- Противоопухолевой иммунитет. Теории канцерогенеза

Содержание
- 2. План Противоопухолевой иммунитет. Введение 1.1 Опухолевые антигены 1.2 Трансформация нормальных клеток в опухолевые 1.3 Проонкогены 1.4
- 3. ПРОТИВООПУХОЛЕВОЙ ИММУНИТЕТ.
- 4. Введение Онкопатология занимает второе место после сердечно-сосудистых заболеваний среди причин смерти в цивилизованных странах. Опухоли (новообразования)
- 5. Главной отличительной особенностью опухолевых клеток от нормальных является способность к неограниченному росту и потеря механизмов аутокринной
- 6. Опухолевые антигены Иммунологические исследования показали, что на опухолевых клетках могут быть представлены два вида антигенов: опухолеспецифические
- 7. Некоторые опухолеспецифические антигены, экспрессируемые при онкопатологии
- 8. Некоторые раково-эмбриональные антигены, выявляемые при онкопатологии ! Исследования показали, что гены, кодирующие опухолеспецифические антигены, часто отличаются
- 9. Опухолевые антигены Специфические антигены вирусиндуцированных опухолей, как правило, представлены вирусными пептидами. Это подтверждается множеством данных. Так,
- 10. Альфа-фетопротеин Альфа-фетопротеин впервые был обнаружен в фетальной сыворотке теленка. Он вырабатывается преимущественно гепатоцитами, а также клетками
- 11. РЭА Раково-эмбриональный антиген является мембранным гликопротеином. Экспрессируется в норме на гастроинтестинальных клетках и клетках печени плода
- 12. Трансформация нормальных клеток в опухолевые В основе злокачественной трансформации клеток лежат изменения в их геноме. К
- 13. Проонкогены Известные к настоящему времени проонкогены и кодируемые ими молекулы. По своим функциональным особенностям они могут
- 14. Гены, индуцирующие клеточную пролиферацию Первая категория проонкогенов и их онкогенных копий кодирует белки, которые функционируют как
- 15. Гены, кодирующие факторы, способствующие клеточной пролиферации
- 16. Гены, подавляющие клеточную пролиферацию, опухолесупрессирующие гены Вторая категория проонкогенов – антионкогены кодируют белки, подавляющие клеточный рост.
- 17. Гены, кодирующие продукты, подавляющие клеточную пролиферацию
- 18. Гены апоптоза Третья категория проонкогенов кодирует белки, регулирующие запрограммированную клеточную смерть (подавляющие или индуцирующие апоптоз). В
- 19. Также онкогены были выявлены у ряда вирусов. Так, у вируса саркомы Роус был идентифицирован онкоген, названный
- 20. Иммунные реакции на опухоли Эксперименты на животных показали, что организм на опухолевые антигены способен развивать как
- 21. НК – Клетки В отличие от Т-цитотоксических лимфоцитов, распознавание НК-клетками опухолевых антигенов не рестриктировано антигенами HLA.
- 22. Макрофаги свое цитотоксическое действие на опухоли оказывают через развитие реакции АЗКЦ, либо в процессе прямого контакта
- 23. Гуморальный иммунитет Гуморальные иммунные реакции при опухолевом росте могут как способствовать регрессии опухоли, так и стимулировать
- 24. Механизмы уклонения опухоли от иммунной защиты организма Ежегодно от рака умирает большое количество людей, что свидетельствует
- 25. 1. Гипотеза иммунного усиления Согласно гипотезе, появляющиеся в ходе развития опухоли антиопухолевые антитела экранируют опухолевые детерминанты,
- 26. Иммунодиагностика рака В настоящее время иммунодиагностика опухолей основана на выявлении в сыворотке и жидкостях больного опухолеспецифических
- 27. ТЕОРИИ КАНЦЕРОГЕНЕЗА
- 28. Введение Теоретические предположения о природе опухолей высказывались давно, но гипотезы, на основании которых могли выполняться научные
- 30. Мутационная теория происхождения рака Согласно этой теории рак в организме человека возникает вследствие накопления мутаций в
- 31. Теория случайных мутаций Автор — Лоренс Леб (учёный из Вашингтонского университета). Генетики утверждают,что в любой клетке
- 32. Теория ранней хромосомной нестабильности Основоположники - Кристоф Лингаур и Берт Фогельштейн. В 1997 г. они обнаружили,
- 33. Теория анеуплоидии Автор — Питер Дюсберг (учёный из Калифорнийского университета в Беркли) создал теорию, согласно которой
- 34. Теория эмбриональных клеток В разные годы разные учёные выдвигали гипотезы насчёт развития рака из эмбриональных клеток.
- 35. Тканевая теория онкогенеза Одним из авторов тканевой теории онкогенеза можно по по праву назвать Ю.М. Васильева.
- 36. Теория Выпадения при малигнизации из клетки исчезают белки, необходимые для регулирования клеточной жизнедеятельности. При действии вирусов,
- 37. Вывод Общепринятой теории этиологии и патогенеза опухолей пока не существует. Сторонники полиэтиологических представлений считают, что каждый
- 39. Скачать презентацию

План
Противоопухолевой иммунитет.
Введение
1.1 Опухолевые антигены
1.2 Трансформация нормальных клеток в опухолевые
1.3
План
Противоопухолевой иммунитет.
Введение
1.1 Опухолевые антигены
1.2 Трансформация нормальных клеток в опухолевые
1.3

ПРОТИВООПУХОЛЕВОЙ ИММУНИТЕТ.
ПРОТИВООПУХОЛЕВОЙ ИММУНИТЕТ.
Введение
Онкопатология занимает второе место после сердечно-сосудистых заболеваний среди причин смерти
Введение
Онкопатология занимает второе место после сердечно-сосудистых заболеваний среди причин смерти

Главной отличительной особенностью опухолевых клеток от нормальных является способность к
Главной отличительной особенностью опухолевых клеток от нормальных является способность к

Опухолевые антигены
Иммунологические исследования показали, что на опухолевых клетках могут быть
Опухолевые антигены
Иммунологические исследования показали, что на опухолевых клетках могут быть
Некоторые опухолеспецифические антигены, экспрессируемые при онкопатологии
Некоторые опухолеспецифические антигены, экспрессируемые при онкопатологии

Некоторые раково-эмбриональные антигены, выявляемые при онкопатологии
! Исследования показали, что гены, кодирующие
Некоторые раково-эмбриональные антигены, выявляемые при онкопатологии
! Исследования показали, что гены, кодирующие

Опухолевые антигены
Специфические антигены вирусиндуцированных опухолей, как правило, представлены вирусными
Опухолевые антигены
Специфические антигены вирусиндуцированных опухолей, как правило, представлены вирусными
Альфа-фетопротеин
Альфа-фетопротеин впервые был обнаружен в фетальной сыворотке теленка. Он
Альфа-фетопротеин
Альфа-фетопротеин впервые был обнаружен в фетальной сыворотке теленка. Он
РЭА
Раково-эмбриональный антиген является мембранным гликопротеином. Экспрессируется в норме на гастроинтестинальных
РЭА
Раково-эмбриональный антиген является мембранным гликопротеином. Экспрессируется в норме на гастроинтестинальных

Трансформация нормальных клеток в опухолевые
В основе злокачественной трансформации клеток лежат
Трансформация нормальных клеток в опухолевые
В основе злокачественной трансформации клеток лежат

Проонкогены
Известные к настоящему времени проонкогены и кодируемые ими молекулы. По
Проонкогены
Известные к настоящему времени проонкогены и кодируемые ими молекулы. По

Гены, индуцирующие клеточную пролиферацию
Первая категория проонкогенов и их онкогенных копий
Гены, индуцирующие клеточную пролиферацию
Первая категория проонкогенов и их онкогенных копий

Гены, кодирующие факторы, способствующие клеточной пролиферации
Гены, кодирующие факторы, способствующие клеточной пролиферации

Гены, подавляющие клеточную пролиферацию, опухолесупрессирующие гены
Вторая категория проонкогенов – антионкогены кодируют
Гены, подавляющие клеточную пролиферацию, опухолесупрессирующие гены
Вторая категория проонкогенов – антионкогены кодируют

Гены, кодирующие продукты, подавляющие клеточную пролиферацию
Гены, кодирующие продукты, подавляющие клеточную пролиферацию

Гены апоптоза
Третья категория проонкогенов кодирует белки, регулирующие запрограммированную клеточную смерть (подавляющие
Гены апоптоза
Третья категория проонкогенов кодирует белки, регулирующие запрограммированную клеточную смерть (подавляющие

Также онкогены были выявлены у ряда вирусов. Так, у вируса
Также онкогены были выявлены у ряда вирусов. Так, у вируса

Иммунные реакции на опухоли
Эксперименты на животных показали, что организм на
Иммунные реакции на опухоли
Эксперименты на животных показали, что организм на
НК – Клетки
В отличие от Т-цитотоксических лимфоцитов, распознавание НК-клетками
НК – Клетки
В отличие от Т-цитотоксических лимфоцитов, распознавание НК-клетками
Макрофаги
свое цитотоксическое действие на опухоли оказывают через развитие реакции АЗКЦ,
Макрофаги
свое цитотоксическое действие на опухоли оказывают через развитие реакции АЗКЦ,

Гуморальный иммунитет
Гуморальные иммунные реакции при опухолевом росте могут как способствовать
Гуморальный иммунитет
Гуморальные иммунные реакции при опухолевом росте могут как способствовать

Механизмы уклонения опухоли от иммунной защиты организма
Ежегодно от рака умирает
Механизмы уклонения опухоли от иммунной защиты организма
Ежегодно от рака умирает

1. Гипотеза иммунного усиления
Согласно гипотезе, появляющиеся в ходе развития опухоли
1. Гипотеза иммунного усиления
Согласно гипотезе, появляющиеся в ходе развития опухоли

Иммунодиагностика рака
В настоящее время иммунодиагностика опухолей основана на выявлении в сыворотке
Иммунодиагностика рака
В настоящее время иммунодиагностика опухолей основана на выявлении в сыворотке

ТЕОРИИ КАНЦЕРОГЕНЕЗА
ТЕОРИИ КАНЦЕРОГЕНЕЗА

Введение
Теоретические предположения о природе опухолей высказывались давно, но гипотезы, на основании
Введение
Теоретические предположения о природе опухолей высказывались давно, но гипотезы, на основании


Мутационная теория происхождения рака
Согласно этой теории рак в организме человека возникает
Мутационная теория происхождения рака
Согласно этой теории рак в организме человека возникает

Теория случайных мутаций
Автор — Лоренс Леб (учёный из Вашингтонского университета).
Генетики утверждают,что
Теория случайных мутаций
Автор — Лоренс Леб (учёный из Вашингтонского университета).
Генетики утверждают,что

Теория ранней хромосомной нестабильности
Основоположники - Кристоф Лингаур и Берт Фогельштейн. В
Теория ранней хромосомной нестабильности
Основоположники - Кристоф Лингаур и Берт Фогельштейн. В

Теория анеуплоидии
Автор — Питер Дюсберг (учёный из Калифорнийского университета в Беркли)
Теория анеуплоидии
Автор — Питер Дюсберг (учёный из Калифорнийского университета в Беркли)

Теория эмбриональных клеток
В разные годы разные учёные выдвигали гипотезы насчёт развития
Теория эмбриональных клеток
В разные годы разные учёные выдвигали гипотезы насчёт развития

Тканевая теория онкогенеза
Одним из авторов тканевой теории онкогенеза можно по по
Тканевая теория онкогенеза
Одним из авторов тканевой теории онкогенеза можно по по

Теория Выпадения
при малигнизации из клетки исчезают белки, необходимые для регулирования клеточной
Теория Выпадения
при малигнизации из клетки исчезают белки, необходимые для регулирования клеточной

Вывод
Общепринятой теории этиологии и патогенеза опухолей пока не существует. Сторонники
Вывод
Общепринятой теории этиологии и патогенеза опухолей пока не существует. Сторонники
 Желтухи новорождённых
Желтухи новорождённых Methods of examination in gynecology
Methods of examination in gynecology Кодекс Республики Казахстан О здоровье народа и системе здравоохранения. Статьи Кодекса о профилактике инфекционных болезней
Кодекс Республики Казахстан О здоровье народа и системе здравоохранения. Статьи Кодекса о профилактике инфекционных болезней Оказание неотложной помощи детям
Оказание неотложной помощи детям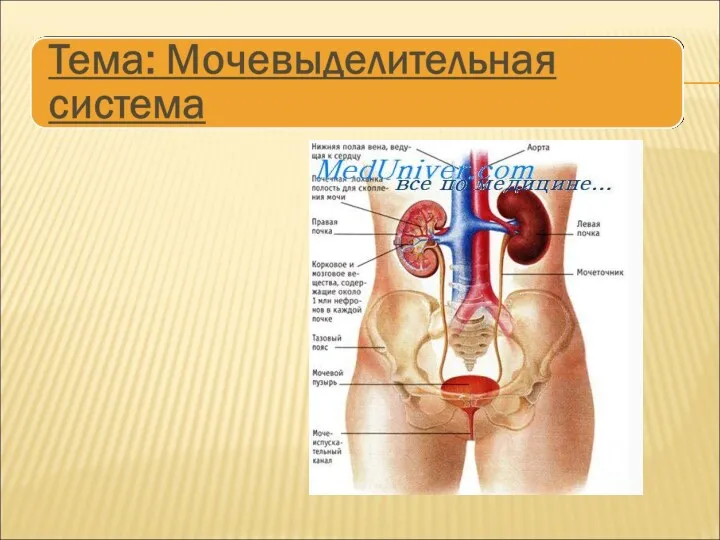 Мочевыделительная система
Мочевыделительная система Инфузионная терапия и парентеральное питание
Инфузионная терапия и парентеральное питание Химические реакции с участием макромолекул
Химические реакции с участием макромолекул Кешенді медициналық ақпараттық жүйе (КМИС)
Кешенді медициналық ақпараттық жүйе (КМИС) Повреждения при выстреле из дробового оружия. Определение дистанции выстрела
Повреждения при выстреле из дробового оружия. Определение дистанции выстрела Вибрациялық аурулар. Профилактикасының принціптері
Вибрациялық аурулар. Профилактикасының принціптері Дезинфекция. Методы дезинфекции
Дезинфекция. Методы дезинфекции Лимфа айналысы. Морфологиялық және функциональдық ерекшеліктері
Лимфа айналысы. Морфологиялық және функциональдық ерекшеліктері Қазақ халық медицинасы
Қазақ халық медицинасы Проблемы наркомании и алкоголизма в России
Проблемы наркомании и алкоголизма в России Общие принципы диагностики и лечения профессиональных заболеваний
Общие принципы диагностики и лечения профессиональных заболеваний Дифференциальная диагностика суставного синдрома
Дифференциальная диагностика суставного синдрома Заболевания ЖКТ
Заболевания ЖКТ Основы конструирования ортодонтических аппаратов
Основы конструирования ортодонтических аппаратов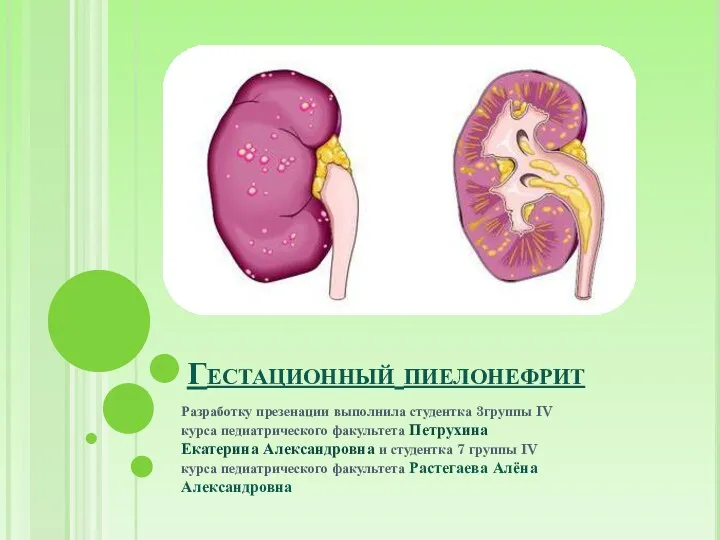 Гестационный пиелонефрит
Гестационный пиелонефрит Сбалансированная инфузионная терапия при тяжелых огнестрельных ранениях
Сбалансированная инфузионная терапия при тяжелых огнестрельных ранениях Классификация, мониторирование, лечение бронхиальной астмы у детей
Классификация, мониторирование, лечение бронхиальной астмы у детей ҰлпАның қабынбалы аурулары
ҰлпАның қабынбалы аурулары Қызыл жалпақ теміреткі. Жүйелі қызыл жегі. Күлдіреуікше. Ауыз қуысындағы көрінісі
Қызыл жалпақ теміреткі. Жүйелі қызыл жегі. Күлдіреуікше. Ауыз қуысындағы көрінісі Аномалии родовой деятельности. Классификация аномалий сократительной деятельности матки
Аномалии родовой деятельности. Классификация аномалий сократительной деятельности матки Предмет и задачи медтоксикологии
Предмет и задачи медтоксикологии Haemolytic disease of the fetus and newborn. Rh isoimmunization
Haemolytic disease of the fetus and newborn. Rh isoimmunization Тромботический синдром
Тромботический синдром Методы лечения аномалий прикуса в периоде смешанных зубах
Методы лечения аномалий прикуса в периоде смешанных зубах