется в результате катаболизма белка. За сутки, из 100
Мочевина – образуется в результате катаболизма белка. За сутки, из 100

Канальцевая реабсорбция
это процесс обратного всасывания воды и ряда растворенных в ней
Канальцевая реабсорбция
это процесс обратного всасывания воды и ряда растворенных в ней

Вещества, содержащиеся в моче, по степени их реабсорбции, делятся на:
Беспороговые (непороговые)
Вещества, содержащиеся в моче, по степени их реабсорбции, делятся на: Беспороговые (непороговые)

Избирательность реабсорбции
Физиологически ценные, жизненно важные вещества реабсорбируются полностью: витамины, аминокислоты,
Избирательность реабсорбции
Физиологически ценные, жизненно важные вещества реабсорбируются полностью: витамины, аминокислоты,

Реабсорбция подразделяется на:
Облигатную (обязательную), нерегулируемую – реабсорбируется фактически та же
Реабсорбция подразделяется на: Облигатную (обязательную), нерегулируемую – реабсорбируется фактически та же

Относительный объём проксимальной (облигатной) реабсорбции – около 2/3 (65 %) от
Относительный объём проксимальной (облигатной) реабсорбции – около 2/3 (65 %) от

Облигатная реабсорбция веществ из проксимальных канальцев происходит за счёт первичной реабсорбции
Облигатная реабсорбция веществ из проксимальных канальцев происходит за счёт первичной реабсорбции

Факультативная реабсорбция – происходит в основном в дистальных извитых канальцах и
Факультативная реабсорбция – происходит в основном в дистальных извитых канальцах и
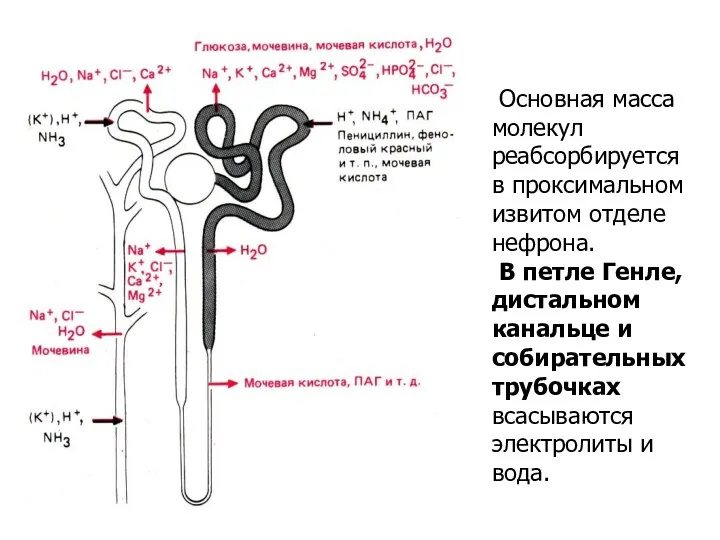
Основная масса молекул реабсорбируется в проксимальном извитом отделе нефрона.
Основная масса молекул реабсорбируется в проксимальном извитом отделе нефрона.
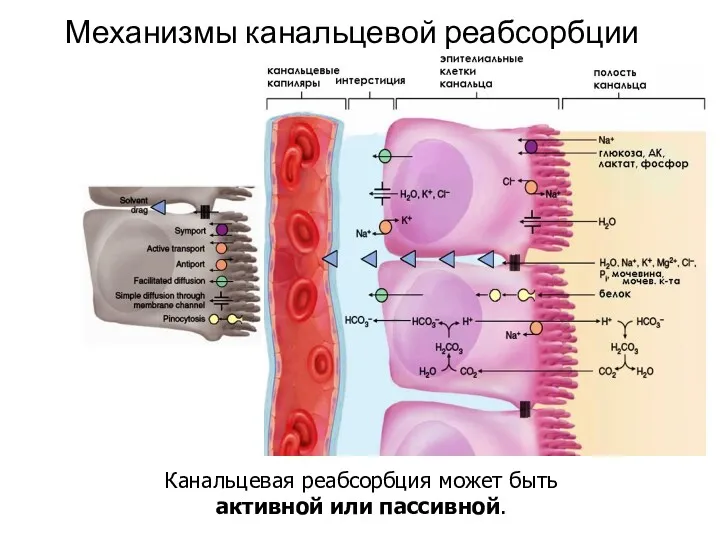
Механизмы канальцевой реабсорбции
Канальцевая реабсорбция может быть
активной или пассивной.
Механизмы канальцевой реабсорбции
Канальцевая реабсорбция может быть
активной или пассивной.

Пассивный транспорт осуществляется по принципу как облегчённой диффузии (реабсорбция Н2О, СО2,
Пассивный транспорт осуществляется по принципу как облегчённой диффузии (реабсорбция Н2О, СО2,

Механизм канальцевой реабсорбции
Обеспечивается: активным и пассивным транспортом.
Виды активного транспорта:
Механизм канальцевой реабсорбции
Обеспечивается: активным и пассивным транспортом.
Виды активного транспорта:

Реабсорбция глюкозы
Глюкоза полностью реабсорбируется в проксимальном канальце путем вторично-активного транспорта с
Реабсорбция глюкозы
Глюкоза полностью реабсорбируется в проксимальном канальце путем вторично-активного транспорта с

Реабсорбция аминокислот (АК)
АК полностью реабсорбируются в проксимальном канальце путем вторично-активного транспорта
Реабсорбция аминокислот (АК)
АК полностью реабсорбируются в проксимальном канальце путем вторично-активного транспорта

В проксимальном отделе нефрона на апикальной мембране эпителия (подобно эпителию тонкого
В проксимальном отделе нефрона на апикальной мембране эпителия (подобно эпителию тонкого

Внутри клеток эпителия концентрация АК увеличивается в несколько раз по сравнению
Внутри клеток эпителия концентрация АК увеличивается в несколько раз по сравнению

Реабсорбция аминокислот
Аминокислоты (АК) – реабсорбируются более чем на 99% в
Реабсорбция аминокислот Аминокислоты (АК) – реабсорбируются более чем на 99% в

Реабсорбция белков
Белки реабсорбируются полностью в проксимальном канальце путем пиноцитоза. В дальнейшем
Реабсорбция белков
Белки реабсорбируются полностью в проксимальном канальце путем пиноцитоза. В дальнейшем

Механизм эндоцитоза белков
За счёт наличия на щёточной кайме рецепторного мегалин-кубилинового
Механизм эндоцитоза белков
За счёт наличия на щёточной кайме рецепторного мегалин-кубилинового
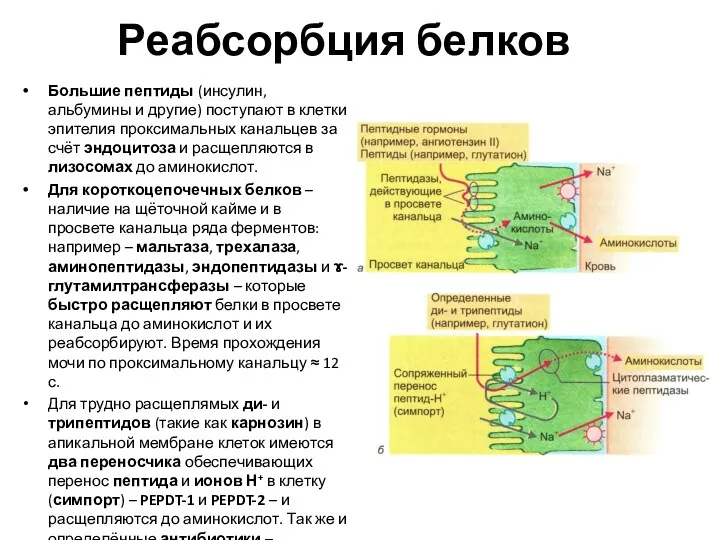
Реабсорбция белков
Большие пептиды (инсулин, альбумины и другие) поступают в клетки
Реабсорбция белков
Большие пептиды (инсулин, альбумины и другие) поступают в клетки

Белок в моче
В ультрафильтрате обнаруживается ряд белков такие как альбумины
Белок в моче
В ультрафильтрате обнаруживается ряд белков такие как альбумины

Реабсорбция мочевины
Мочевина – это неполярное низкомолекулярное соединение, поэтому легко проникает через
Реабсорбция мочевины
Мочевина – это неполярное низкомолекулярное соединение, поэтому легко проникает через

Реабсорбция
слабых кислот и оснований
Реабсорция зависит от pH внутриканальцевой жидкости.
Слабые основания
Реабсорбция
слабых кислот и оснований
Реабсорция зависит от pH внутриканальцевой жидкости.
Слабые основания

Реабсорбция воды по всему протяжению почечных канальцев происходит только пассивно. Из
Реабсорбция воды по всему протяжению почечных канальцев происходит только пассивно. Из

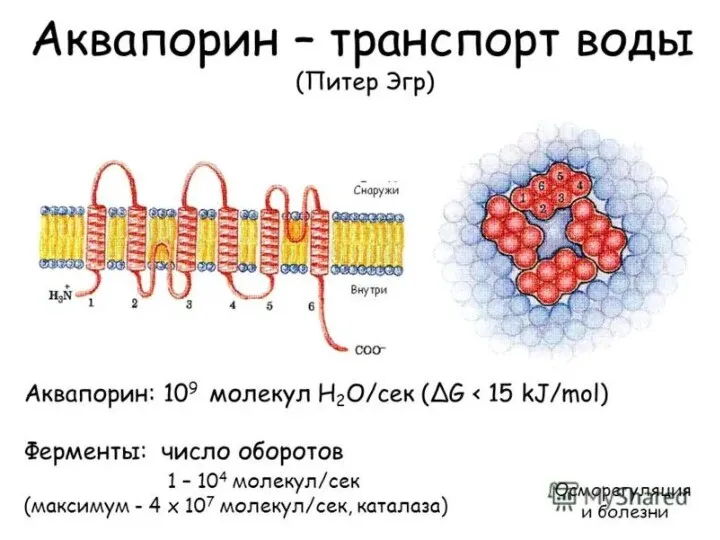
Аквапорины (AQP) — семейство мембранных белков-пор для воды. Идентифицировано 10 аквапоринов. AQP3,
Аквапорины (AQP) — семейство мембранных белков-пор для воды. Идентифицировано 10 аквапоринов. AQP3,
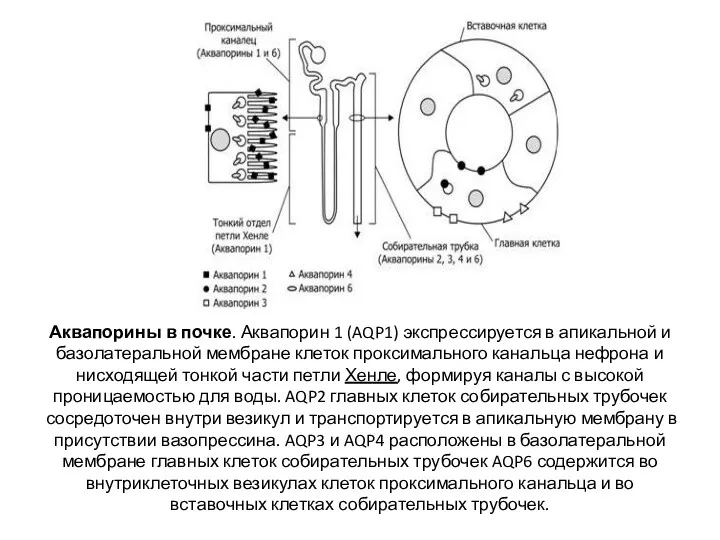
Аквапорины в почке. Аквапорин 1 (AQP1) экспрессируется в апикальной и базолатеральной
Аквапорины в почке. Аквапорин 1 (AQP1) экспрессируется в апикальной и базолатеральной
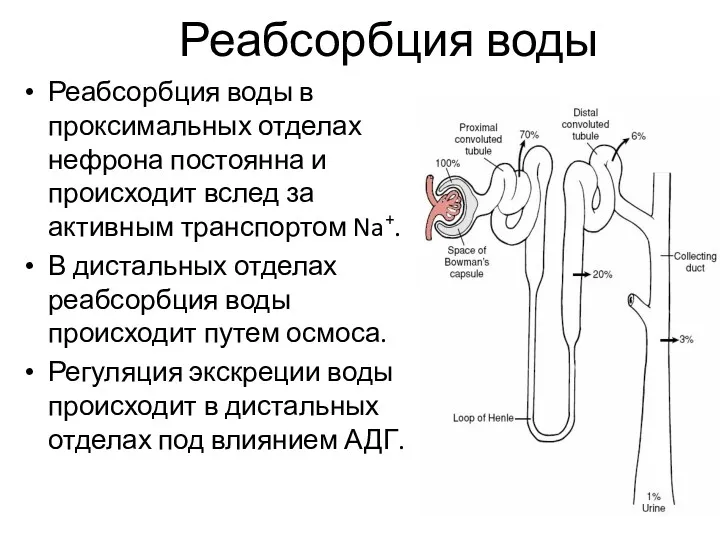
Реабсорбция воды
Реабсорбция воды в проксимальных отделах нефрона постоянна и происходит
Реабсорбция воды
Реабсорбция воды в проксимальных отделах нефрона постоянна и происходит


Реабсорбция электролитов
1. Реабсорбция Na+ идет в 2 этапа:
а. выкачивание Na+
Реабсорбция электролитов
1. Реабсорбция Na+ идет в 2 этапа:
а. выкачивание Na+

ПРОТИВОТОЧНО-МНОЖИТЕЛЬНЫЙ МЕХАНИЗМ КОНЦЕНТРИРОВАНИЯ МОЧИ
ПРОТИВОТОЧНО-МНОЖИТЕЛЬНЫЙ МЕХАНИЗМ КОНЦЕНТРИРОВАНИЯ МОЧИ

Эта система объединяет в себе 3 взаимодополняющих звена, каждое из которых
Эта система объединяет в себе 3 взаимодополняющих звена, каждое из которых
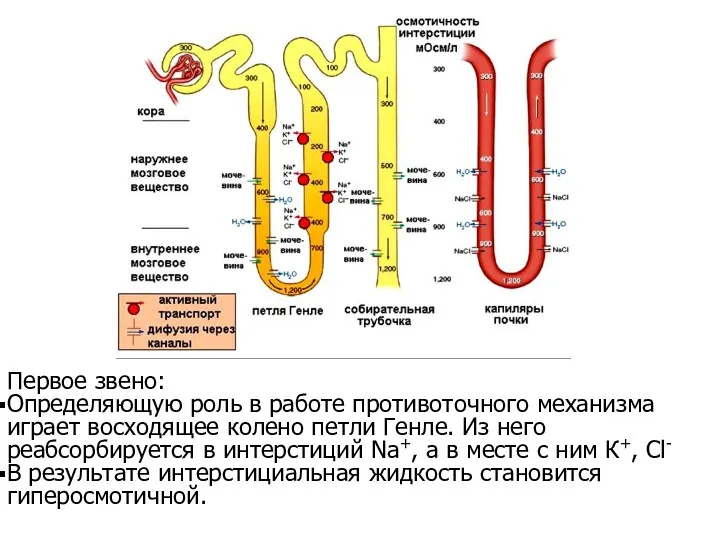
Первое звено:
Определяющую роль в работе противоточного механизма играет восходящее колено петли
Первое звено:
Определяющую роль в работе противоточного механизма играет восходящее колено петли
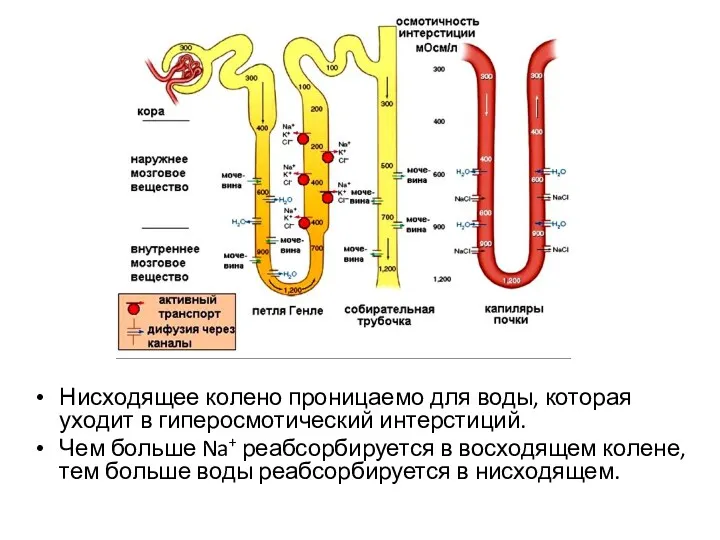
Нисходящее колено проницаемо для воды, которая уходит в гиперосмотический интерстиций.
Чем
Нисходящее колено проницаемо для воды, которая уходит в гиперосмотический интерстиций.
Чем
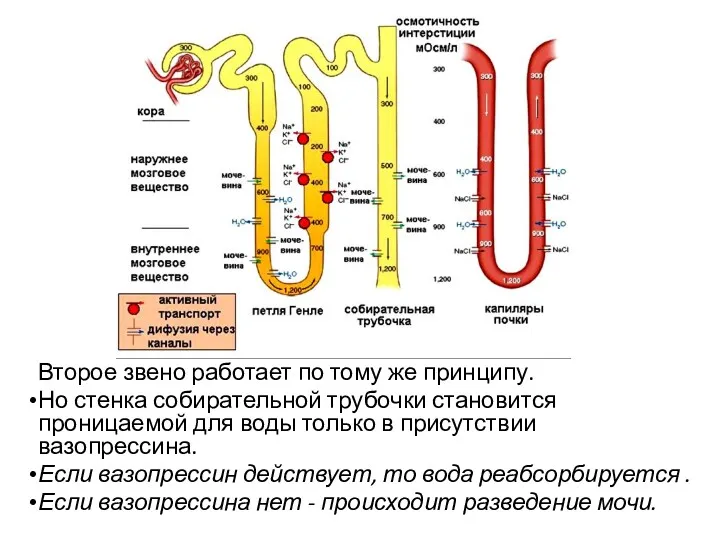
Второе звено работает по тому же принципу.
Но стенка собирательной трубочки становится
Второе звено работает по тому же принципу.
Но стенка собирательной трубочки становится
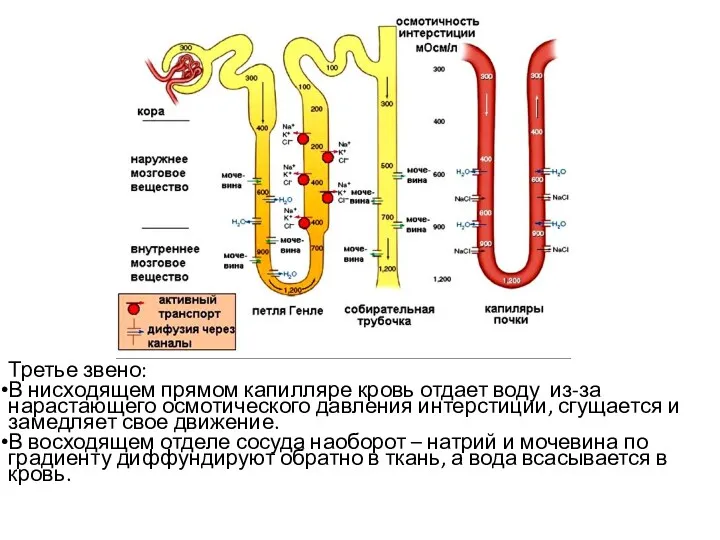
Третье звено:
В нисходящем прямом капилляре кровь отдает воду из-за нарастающего осмотического
Третье звено:
В нисходящем прямом капилляре кровь отдает воду из-за нарастающего осмотического
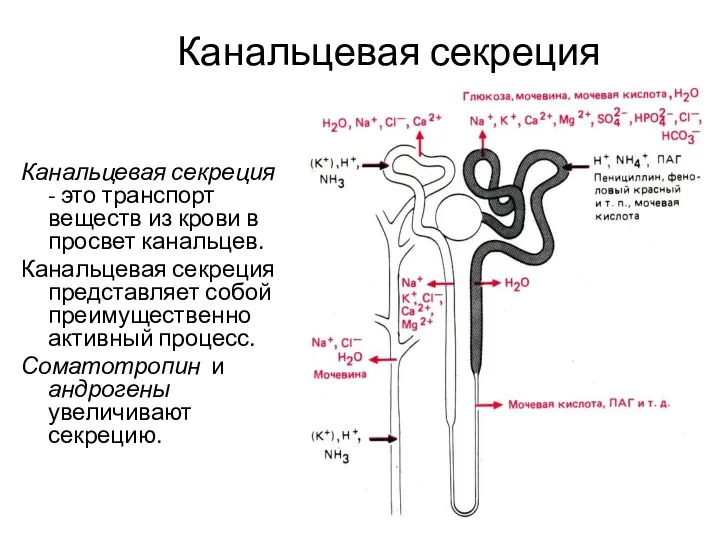
Канальцевая секреция
Канальцевая секреция - это транспорт веществ из крови в просвет
Канальцевая секреция
Канальцевая секреция - это транспорт веществ из крови в просвет

Регуляция водно-солевого баланса
Повышение осмотического давления крови
возбуждение осморецепторов (в области супраоптического
Регуляция водно-солевого баланса
Повышение осмотического давления крови
возбуждение осморецепторов (в области супраоптического

Инсулин. Недостаток этого гормона приводит к гипергликемии, глюкозурии, увеличению осмотического давления
Инсулин. Недостаток этого гормона приводит к гипергликемии, глюкозурии, увеличению осмотического давления

Регуляция ионного состава крови
Регуляция ионного состава крови

Na+
Альдостерон усиливает реабсорбцию Na+ в дистальных отделах канальцев и в собирательных
Na+
Альдостерон усиливает реабсорбцию Na+ в дистальных отделах канальцев и в собирательных

.Различные лекарственные средства (диуретики), подавляя реабсорбцию Na+, увеличивают экскрецию Na+ и
.Различные лекарственные средства (диуретики), подавляя реабсорбцию Na+, увеличивают экскрецию Na+ и

K+
Альдостерон усиливает секрецию K+ в дистальном отделе канальцев и собирательных трубочках.
K+
Альдостерон усиливает секрецию K+ в дистальном отделе канальцев и собирательных трубочках.
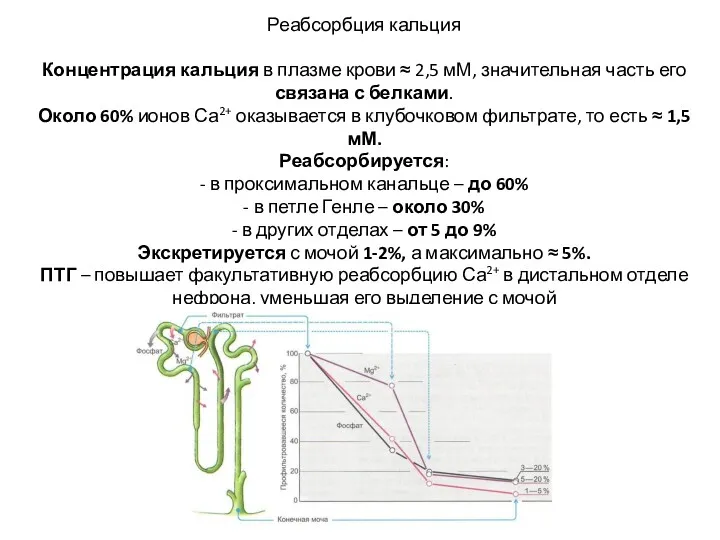
Реабсорбция кальция
Концентрация кальция в плазме крови ≈ 2,5 мМ, значительная
Реабсорбция кальция Концентрация кальция в плазме крови ≈ 2,5 мМ, значительная
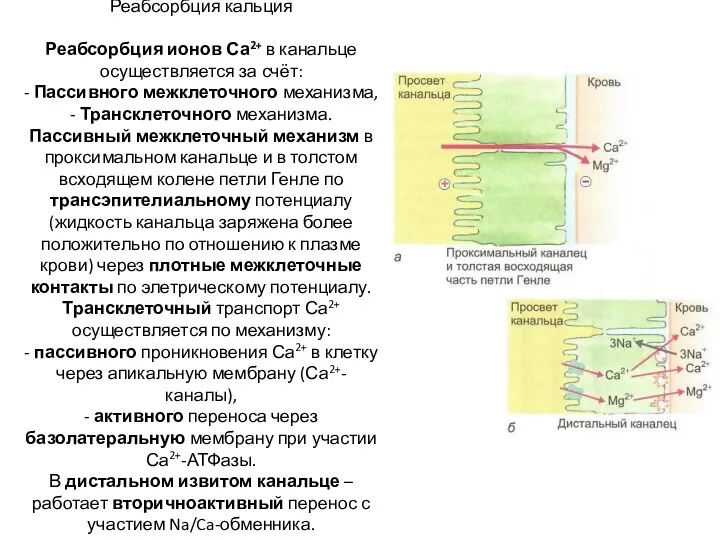
Реабсорбция кальция
Реабсорбция ионов Са2+ в канальце осуществляется за счёт:
-
Реабсорбция кальция Реабсорбция ионов Са2+ в канальце осуществляется за счёт: -

Ca2+
Паратгормон увеличивает реабсорбцию Ca2+ и высвобождение Ca 2+ из костей, что
Ca2+
Паратгормон увеличивает реабсорбцию Ca2+ и высвобождение Ca 2+ из костей, что

Cl -
Альдостерон участвует в регуляции уровня Cl- в плазме крови.
При
Cl -
Альдостерон участвует в регуляции уровня Cl- в плазме крови.
При

Реабсорбция Pi продолжается и дистальнее проксимального канальца, так что в итоге
Реабсорбция Pi продолжается и дистальнее проксимального канальца, так что в итоге

Реабсорбция фосфата в проксимальном канальце
Неорганический фосфат (Pi) – содержится в
Реабсорбция фосфата в проксимальном канальце Неорганический фосфат (Pi) – содержится в

Паратиреоидный гормон (ПТГ) (паращитовидные железы), действие которого опосредуется через цАМФ или
Паратиреоидный гормон (ПТГ) (паращитовидные железы), действие которого опосредуется через цАМФ или

Поддержание кислотно-щелочного равновесия
Поддержание кислотно-щелочного равновесия

Основания с большей скоростью экскретируются с кислой мочой, а кислоты -
Основания с большей скоростью экскретируются с кислой мочой, а кислоты -
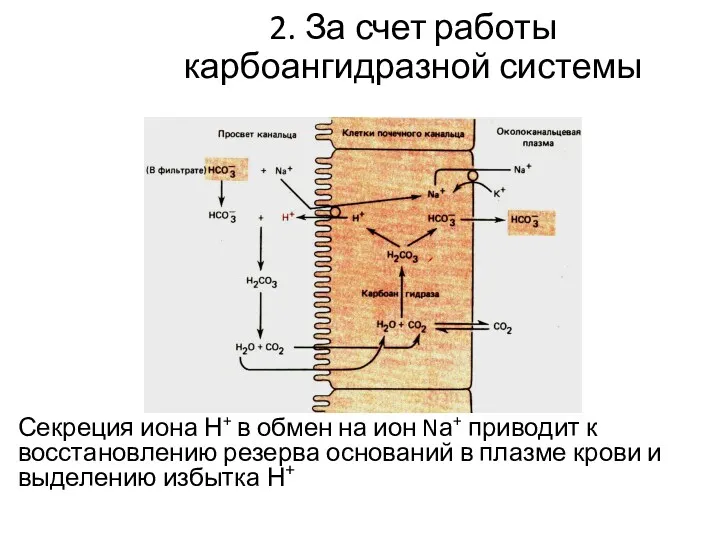
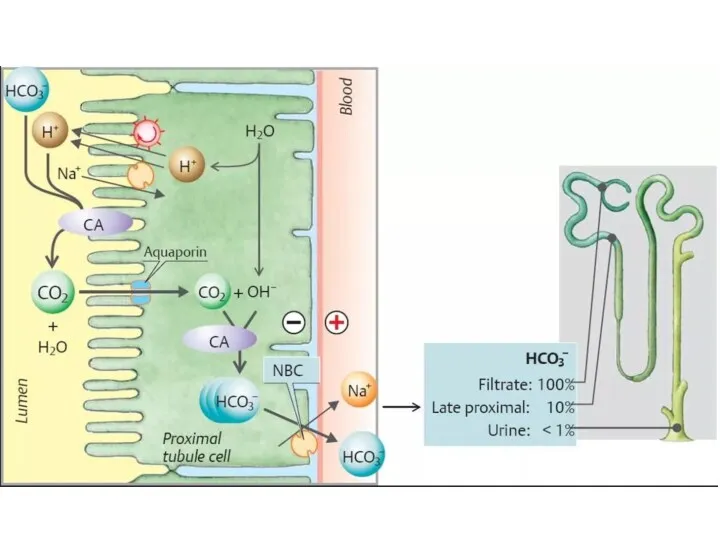
2. За счет работы карбоангидразной системы
Секреция иона Н+ в обмен на
2. За счет работы карбоангидразной системы
Секреция иона Н+ в обмен на
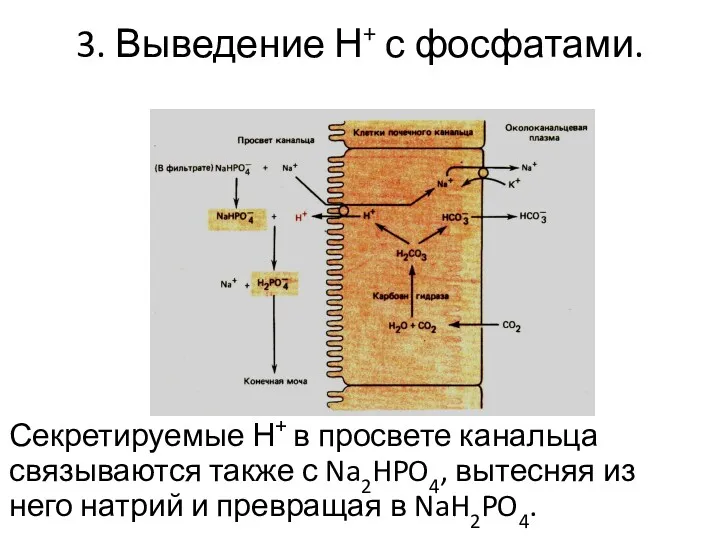
3. Выведение Н+ с фосфатами.
Секретируемые Н+ в просвете канальца связываются также
3. Выведение Н+ с фосфатами.
Секретируемые Н+ в просвете канальца связываются также
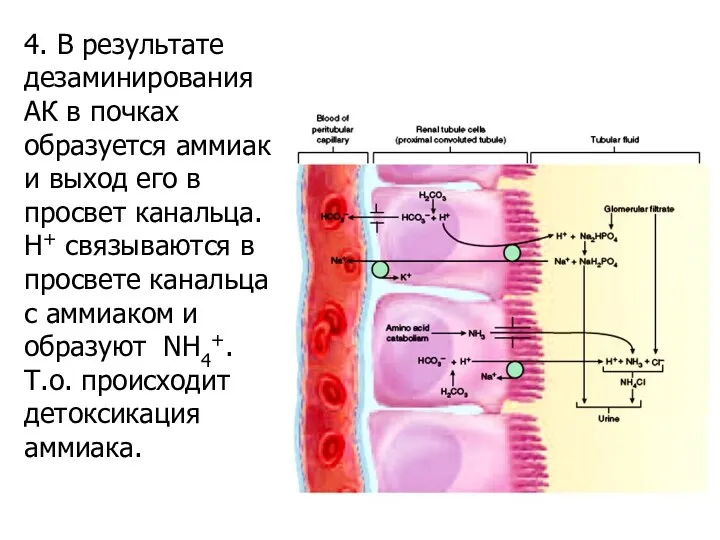
4. В результате дезаминирования АК в почках образуется аммиак и выход
4. В результате дезаминирования АК в почках образуется аммиак и выход

Нервная регуляция мочеобразования.
Морфологические основы.
Симпатическая эфферентная иннервация почек осуществляется из Th5-L3 сегментов
Нервная регуляция мочеобразования. Морфологические основы. Симпатическая эфферентная иннервация почек осуществляется из Th5-L3 сегментов

Физиологические основы регуляции
фильтрации
Возбуждение симпатической нервной системы уменьшает фильтрацию, за счёт
Физиологические основы регуляции фильтрации Возбуждение симпатической нервной системы уменьшает фильтрацию, за счёт

Регуляция величины АД
Регуляция величины АД
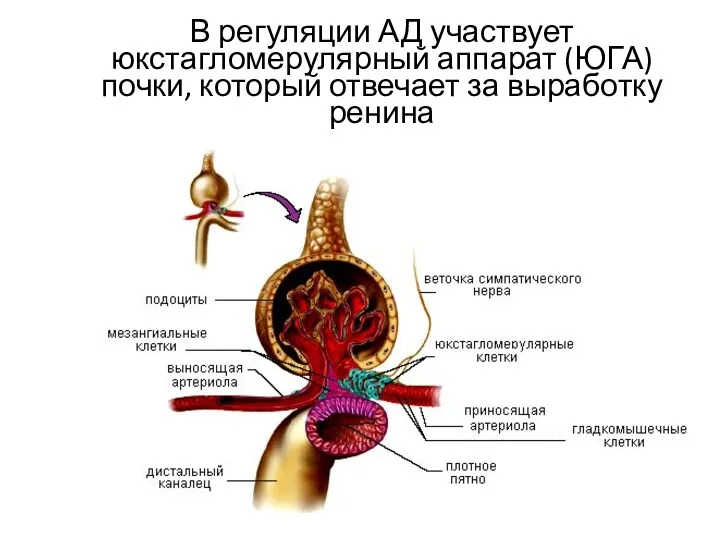
В регуляции АД участвует юкстагломерулярный аппарат (ЮГА) почки, который отвечает за
В регуляции АД участвует юкстагломерулярный аппарат (ЮГА) почки, который отвечает за

Часть начального отдела дистального извитого канальца очень плотно прилегает к корню
Часть начального отдела дистального извитого канальца очень плотно прилегает к корню

Клетки ЮГА иннервируются симпатическими волокнами и содержат β- и α-ADR. Активация
Клетки ЮГА иннервируются симпатическими волокнами и содержат β- и α-ADR. Активация

Юкстагломерулярный комплекс работает по принципу обратной отрицательной связи:
1. при снижении
Юкстагломерулярный комплекс работает по принципу обратной отрицательной связи: 1. при снижении

Регуляция выработки ренина
При понижении давления в приносящей артериоле в гранулярных клетках
Регуляция выработки ренина
При понижении давления в приносящей артериоле в гранулярных клетках

Возбуждение β-адренорецепторов гранулярных клеток приводит к усилению секреции ренина, а α-адренорецепторов
Возбуждение β-адренорецепторов гранулярных клеток приводит к усилению секреции ренина, а α-адренорецепторов
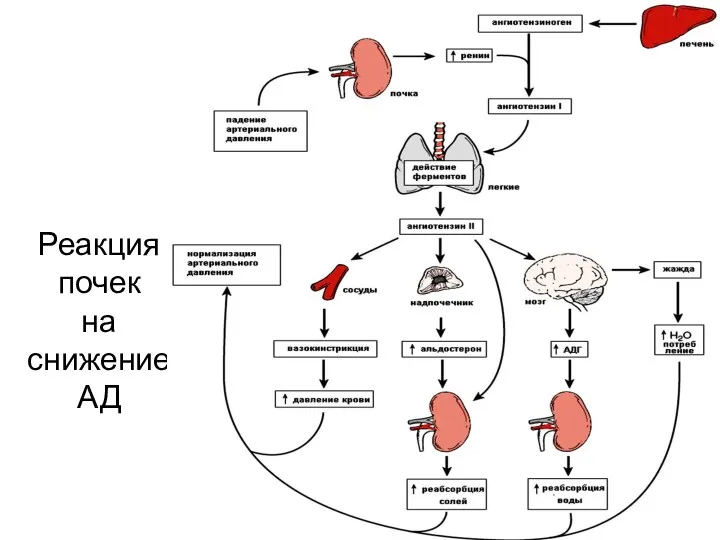
Реакция
почек
на снижение
АД
Реакция
почек
на снижение
АД

Реакция почек на увеличение АД (внутрисистемная и внесистемная)
Реакция почек на увеличение АД (внутрисистемная и внесистемная)
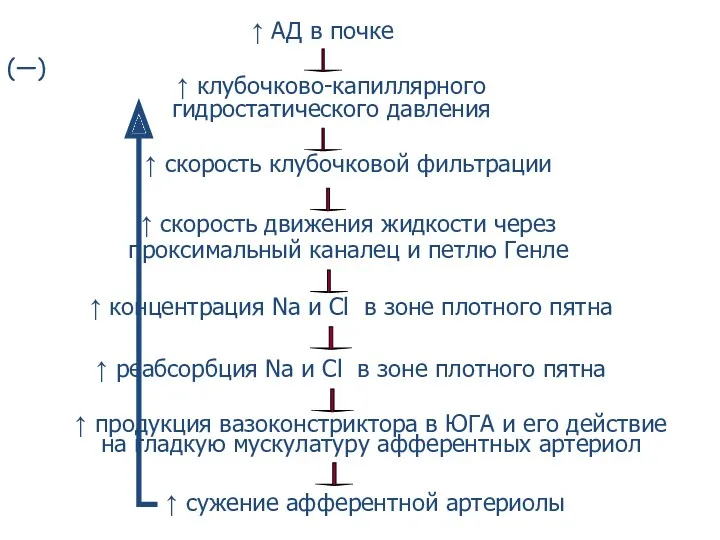
↑ АД в почке
↑ клубочково-капиллярного
гидростатического давления
↑ скорость клубочковой фильтрации
↑ скорость движения
↑ АД в почке
↑ клубочково-капиллярного
гидростатического давления
↑ скорость клубочковой фильтрации
↑ скорость движения
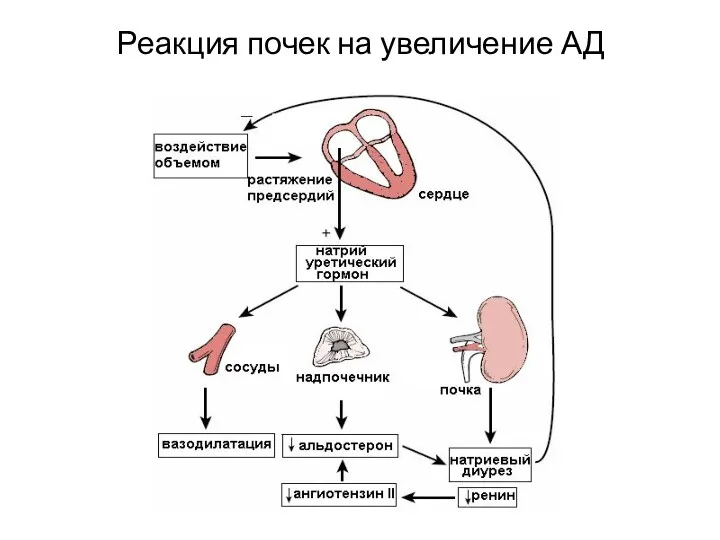
Реакция почек на увеличение АД
Реакция почек на увеличение АД

Нервная регуляция мочеобразования
Нервная регуляция мочеобразования

Влияние СНС:
При сужении приносящих артериол
→ ↓ фильтрация.
Сужение выносящих артериол
Влияние СНС:
При сужении приносящих артериол
→ ↓ фильтрация.
Сужение выносящих артериол

При болевых раздражениях диурез рефлекторно уменьшается вплоть до анурии. Механизм:
- сужение
При болевых раздражениях диурез рефлекторно уменьшается вплоть до анурии. Механизм:
- сужение

Раздражение ПСНС вызывает усиление реабсорбции глюкозы и секреции органических кислот.
Условно-рефлекторно
Раздражение ПСНС вызывает усиление реабсорбции глюкозы и секреции органических кислот.
Условно-рефлекторно

Непочечные органы выделения:
Непочечные органы выделения:
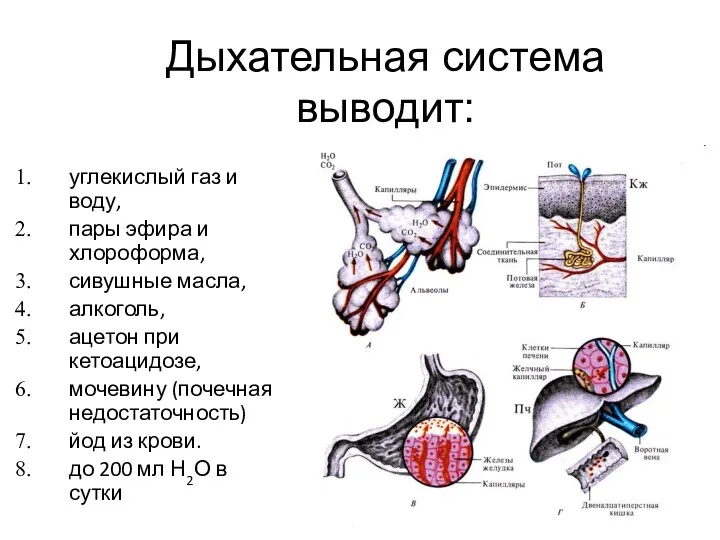
Дыхательная система
выводит:
углекислый газ и воду,
пары эфира и хлороформа,
сивушные масла,
алкоголь,
ацетон при кетоацидозе,
мочевину
Дыхательная система
выводит:
углекислый газ и воду,
пары эфира и хлороформа,
сивушные масла,
алкоголь,
ацетон при кетоацидозе,
мочевину
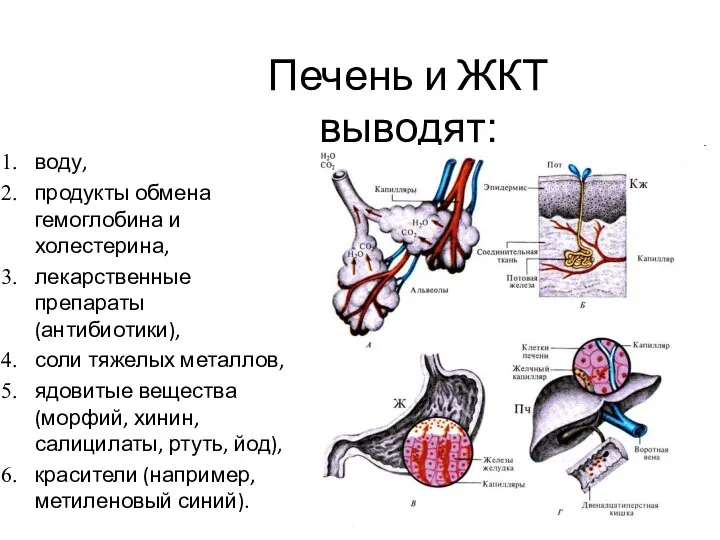
Печень и ЖКТ
выводят:
воду,
продукты обмена гемоглобина и холестерина,
лекарственные препараты (антибиотики),
соли тяжелых
Печень и ЖКТ
выводят:
воду,
продукты обмена гемоглобина и холестерина,
лекарственные препараты (антибиотики),
соли тяжелых
Потовые железы удаляют:
воду,
мочевину,
мочевую кислоту,
креатинин,
молочную кислоту,
соли щелочных
Потовые железы удаляют:
воду,
мочевину,
мочевую кислоту,
креатинин,
молочную кислоту,
соли щелочных
свободные жирные и неомыляемые кислоты,
продукты обмена половых гормонов.
Сальные железы
свободные жирные и неомыляемые кислоты,
продукты обмена половых гормонов.
Сальные железы

Инструментальные методы исследования.
Обзорный рентгеновский снимок определит положение, величину и форму почек,
Инструментальные методы исследования.
Обзорный рентгеновский снимок определит положение, величину и форму почек,
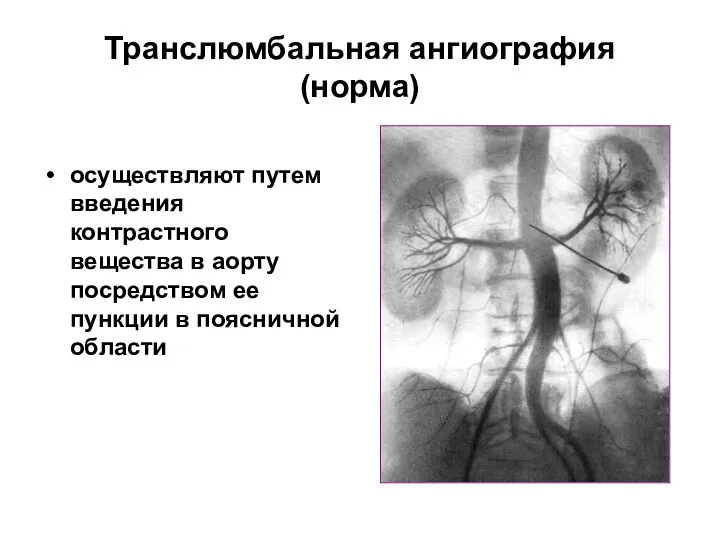
Транслюмбальная ангиография (норма)
осуществляют путем введения контрастного вещества в аорту посредством ее
Транслюмбальная ангиография (норма)
осуществляют путем введения контрастного вещества в аорту посредством ее
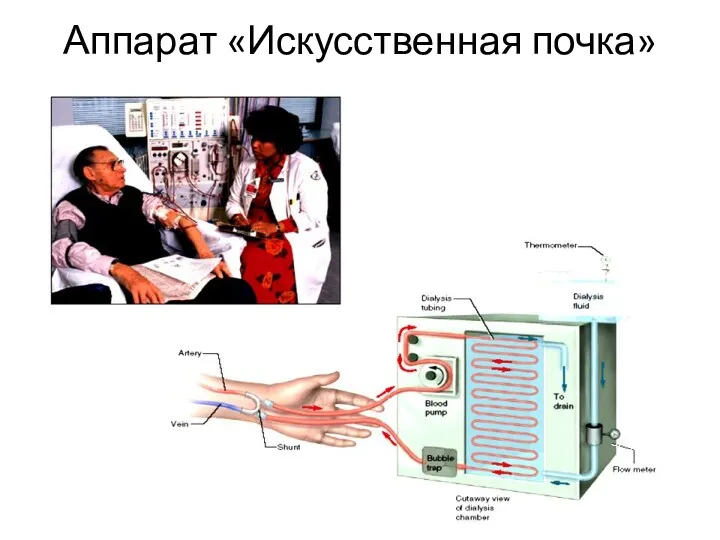
Аппарат «Искусственная почка»
Аппарат «Искусственная почка»
Аппарат «Искусственная почка»
Американские ученые разработали диализный аппарат ("искусственную почку"), который можно
Аппарат «Искусственная почка»
Американские ученые разработали диализный аппарат ("искусственную почку"), который можно
ФИЗИОЛОГИЯ МОЧЕВЫДЕЛЕНИЯ
ФИЗИОЛОГИЯ МОЧЕВЫДЕЛЕНИЯ

Морфологические основы
В процессе мочеобразования моча, из собирательных трубочек по выводным протокам
Морфологические основы
В процессе мочеобразования моча, из собирательных трубочек по выводным протокам

Мускулатура мочевого пузыря – трёхслойная:
наружный и внутренний слои – продольные,
средний
Мускулатура мочевого пузыря – трёхслойная:
наружный и внутренний слои – продольные,
средний

Позыв к мочеиспусканию возникает при 150-200 мл. Наполнение до 300 мл
Позыв к мочеиспусканию возникает при 150-200 мл. Наполнение до 300 мл

Эфферентные волокна – идут от пирамидных нейронов КБП к различным структурам
Эфферентные волокна – идут от пирамидных нейронов КБП к различным структурам
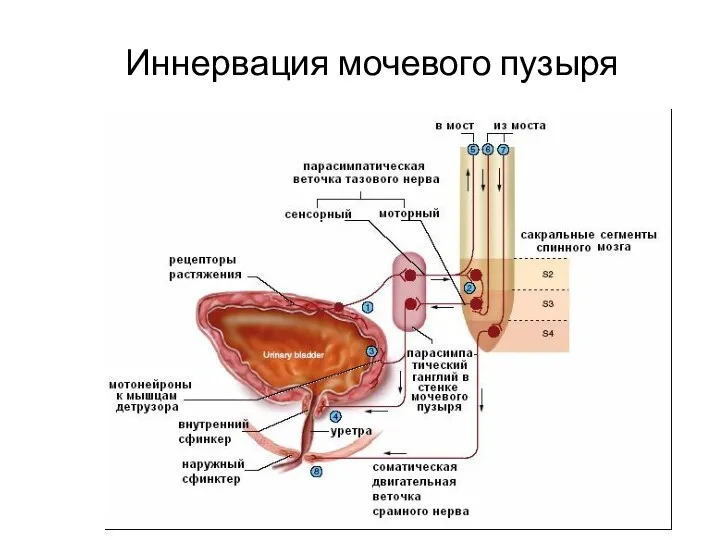
Иннервация мочевого пузыря
Иннервация мочевого пузыря

Парасимпатическая система оказывает стимулирующее влияние на сокращение гладких мышц мочевого пузыря.
Парасимпатическая система оказывает стимулирующее влияние на сокращение гладких мышц мочевого пузыря.

Повышение активности симпатической системы – создаёт условия для наполнения мочевого пузыря.
Повышение активности симпатической системы – создаёт условия для наполнения мочевого пузыря.
Мочевыделение
Физиологические основы.
Мочевыделение
Физиологические основы.

Центры контроля мочеиспускания находятся: – коркового контроля – в первой сенсорной
Центры контроля мочеиспускания находятся: – коркового контроля – в первой сенсорной

Эфференты парасимпатической системы вызывают сокращение гладких мышц мочевого пузыря и его
Эфференты парасимпатической системы вызывают сокращение гладких мышц мочевого пузыря и его

Таким образом:
корковые нейроны и средний мозг оказывают тормозящее влияние на
Таким образом:
корковые нейроны и средний мозг оказывают тормозящее влияние на
 рH-метрия
рH-метрия Орталық жүйке жүйесі зақымдалып өмірге қауіп төнген жағдайда жедел көмек көрсету және диагностика алгоритмі
Орталық жүйке жүйесі зақымдалып өмірге қауіп төнген жағдайда жедел көмек көрсету және диагностика алгоритмі Группы здоровья детей в детском саду
Группы здоровья детей в детском саду Здоровьесберегающие технологии на уроках физической культуры
Здоровьесберегающие технологии на уроках физической культуры Механикалық жұмыс істейтін ортодонтиялық апараттар қолдануға арналған көрсеткіштер
Механикалық жұмыс істейтін ортодонтиялық апараттар қолдануға арналған көрсеткіштер Противоопухолевый иммунитет
Противоопухолевый иммунитет Заболевания шеи и пищевода
Заболевания шеи и пищевода Leptospirosis
Leptospirosis Антисептики і дезінфекційні. Окисники. Розчин перекису водню
Антисептики і дезінфекційні. Окисники. Розчин перекису водню Системная склеродермия
Системная склеродермия Синдром пангипопитуитаризма
Синдром пангипопитуитаризма Гемофильная инфекция
Гемофильная инфекция Вплив тютюнопаління, алкоголю, наркотиків і токсинів на нервову систему та поведінку людини
Вплив тютюнопаління, алкоголю, наркотиків і токсинів на нервову систему та поведінку людини Аномалии конституции. Диатез. Синдром внезапной смерти
Аномалии конституции. Диатез. Синдром внезапной смерти Лекарственные средства, синапсы
Лекарственные средства, синапсы Pulmonary tuberculosis
Pulmonary tuberculosis Организация противоэпидемических мероприятий при чрезвычайных ситуациях
Организация противоэпидемических мероприятий при чрезвычайных ситуациях Внутрибольничные инфекции. Стрептококки
Внутрибольничные инфекции. Стрептококки Компания Буарон. Производство гомеопатических препаратов
Компания Буарон. Производство гомеопатических препаратов Предмет и задачи судебной психиатрии
Предмет и задачи судебной психиатрии Предмет логопедии, ее становление как интегративной отрасли знаний
Предмет логопедии, ее становление как интегративной отрасли знаний Неправильное положение плода. Тема 6
Неправильное положение плода. Тема 6 Диагностика отосклероза по данным МСКТ
Диагностика отосклероза по данным МСКТ Пародонт ауруы бар науқастарды металды крамикалық протездермен емдеу ерекшеліктері
Пародонт ауруы бар науқастарды металды крамикалық протездермен емдеу ерекшеліктері Мази и линименты
Мази и линименты Диагностика заболеваний кожи
Диагностика заболеваний кожи Гемолитическая болезнь плода и новорожденного. Повышение непрямого билирубина
Гемолитическая болезнь плода и новорожденного. Повышение непрямого билирубина Антимикробные препараты (антибиотики)
Антимикробные препараты (антибиотики)