- Иммортализация (неограниченный репликативный потенциал) опухолевых клеток. Лекция 3

Содержание
- 2. Неограниченное деление стволовых клеток опухолей Стимуляция деления, потеря чувствительности к рост-ингибирующим сигналам Иммортализация (приобретение бессмертия)
- 3. Старение организма (Ageing, senescence) – исчерпание пула стволовых клеток различных тканей “Старение” клеток (“Cell senescence”) –
- 4. Иммортализация – отмена репликативного клеточного старения Нормальные клетки могут быть иммортальны – эмбриональные стволовые клетки Старение
- 5. Феномен: “Репликативное старение” и “иммортализация” клеток in vitro Человек: 50-70 делений (Число Хейфлика; Hayflick & Moorhead,
- 6. Феномен: “Репликативное старение” и “иммортализация” клеток in vitro Человек: 50-70 делений Фибробласты кожи: Иммортализация генетически изме-
- 7. Иммортализация – отмена репликативного клеточного старения Опухолевые клетки иммортальны – перевиваемые линии клеток (асцит Эрлиха, HeLa
- 8. 60 делений 1018 клеток
- 9. Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
- 10. Наиболее общие признаки репликативного старения клеток Необратимый арест пролиферации (не может быть отменен физиологическими стимулами) Увеличение
- 11. Четыре функции клеточного старения Rodier and Campisi J Cell Biol. 2011
- 12. стадия М0 стадии М1, М2 Человек (эпителиоциты и др. клетки кроме фибробластов кожи): «Старение», кризис Мышь
- 13. Неадекватные условия культивирования (культуральный шок) ускоряют старение клеток
- 14. стадия М0 стадии М1, М2 Человек (эпителиоциты и др. клетки кроме фибробластов кожи): «Старение», кризис Мышь
- 15. Число делений 3 18 26 32 36 42 43 p21Cip1 p16Ink4А Adapted from R.Weinberg, The Biology
- 16. Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007) Неадекватные условия культивирования кератиноцитов человека
- 17. Роль р16Ink4a в «старении» гемопоэтических cтволовых клеток (ГСК) in vivo 1. В популяции отсортированных ГСК уровень
- 18. Репликативное старение стволовых и недифференцированных клеток in vivo Ключевой механизм – увеличение содержания p16Ink4a вследствие инактивации
- 19. p16Ink4a-positive cells shorten healthy lifespan Baker et al., Nature. 2016 doi: 10.1038/nature16932 Gil J and Withers
- 20. Раунды деления клеток Cdk4 Cdk2 Остановка в G1 P16INK4A PRC1/2 (Bmi1 и др.) ? Дефосфорилирование pRb
- 21. Локус INK4А кодирует два негомологичных белка, инактивирующих сигнальные пути, регулируемые pRb и p53
- 22. Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007) Эксперименты с введением гена большого
- 23. Раунды деления клеток p21Cip1 Остановка в G1, дифференцировка p16Ink4a Bmi1, Twist p53 p14Arf Отмена остановки клеточного
- 24. Другие функции Bmi1 Компонент PRC1 комплекса, участвующего в ремоделировании структуры хроматина и модификации гистонов Активирован во
- 25. ИММОРТАЛИЗАЦИЯ Отмена рост-инигибирующих сигналов и/или нечувствительность к их действию Высокая активность теломеразы, поддержание нормальной структуры концов
- 26. Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007) FISH на метафазных хромосомах Теломеры
- 27. Строение теломер Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
- 28. Проблема недорепликации концов ДНК Теломеры теряют 50-100 пар нуклеотидов за один раунд репликации
- 29. «Репликативное старение» (стадия М1), остановка деления как при повреждениях ДНК «Генетическая катастрофа» кризис (стадия М2), слипание
- 30. кризис мортальные иммортальные Увеличивающееся число делений длина (kbp) Укорочение длины теломерной ДНК при культивировании лифоцитов человека
- 31. Как при укорочении/нарушении структуры 3’-конца теломерной ДНК индуцируется остановка клеточного цикла? Azzalin & Lingner, Nature, 2007,
- 32. Теломерный кризис и хромосомные перестройки Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
- 33. Механизмы поддержания постоянной длины теломер Активация теломеразы (эмбрионы, половые клетки, опухоли (85-90%)) ALT (Alternative Lengthening of
- 34. Введение гена hTERT повышает репликативный потенциал нормальных клеток (слева), а его доминантно-негативный мутант подавляет размножение опухолевых
- 35. Усовершенствованная теломерная модель клеточного старения Теломерные повторы ДНК Однонитчатая петля» ТTAGGGTTAGGG 5’ 3’ AATCCCAATCCC 5’ 3’
- 36. Возможная связь между дисфункцией теломер и воспалением Wang Z and Lieberman PM. RNA Biol.2016;13(8):690-5. Длинные некодирующие
- 37. + Теломераза (TERT) Поддержание структуры теломер Отмена остановки клеточного цикла - INK4a Е7 + Е6 +
- 38. Функция TERT в стволовых и недифференцированных клетках не ограничивается поддержанием структуры теломер Choi et al., 2008,
- 39. Функция hTERT в стволовых и недифференцированных клетках не ограничивается поддержанием структуры теломер Экспрессия TERT с дефектной
- 40. Активация сигнальных путей факторов роста, Wnt, NF-κB Усиление стволовых характеристик клетки Улучшение рибосомного биогенеза Подавление апоптоза
- 41. Почему теломераза активирована в опухолях? Kumar M. et al., 2016, Genes, 7:43
- 42. Механизмы, определяющие удлинение теломер в опухолевых клетках Gaspar et al., Gene, 2018, doi: 10.3390/genes9050241 Ets site
- 43. Возможные стратегии антителомеразной терапии Прямое ингибирование теломеразы (ингибирование TR и TERT). Формирование G-квадруплексов на концах теломер
- 44. Неограниченное деление стволовых клеток опухолей Стимуляция деления Иммортализация TERT Bmi1 p16Ink4a ? Myтации протоонкогенов, опухолевых супрессоров
- 45. Нестабильность генома (мутации и эпигенетические изменения) Нечувствительность к рост-ингибирующим сигналам (приобретение бессмертия и др.) Постоянная инициация
- 46. Способы программируемой гибели клеток: Апоптоз 2. Митотическая катастрофа 3. Аутофагия 4. Некроптоз 5. Энтоз
- 48. Скачать презентацию
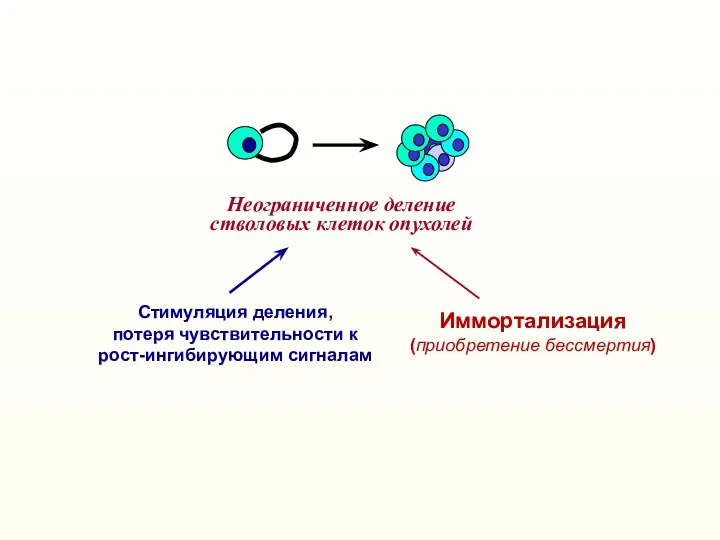
Неограниченное деление
стволовых клеток опухолей
Стимуляция деления,
потеря чувствительности к
рост-ингибирующим сигналам
Иммортализация
(приобретение бессмертия)
Неограниченное деление
стволовых клеток опухолей
Стимуляция деления,
потеря чувствительности к
рост-ингибирующим сигналам
Иммортализация
(приобретение бессмертия)
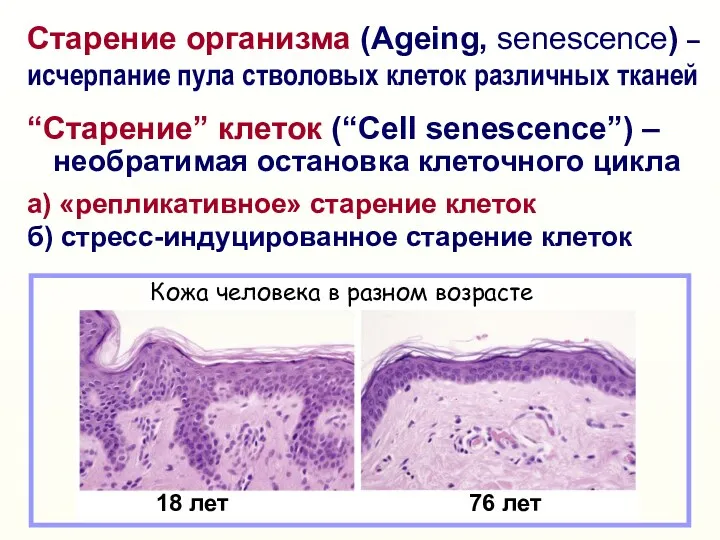
Старение организма (Ageing, senescence) – исчерпание пула стволовых клеток различных тканей
“Старение”
Старение организма (Ageing, senescence) – исчерпание пула стволовых клеток различных тканей
“Старение”

Иммортализация – отмена репликативного
клеточного старения
Нормальные клетки могут быть иммортальны
Иммортализация – отмена репликативного
клеточного старения
Нормальные клетки могут быть иммортальны
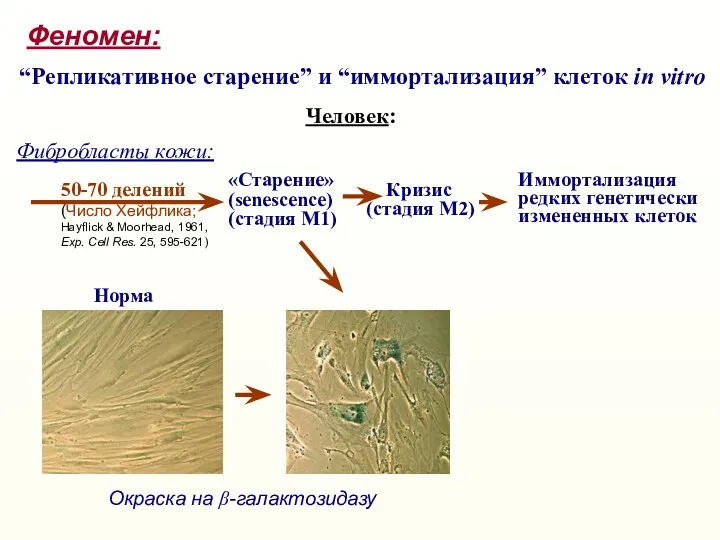
Феномен:
“Репликативное старение” и “иммортализация” клеток in vitro
Человек:
50-70 делений
(Число Хейфлика;
Hayflick &
Феномен:
“Репликативное старение” и “иммортализация” клеток in vitro
Человек:
50-70 делений
(Число Хейфлика;
Hayflick &
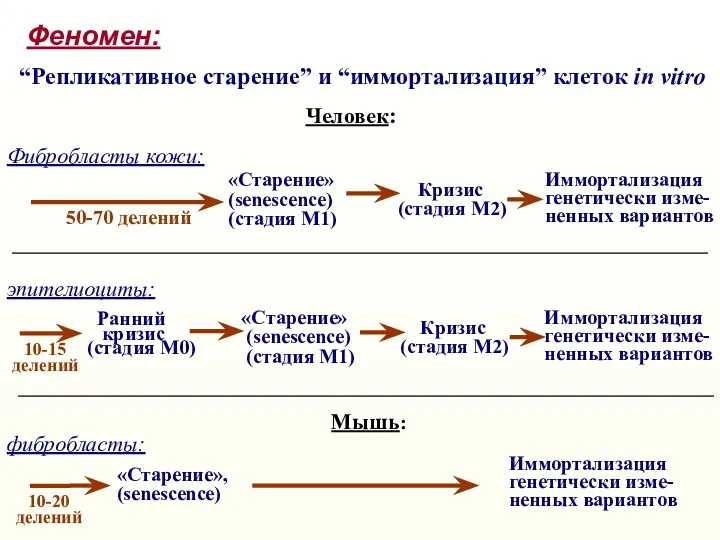
Феномен:
“Репликативное старение” и “иммортализация” клеток in vitro
Человек:
50-70 делений
Фибробласты кожи:
Иммортализация
генетически изме-
ненных
Феномен:
“Репликативное старение” и “иммортализация” клеток in vitro
Человек:
50-70 делений
Фибробласты кожи:
Иммортализация
генетически изме-
ненных

Иммортализация – отмена репликативного
клеточного старения
Опухолевые клетки иммортальны – перевиваемые
Иммортализация – отмена репликативного
клеточного старения
Опухолевые клетки иммортальны – перевиваемые
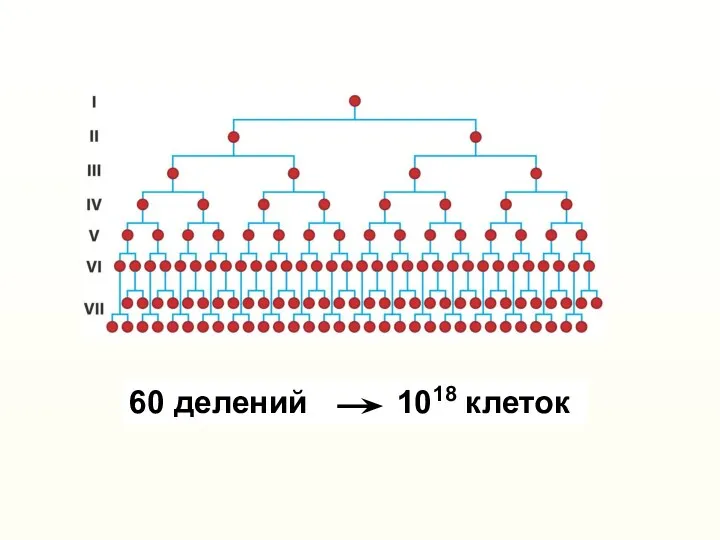
60 делений 1018 клеток
60 делений 1018 клеток
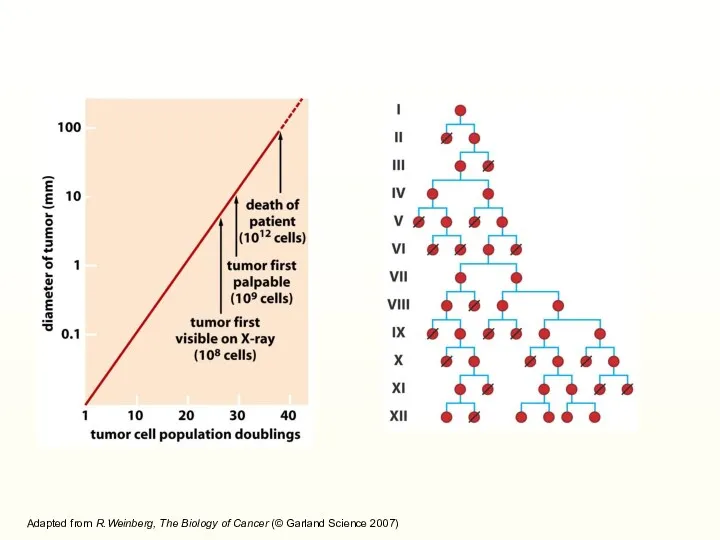
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)

Наиболее общие признаки
репликативного старения клеток
Необратимый арест пролиферации (не может быть
Наиболее общие признаки
репликативного старения клеток
Необратимый арест пролиферации (не может быть
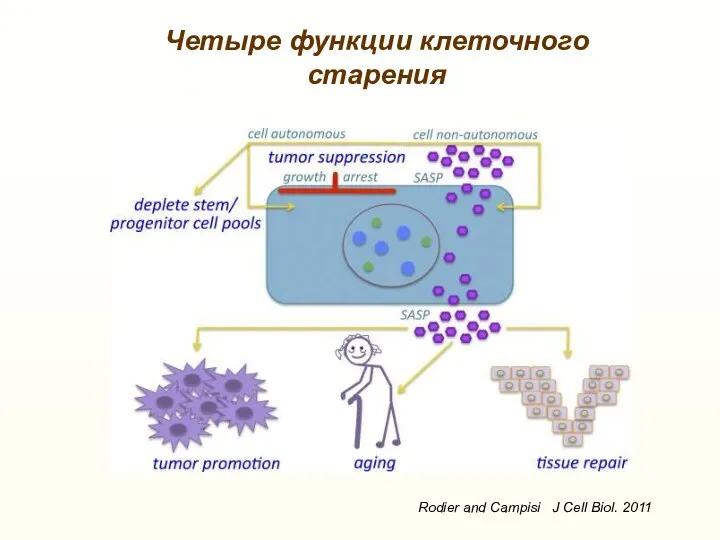
Четыре функции клеточного старения
Rodier and Campisi J Cell Biol. 2011
Четыре функции клеточного старения
Rodier and Campisi J Cell Biol. 2011
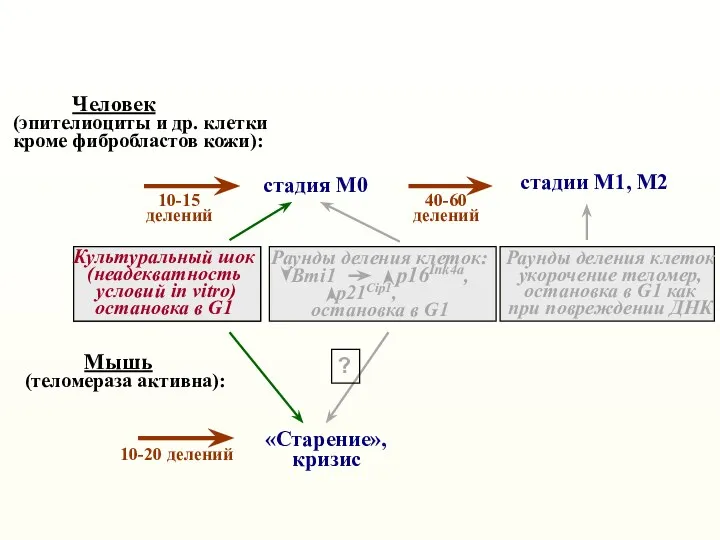
стадия М0
стадии М1, М2
Человек
(эпителиоциты и др. клетки
кроме фибробластов кожи):
«Старение»,
кризис
стадия М0
стадии М1, М2
Человек
(эпителиоциты и др. клетки
кроме фибробластов кожи):
«Старение»,
кризис
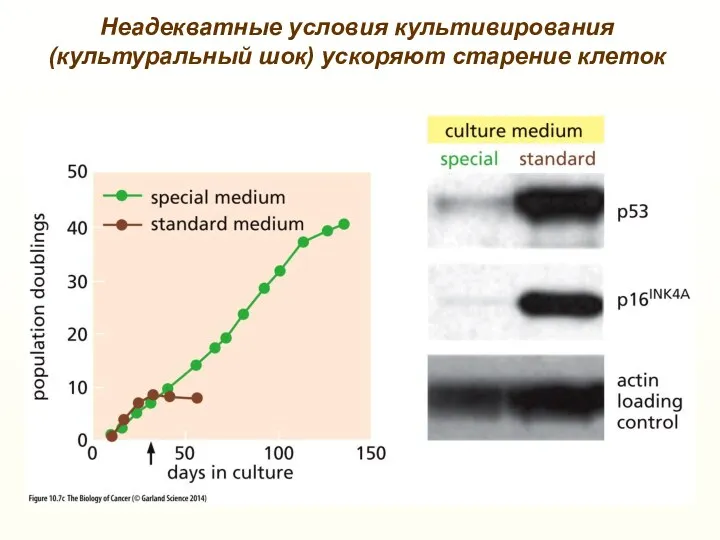
Неадекватные условия культивирования
(культуральный шок) ускоряют старение клеток
Неадекватные условия культивирования
(культуральный шок) ускоряют старение клеток
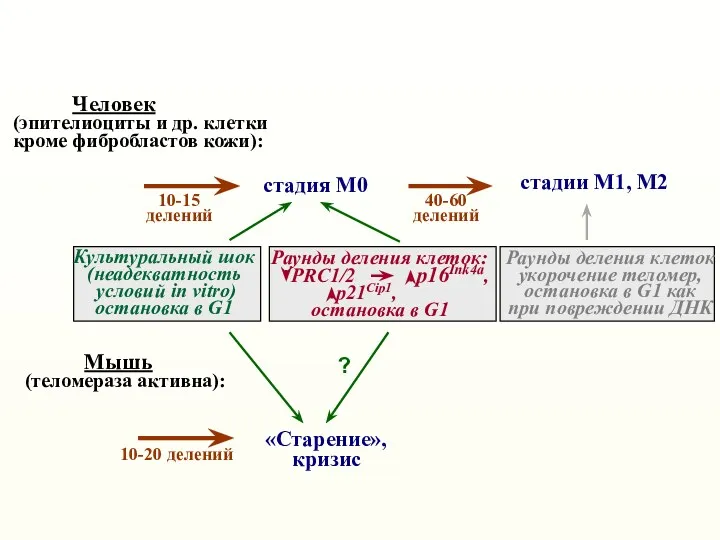
стадия М0
стадии М1, М2
Человек
(эпителиоциты и др. клетки
кроме фибробластов кожи):
«Старение»,
кризис
стадия М0
стадии М1, М2
Человек
(эпителиоциты и др. клетки
кроме фибробластов кожи):
«Старение»,
кризис
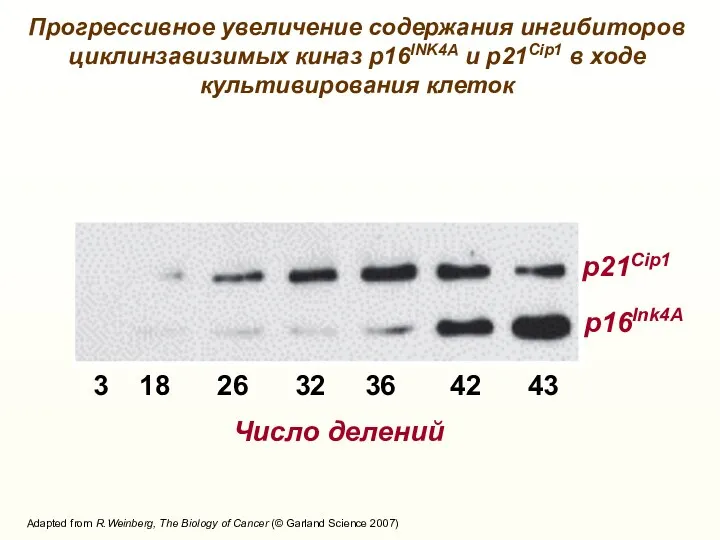
Число делений
3 18 26 32 36 42 43
p21Cip1
p16Ink4А
Adapted from R.Weinberg,
Число делений
3 18 26 32 36 42 43
p21Cip1
p16Ink4А
Adapted from R.Weinberg,
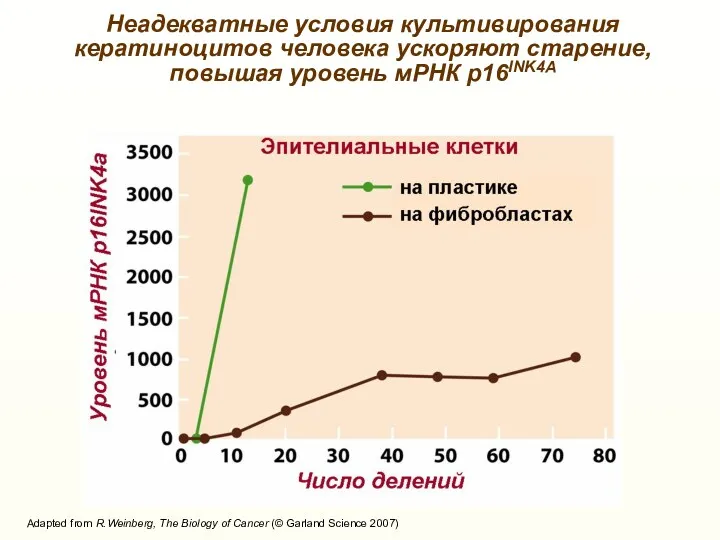
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
Неадекватные
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
Неадекватные

Роль р16Ink4a в «старении» гемопоэтических
cтволовых клеток (ГСК) in vivo
1. В популяции
Роль р16Ink4a в «старении» гемопоэтических
cтволовых клеток (ГСК) in vivo
1. В популяции

Репликативное старение стволовых и
недифференцированных клеток in vivo
Ключевой механизм – увеличение содержания
Репликативное старение стволовых и
недифференцированных клеток in vivo
Ключевой механизм – увеличение содержания
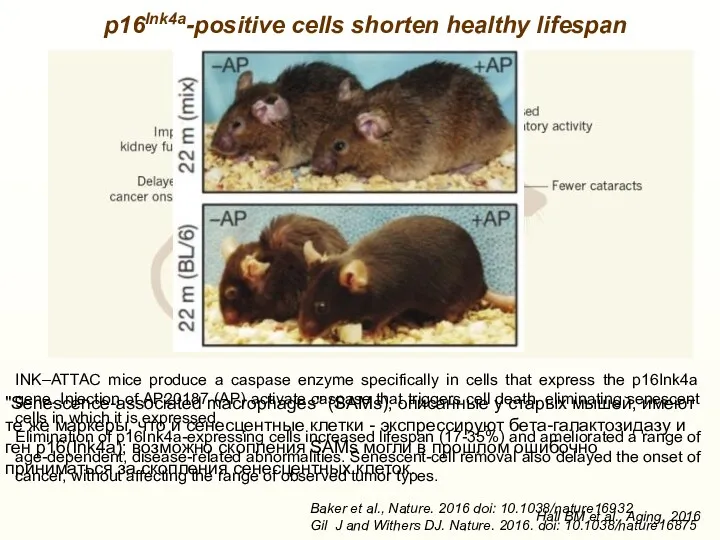
p16Ink4a-positive cells shorten healthy lifespan
Baker et al., Nature. 2016 doi: 10.1038/nature16932
Gil
p16Ink4a-positive cells shorten healthy lifespan
Baker et al., Nature. 2016 doi: 10.1038/nature16932
Gil
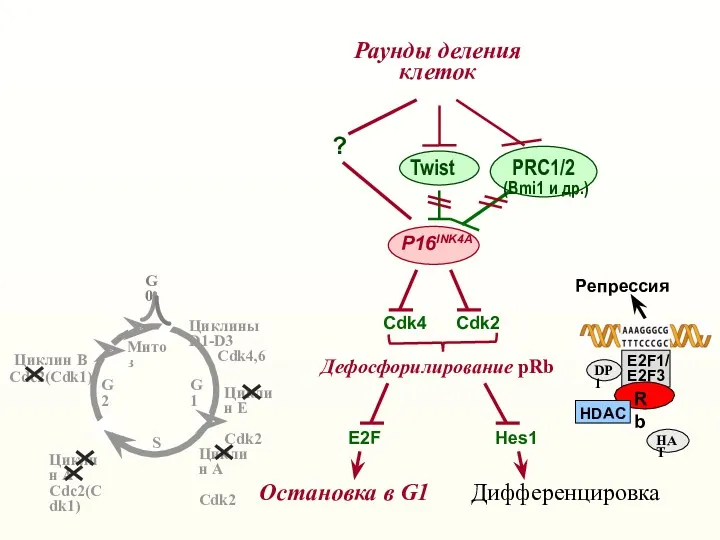
Раунды деления
клеток
Cdk4
Cdk2
Остановка в G1
P16INK4A
PRC1/2
(Bmi1 и др.)
?
Дефосфорилирование pRb
E2F
Hes1
Дифференцировка
Twist
Раунды деления
клеток
Cdk4
Cdk2
Остановка в G1
P16INK4A
PRC1/2
(Bmi1 и др.)
?
Дефосфорилирование pRb
E2F
Hes1
Дифференцировка
Twist
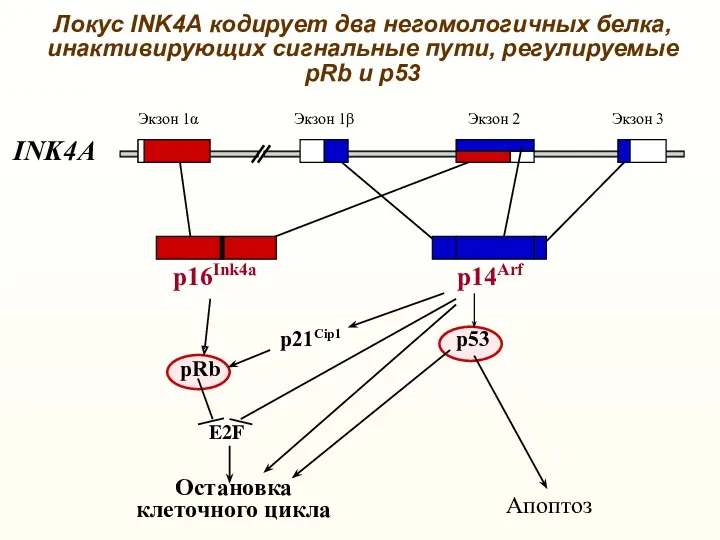
Локус INK4А кодирует два негомологичных белка, инактивирующих сигнальные пути, регулируемые pRb
Локус INK4А кодирует два негомологичных белка, инактивирующих сигнальные пути, регулируемые pRb
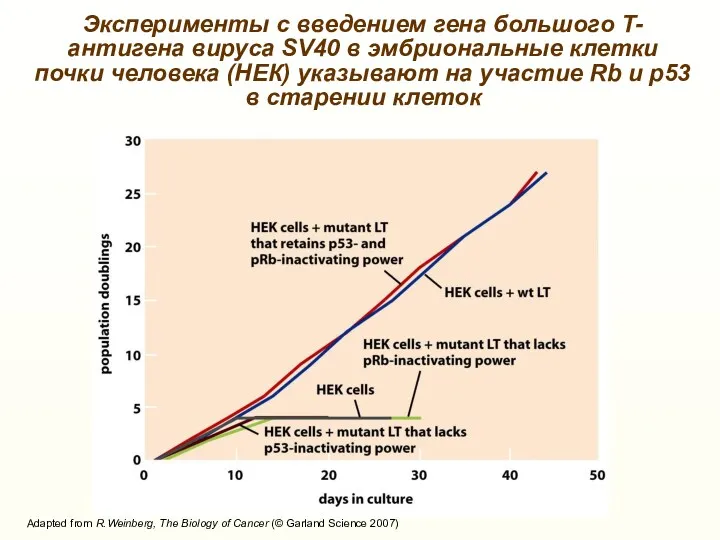
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
Эксперименты
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
Эксперименты
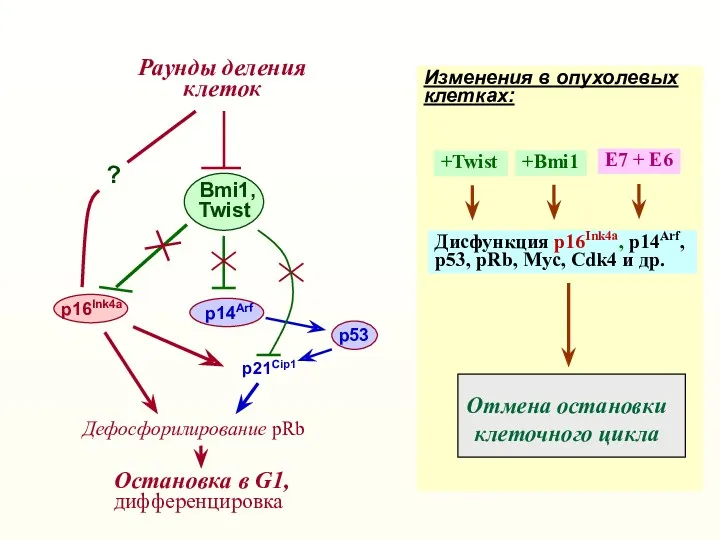
Раунды деления
клеток
p21Cip1
Остановка в G1,
дифференцировка
p16Ink4a
Bmi1,
Twist
p53
p14Arf
Отмена остановки
клеточного цикла
Е7 + Е6
Дисфункция p16Ink4a, p14Arf,
Раунды деления
клеток
p21Cip1
Остановка в G1,
дифференцировка
p16Ink4a
Bmi1,
Twist
p53
p14Arf
Отмена остановки
клеточного цикла
Е7 + Е6
Дисфункция p16Ink4a, p14Arf,

Другие функции Bmi1
Компонент PRC1 комплекса, участвующего в ремоделировании структуры хроматина и
Другие функции Bmi1
Компонент PRC1 комплекса, участвующего в ремоделировании структуры хроматина и
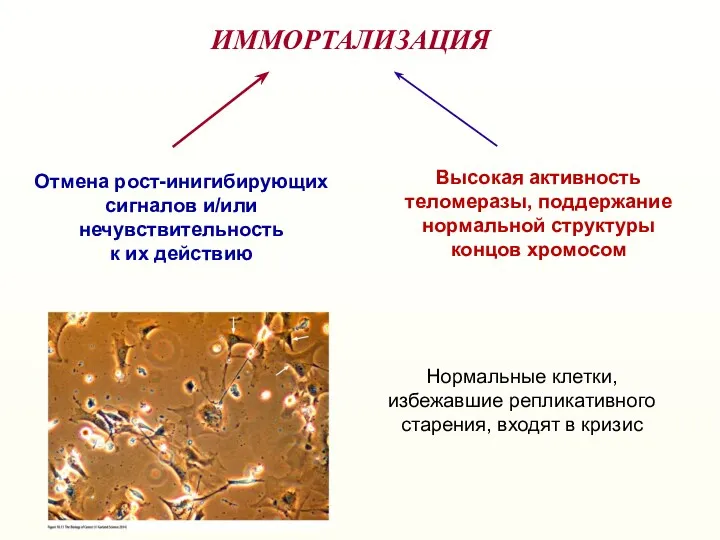
ИММОРТАЛИЗАЦИЯ
Отмена рост-инигибирующих
сигналов и/или
нечувствительность
к их действию
Высокая активность теломеразы, поддержание
нормальной структуры
ИММОРТАЛИЗАЦИЯ
Отмена рост-инигибирующих
сигналов и/или
нечувствительность
к их действию
Высокая активность теломеразы, поддержание
нормальной структуры
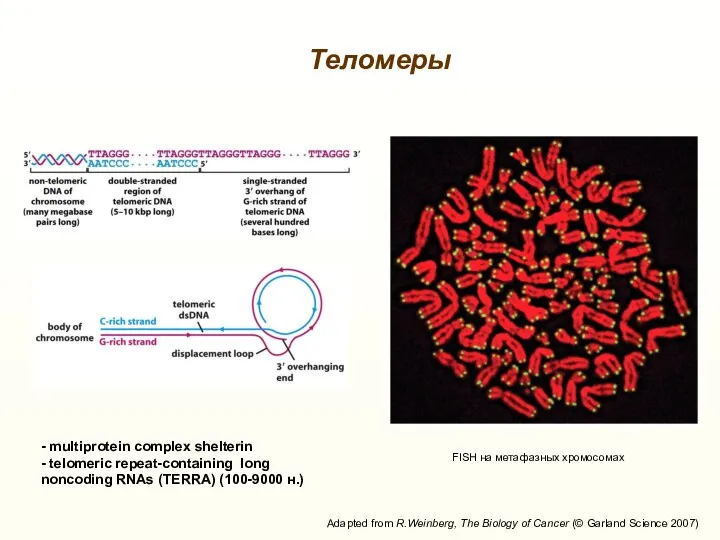
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
FISH
Adapted from R.Weinberg, The Biology of Cancer (© Garland Science 2007)
FISH
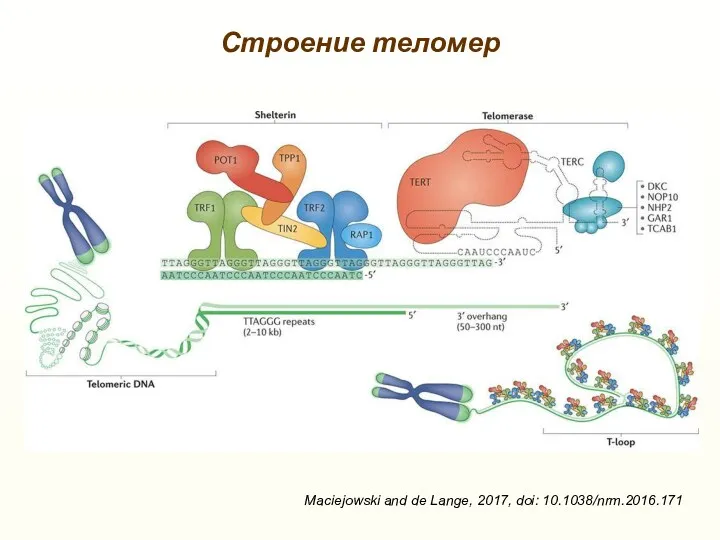
Строение теломер
Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
Строение теломер
Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
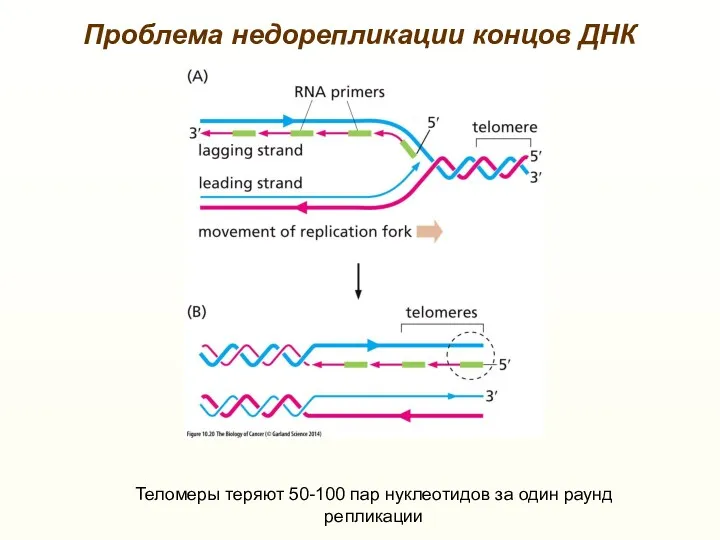
Проблема недорепликации концов ДНК
Теломеры теряют 50-100 пар нуклеотидов за один раунд
Проблема недорепликации концов ДНК
Теломеры теряют 50-100 пар нуклеотидов за один раунд
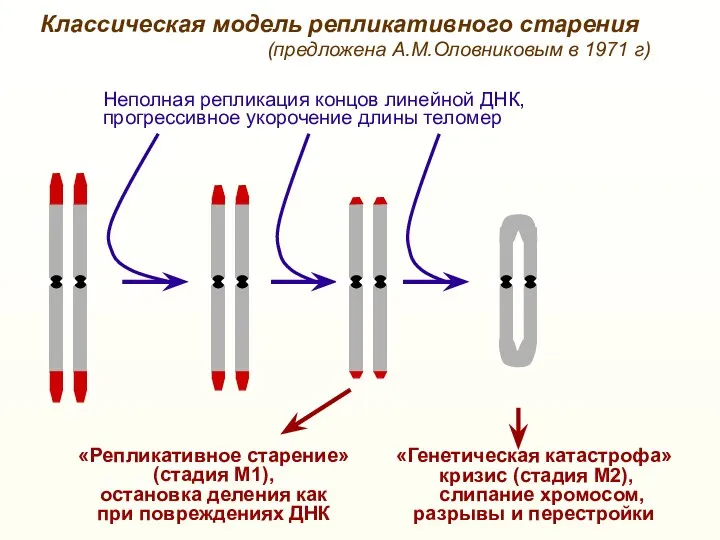
«Репликативное старение»
(стадия М1),
остановка деления как
при повреждениях ДНК
«Генетическая катастрофа»
«Репликативное старение»
(стадия М1),
остановка деления как
при повреждениях ДНК
«Генетическая катастрофа»
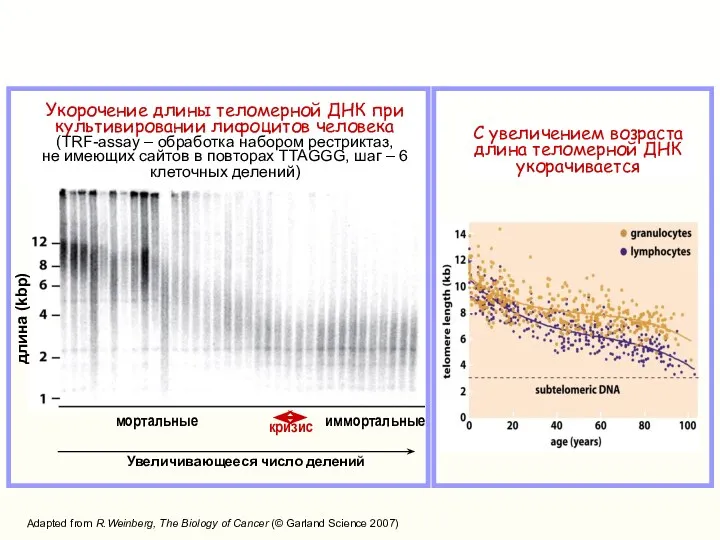
кризис
мортальные
иммортальные
Увеличивающееся число делений
длина (kbp)
Укорочение длины теломерной ДНК при культивировании лифоцитов человека
(TRF-assay
кризис
мортальные
иммортальные
Увеличивающееся число делений
длина (kbp)
Укорочение длины теломерной ДНК при культивировании лифоцитов человека
(TRF-assay
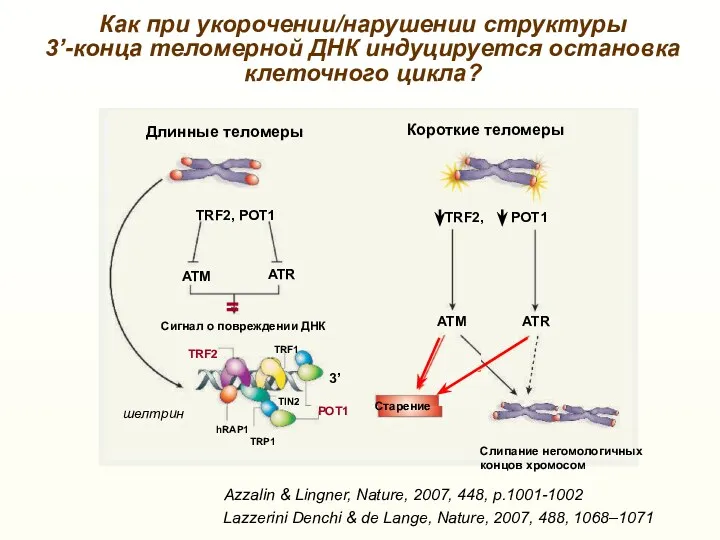
Как при укорочении/нарушении структуры
3’-конца теломерной ДНК индуцируется остановка клеточного цикла?
Как при укорочении/нарушении структуры
3’-конца теломерной ДНК индуцируется остановка клеточного цикла?
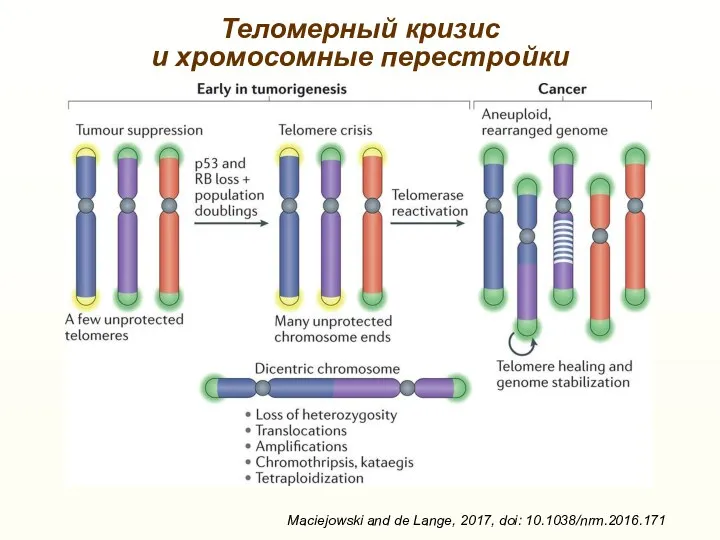
Теломерный кризис
и хромосомные перестройки
Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
Теломерный кризис
и хромосомные перестройки
Maciejowski and de Lange, 2017, doi: 10.1038/nrm.2016.171
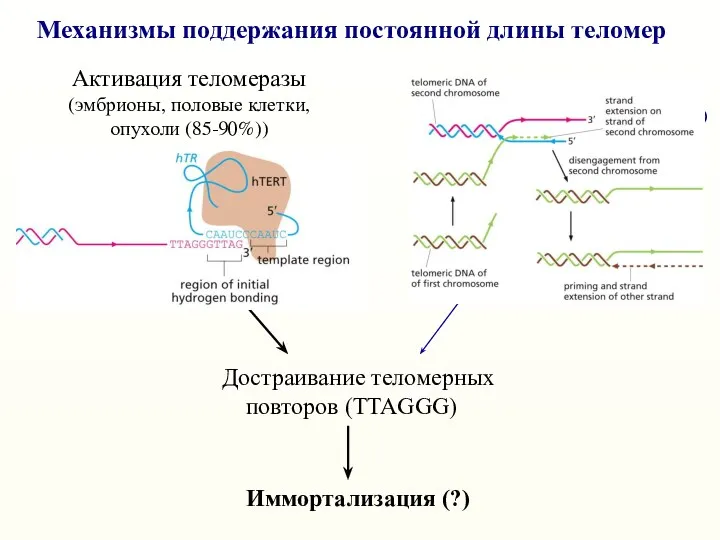
Механизмы поддержания постоянной длины теломер
Активация теломеразы
(эмбрионы, половые клетки,
опухоли (85-90%))
Механизмы поддержания постоянной длины теломер
Активация теломеразы
(эмбрионы, половые клетки,
опухоли (85-90%))
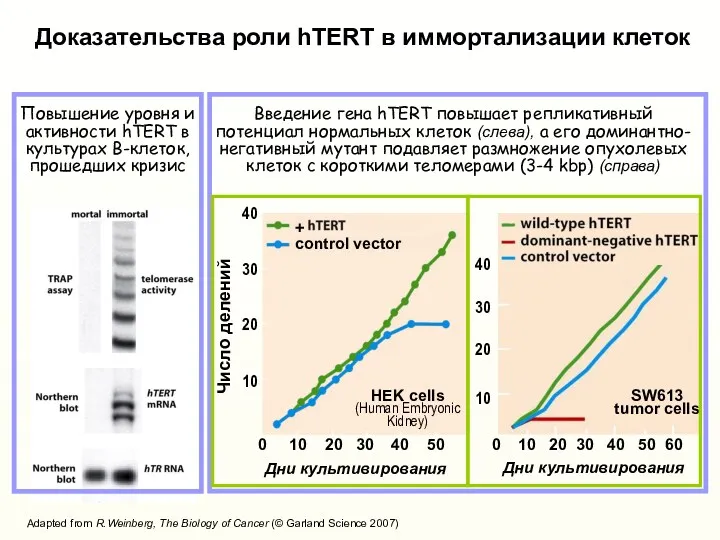
Введение гена hTERT повышает репликативный потенциал нормальных клеток (слева), а его
Введение гена hTERT повышает репликативный потенциал нормальных клеток (слева), а его
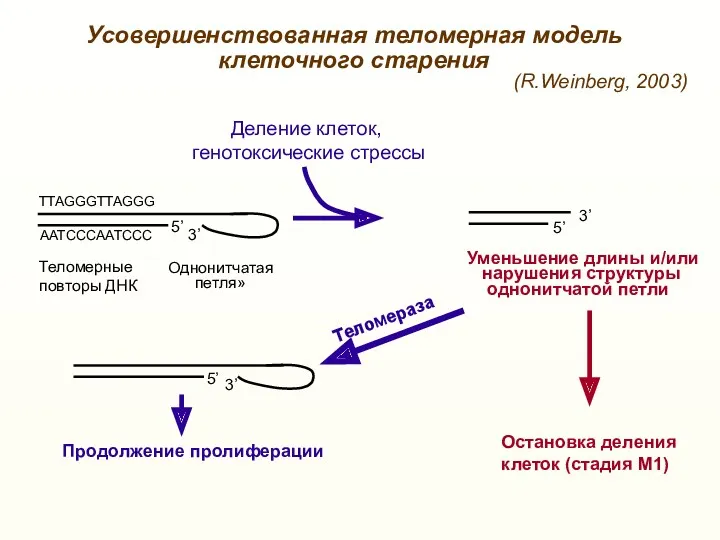
Усовершенствованная теломерная модель
клеточного старения
Теломерные
повторы ДНК
Однонитчатая
петля»
ТTAGGGTTAGGG
5’
3’
AATCCCAATCCC
5’
3’
Деление клеток,
генотоксические стрессы
Остановка деления
клеток (стадия
Усовершенствованная теломерная модель
клеточного старения
Теломерные
повторы ДНК
Однонитчатая
петля»
ТTAGGGTTAGGG
5’
3’
AATCCCAATCCC
5’
3’
Деление клеток,
генотоксические стрессы
Остановка деления
клеток (стадия
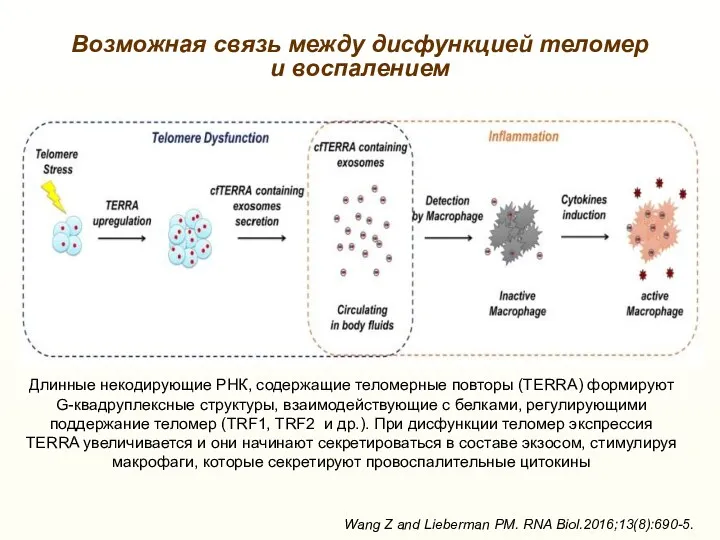
Возможная связь между дисфункцией теломер и воспалением
Wang Z and Lieberman PM.
Возможная связь между дисфункцией теломер и воспалением
Wang Z and Lieberman PM.
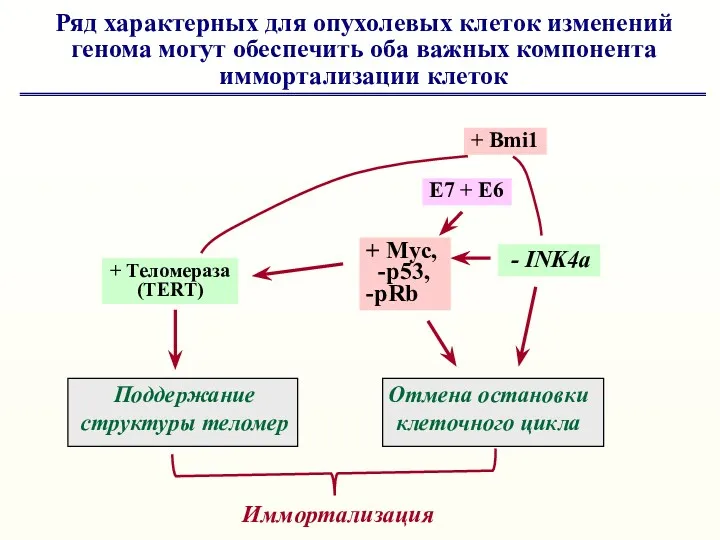
+ Теломераза
(TERT)
Поддержание
структуры теломер
Отмена остановки
клеточного цикла
- INK4a
Е7 + Е6
+ Myc,
+ Теломераза
(TERT)
Поддержание
структуры теломер
Отмена остановки
клеточного цикла
- INK4a
Е7 + Е6
+ Myc,
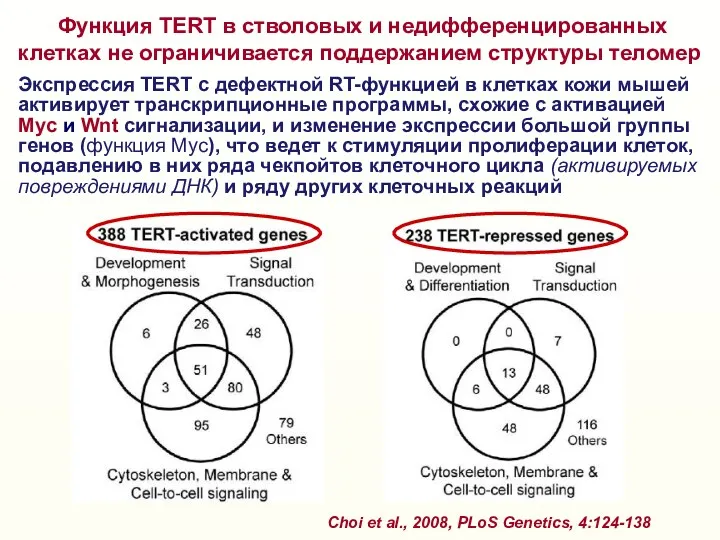
Функция TERT в стволовых и недифференцированных
клетках не ограничивается поддержанием структуры
Функция TERT в стволовых и недифференцированных
клетках не ограничивается поддержанием структуры
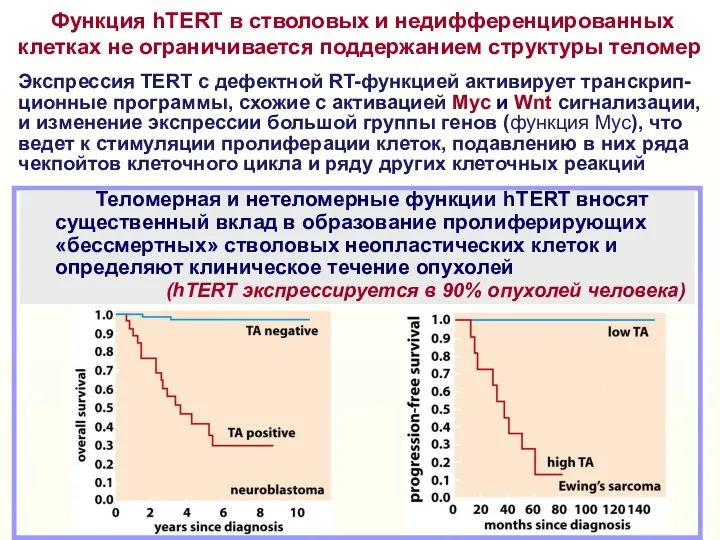
Функция hTERT в стволовых и недифференцированных
клетках не ограничивается поддержанием структуры
Функция hTERT в стволовых и недифференцированных
клетках не ограничивается поддержанием структуры

Активация сигнальных путей факторов роста, Wnt, NF-κB
Усиление стволовых характеристик клетки
Улучшение рибосомного
Активация сигнальных путей факторов роста, Wnt, NF-κB
Усиление стволовых характеристик клетки
Улучшение рибосомного
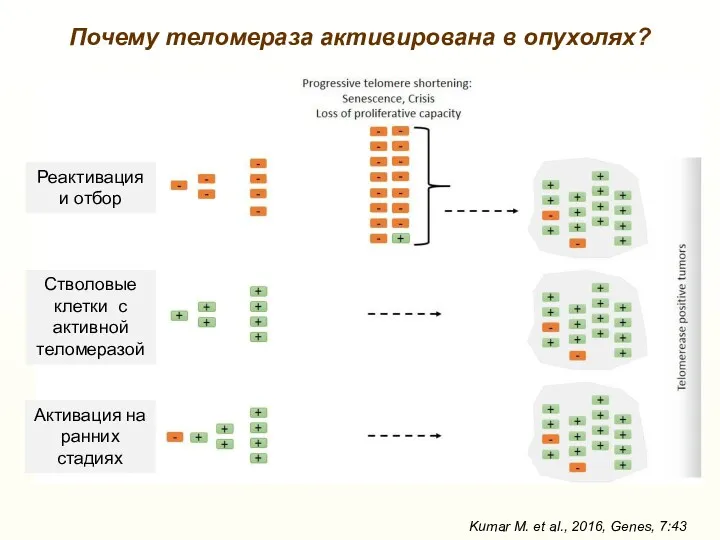
Почему теломераза активирована в опухолях?
Kumar M. et al., 2016, Genes, 7:43
Почему теломераза активирована в опухолях?
Kumar M. et al., 2016, Genes, 7:43
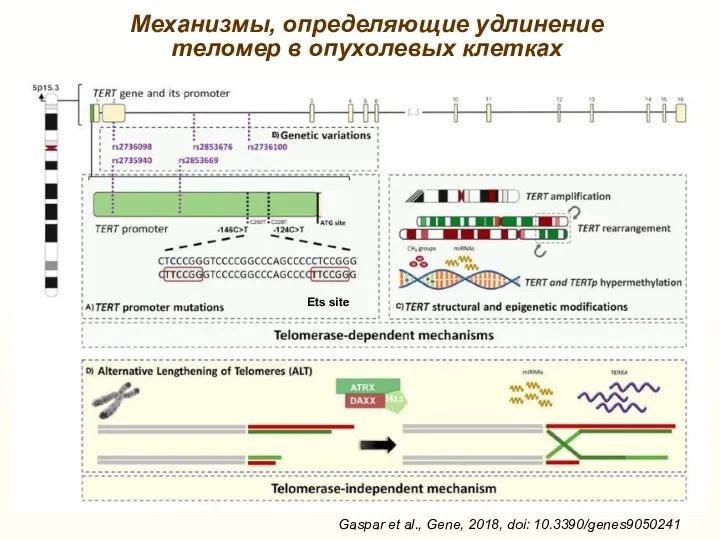
Механизмы, определяющие удлинение теломер в опухолевых клетках
Gaspar et al., Gene, 2018,
Механизмы, определяющие удлинение теломер в опухолевых клетках
Gaspar et al., Gene, 2018,
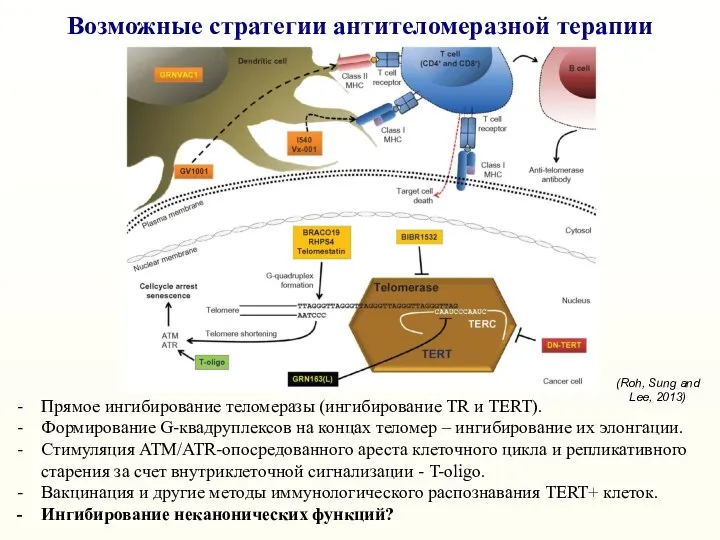
Возможные стратегии антителомеразной терапии
Прямое ингибирование теломеразы (ингибирование TR и TERT).
Формирование G-квадруплексов
Возможные стратегии антителомеразной терапии
Прямое ингибирование теломеразы (ингибирование TR и TERT).
Формирование G-квадруплексов
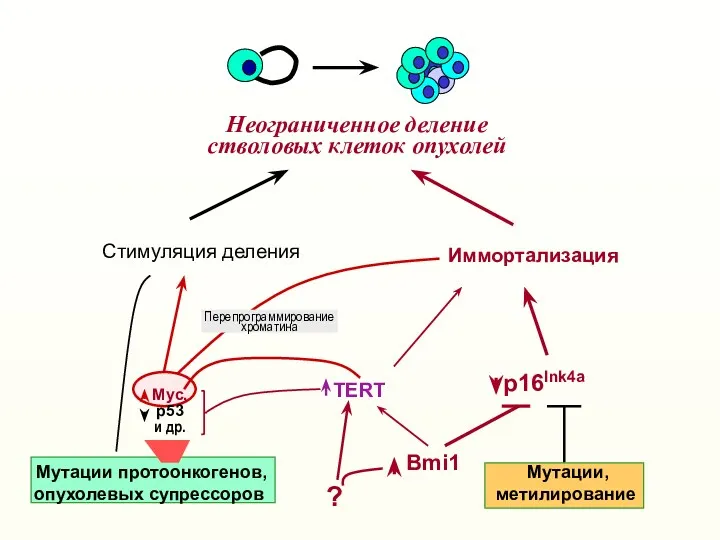
Неограниченное деление
стволовых клеток опухолей
Стимуляция деления
Иммортализация
TERT
Bmi1
p16Ink4a
?
Myтации протоонкогенов,
опухолевых супрессоров
Myтации,
метилирование
Перепрограммирование
хроматина
Myc, р53
и
Неограниченное деление
стволовых клеток опухолей
Стимуляция деления
Иммортализация
TERT
Bmi1
p16Ink4a
?
Myтации протоонкогенов,
опухолевых супрессоров
Myтации,
метилирование
Перепрограммирование
хроматина
Myc, р53
и
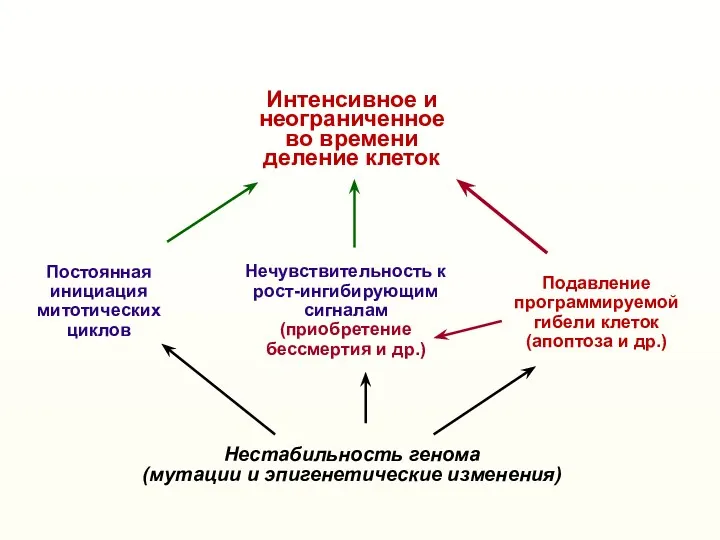
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация

Способы программируемой гибели клеток:
Апоптоз
2. Митотическая катастрофа
3. Аутофагия
4. Некроптоз
5. Энтоз
Способы программируемой гибели клеток:
Апоптоз
2. Митотическая катастрофа
3. Аутофагия
4. Некроптоз
5. Энтоз
 Рахіт: причини, фактори, лікування
Рахіт: причини, фактори, лікування Сосудистый тонус и его компоненты
Сосудистый тонус и его компоненты Эпидемиология наркоманий и токсикоманий. Соматические нарушения при болезнях зависимости
Эпидемиология наркоманий и токсикоманий. Соматические нарушения при болезнях зависимости Микола Михайлович Амосов
Микола Михайлович Амосов Методы обследования беременной в женской консультации
Методы обследования беременной в женской консультации Плеврит. Основные формы плевритов
Плеврит. Основные формы плевритов Экстрапирамидалық жүйе жане мишық зақымдану синдромы
Экстрапирамидалық жүйе жане мишық зақымдану синдромы Клинико-психологическая характеристика акалькулии и дискалькулии детского возраста
Клинико-психологическая характеристика акалькулии и дискалькулии детского возраста Тұлғалық сұрақнама, медицинадағы соматикалық науқастардың психофизикалық қалпын клиникалық стандартын бағалау шкаласы
Тұлғалық сұрақнама, медицинадағы соматикалық науқастардың психофизикалық қалпын клиникалық стандартын бағалау шкаласы Методы физиотерапии в реабилитации онкологических больных
Методы физиотерапии в реабилитации онкологических больных Вирусные гепатиты
Вирусные гепатиты Топографическая анатомия нижней конечности. Области мышечной и сосудистой лакуны, бедра, подколенная ямка, ягодичная область
Топографическая анатомия нижней конечности. Области мышечной и сосудистой лакуны, бедра, подколенная ямка, ягодичная область Экстракорпоральные методы коррекции волемического статуса у пациентов в критических состояниях
Экстракорпоральные методы коррекции волемического статуса у пациентов в критических состояниях Имплантаты в стоматологии
Имплантаты в стоматологии Избранные вопросы интенсивной терапии неотложных состояний у детей
Избранные вопросы интенсивной терапии неотложных состояний у детей Психоорганический синдром и когнитивные нарушения – взгляд психиатра
Психоорганический синдром и когнитивные нарушения – взгляд психиатра Доғалы протез
Доғалы протез Тромбопрофилактика у пациентов с ФП как путь снижения смертности от неинфекционных заболеваний
Тромбопрофилактика у пациентов с ФП как путь снижения смертности от неинфекционных заболеваний Шок. Общие патофизиологические аспекты шока. Контрольные вопросы
Шок. Общие патофизиологические аспекты шока. Контрольные вопросы Балалардағы ауыз қуысы шырышты қабатының аурулары
Балалардағы ауыз қуысы шырышты қабатының аурулары Папулосквамозні дерматози
Папулосквамозні дерматози СПИД – последняя, смертельная стадия ВИЧ-инфекции. Часть 2
СПИД – последняя, смертельная стадия ВИЧ-инфекции. Часть 2 Гемофильная инфекция
Гемофильная инфекция Синдром поликистозных яичников
Синдром поликистозных яичников ҚР дәрілік заттарға сараптаманы жүргізу ережесі мен тәртібі
ҚР дәрілік заттарға сараптаманы жүргізу ережесі мен тәртібі Ведение физиологических родов
Ведение физиологических родов Аутоиммунный гепатит. Перекрест с первичным биллиарным холангитом
Аутоиммунный гепатит. Перекрест с первичным биллиарным холангитом Болезни оперированного желудка. Демпинг-синдром
Болезни оперированного желудка. Демпинг-синдром