- Экспрессия генов и механизмы регуляции. (Лекция 6-7)

Содержание
- 2. Особенности механизмов контроля инициации транскрипции Экспрессия генов осуществляется с разной интенсивностью на протяжении жизни бактерии. Ингибирование
- 3. «Принятие решения». Синтезировать или не синтезировать?
- 4. Особенности белков-активаторов На оперон могут воздействовать одновременно специфический и общий активаторы. Активаторы не являются особой группой
- 5. Энхансеры и удаленные сайты Сайты активации расположенные на большом удалении от промотора – Энхансеры. Обнаружены у
- 6. Аттенуация на примере оперона синтеза триптофана После начала транскрипции лидерного региона процесс может быть остановлен –
- 7. Аттенуация на примере оперона синтеза триптофана Причина – содержание 2 кодонов Trp в области 1, если
- 8. Аттенуация на примере оперона синтеза триптофана
- 11. РНК-переключатели (riboswitch) Особая форма аттеннуации в ходе транскрипции без участия рибосом. Лидерный регион иРНК (РНК-переключатель) формирует
- 12. РНК-переключатели
- 13. Регуляция путем рекомбинации ДНК Сайт-специфическая реципрокная рекомбинация с перестройкой структуры генов может многократно включать и отключать
- 14. Регуляция и метаболизм РНК Обычно РНК у прокариот транслируется в том же виде что синтезируется. Исключение
- 15. Регуляция на уровне трансляции РНК-переключатели грамотрицательных бактерий регулируют трансляцию иРНК, блокируя ее инициацию. У грамположительных бактерий
- 16. РНК-переключатели В отсутствие метаболита (лиганда), комплиментарные последовательности на лидерной РНК связаны, рибосома имеет доступ к Шайн-Дельгарно
- 17. Малые РНК Известно множество молекул РНК не относящихся к тРНК, иРНК или рРНК – малые РНК.
- 18. Малые РНК Трансляция гена ompF (белка порина) регулируется антисмысловой РНК MicF – продукта гена micF.
- 19. Общие регуляторные системы Помимо систем регуляции на уровне отдельных оперонов, изменения условий среды требует согласованной работы
- 20. Группы оперонов Опероны (сходные функции), которые регулируются общим регуляторным протеином – регулон. Опероны (общее направление), которые
- 21. Катаболическая репрессия Наличие предпочтительного источника углерода (углерода) требует скоординированной репресии генов, кодирующий альтернативные пути получения энергии
- 22. цАМФ Координация катаболических оперонов осуществляется при помощи цАМФ-рецепторного протеина. Активация происходит при связывании цАМФ. Уровень цАМФ
- 23. Катаболическая репрессия Катаболические опероны контролируются 2 типами белков: 1) Специфическим для оперона 2) Общий механизм –
- 24. Катаболическая репрессия на примере лактозного оперона
- 25. Катаболическая репрессия на примере лактозного оперона
- 26. «Ощущение кворума» (Quorum Sensing) В микробных популяциях большую роль в координации экспрессии генов играют сигнальные молекулы.
- 27. «Ощущение кворума» (Quorum Sensing) Небольшие количества АГЛ в популяциях с низкой плотностью микробных клеток выходит во
- 28. Quorum Sensing у Vibrio fischeri
- 29. Регуляция споруляции Переход к спорообразованию в условиях недостатка питательных веществ запускается автофосфорилазой KinА. KinА в свою
- 30. Регуляция экспрессии у Эукариот и Архей Подобно Эубактериям, регуляция у Эукариот и Архей осуществляется на этапах
- 31. Регуляторные факторы транскрипции Эукариот
- 32. Регуляторные факторы транскрипции Эукариот
- 33. Транскрипция
- 34. Транскрипция ингибирована
- 35. Природа взаимодействий белок-ДНК Регуляторные белки узнают последовательности ДНК (8-20 т.п.н.) из миллионов нуклеотидных пар. В структуре
- 37. Скачать презентацию

Особенности механизмов контроля инициации транскрипции
Экспрессия генов осуществляется с разной интенсивностью на
Особенности механизмов контроля инициации транскрипции
Экспрессия генов осуществляется с разной интенсивностью на
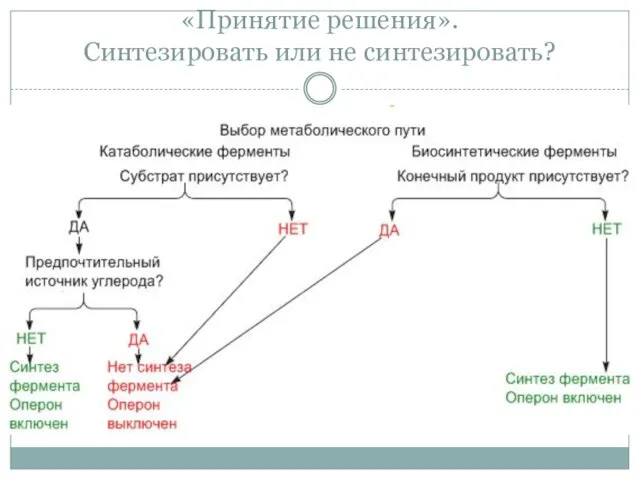
«Принятие решения».
Синтезировать или не синтезировать?
«Принятие решения».
Синтезировать или не синтезировать?

Особенности белков-активаторов
На оперон могут воздействовать одновременно специфический и общий активаторы.
Активаторы
Особенности белков-активаторов
На оперон могут воздействовать одновременно специфический и общий активаторы.
Активаторы

Энхансеры и удаленные сайты
Сайты активации расположенные на большом удалении от промотора
Энхансеры и удаленные сайты
Сайты активации расположенные на большом удалении от промотора

Аттенуация
на примере оперона синтеза триптофана
После начала транскрипции лидерного региона процесс
Аттенуация
на примере оперона синтеза триптофана
После начала транскрипции лидерного региона процесс

Аттенуация
на примере оперона синтеза триптофана
Причина – содержание 2 кодонов Trp
Аттенуация
на примере оперона синтеза триптофана
Причина – содержание 2 кодонов Trp
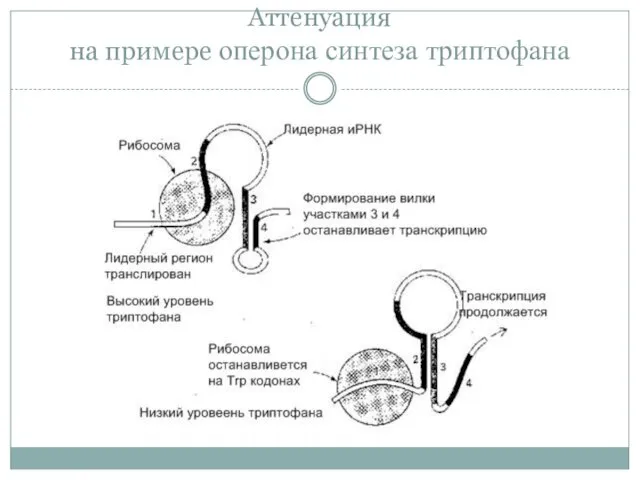
Аттенуация
на примере оперона синтеза триптофана
Аттенуация
на примере оперона синтеза триптофана
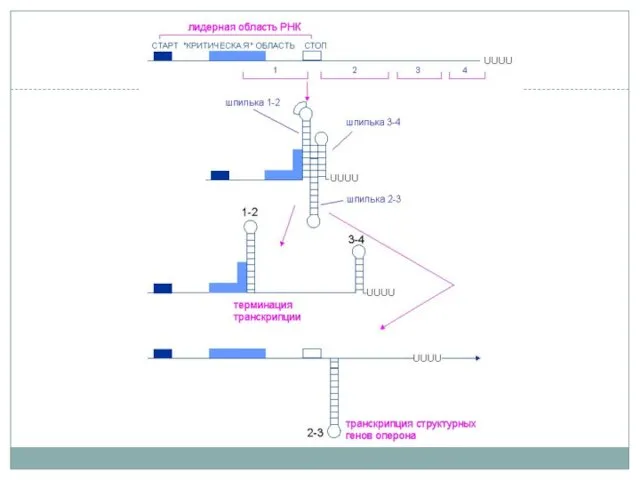
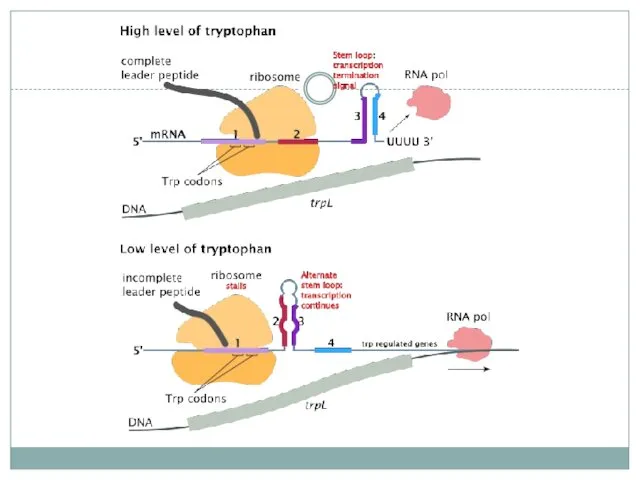

РНК-переключатели (riboswitch)
Особая форма аттеннуации в ходе транскрипции без участия рибосом. Лидерный
РНК-переключатели (riboswitch)
Особая форма аттеннуации в ходе транскрипции без участия рибосом. Лидерный
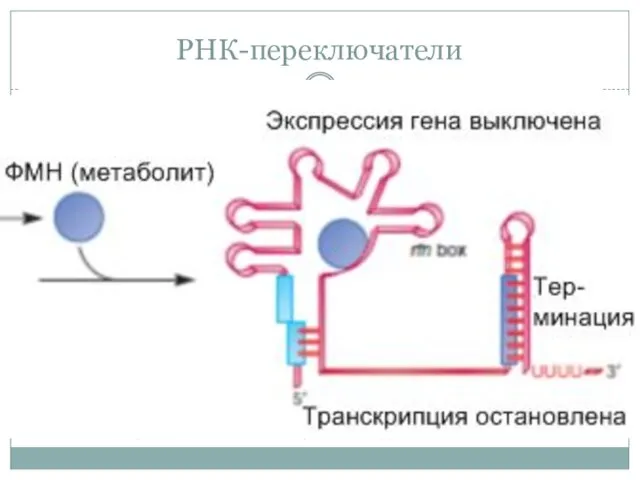
РНК-переключатели
РНК-переключатели

Регуляция путем рекомбинации ДНК
Сайт-специфическая реципрокная рекомбинация с перестройкой структуры генов может
Регуляция путем рекомбинации ДНК
Сайт-специфическая реципрокная рекомбинация с перестройкой структуры генов может

Регуляция и метаболизм РНК
Обычно РНК у прокариот транслируется в том же
Регуляция и метаболизм РНК
Обычно РНК у прокариот транслируется в том же

Регуляция на уровне трансляции
РНК-переключатели грамотрицательных бактерий регулируют трансляцию иРНК, блокируя ее
Регуляция на уровне трансляции
РНК-переключатели грамотрицательных бактерий регулируют трансляцию иРНК, блокируя ее
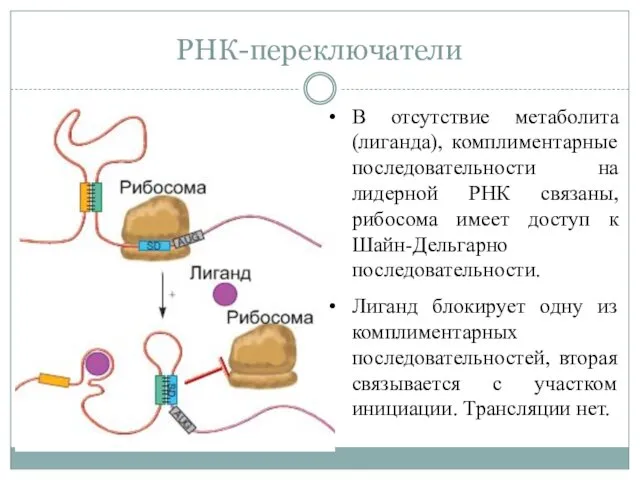
РНК-переключатели
В отсутствие метаболита (лиганда), комплиментарные последовательности на лидерной РНК связаны, рибосома
РНК-переключатели
В отсутствие метаболита (лиганда), комплиментарные последовательности на лидерной РНК связаны, рибосома

Малые РНК
Известно множество молекул РНК не относящихся к тРНК, иРНК или
Малые РНК
Известно множество молекул РНК не относящихся к тРНК, иРНК или
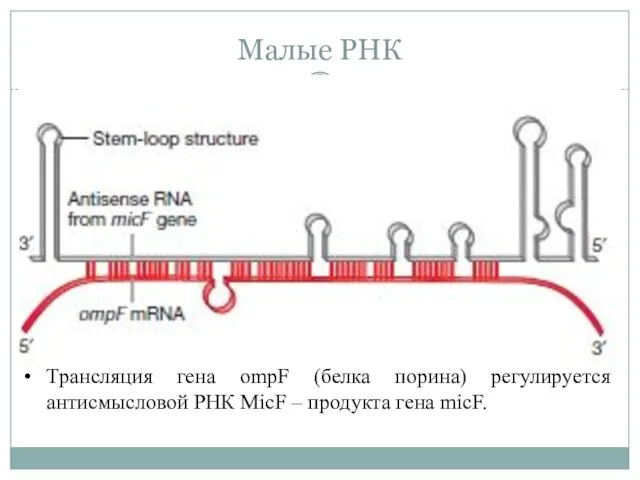
Малые РНК
Трансляция гена ompF (белка порина) регулируется антисмысловой РНК MicF –
Малые РНК
Трансляция гена ompF (белка порина) регулируется антисмысловой РНК MicF –

Общие регуляторные системы
Помимо систем регуляции на уровне отдельных оперонов, изменения условий
Общие регуляторные системы
Помимо систем регуляции на уровне отдельных оперонов, изменения условий

Группы оперонов
Опероны (сходные функции), которые регулируются общим регуляторным протеином – регулон.
Опероны
Группы оперонов
Опероны (сходные функции), которые регулируются общим регуляторным протеином – регулон.
Опероны
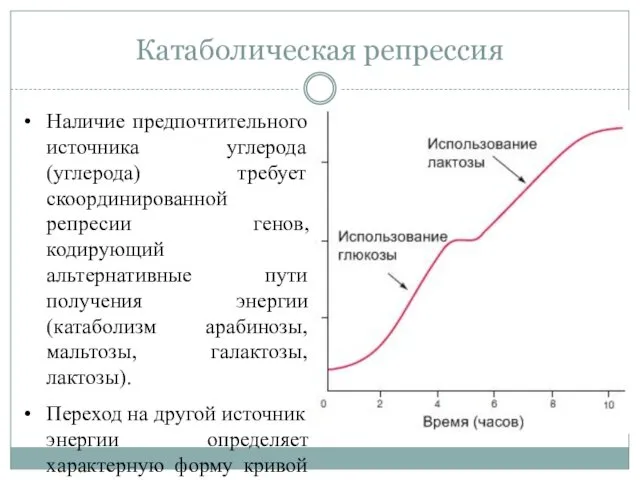
Катаболическая репрессия
Наличие предпочтительного источника углерода (углерода) требует скоординированной репресии генов, кодирующий
Катаболическая репрессия
Наличие предпочтительного источника углерода (углерода) требует скоординированной репресии генов, кодирующий
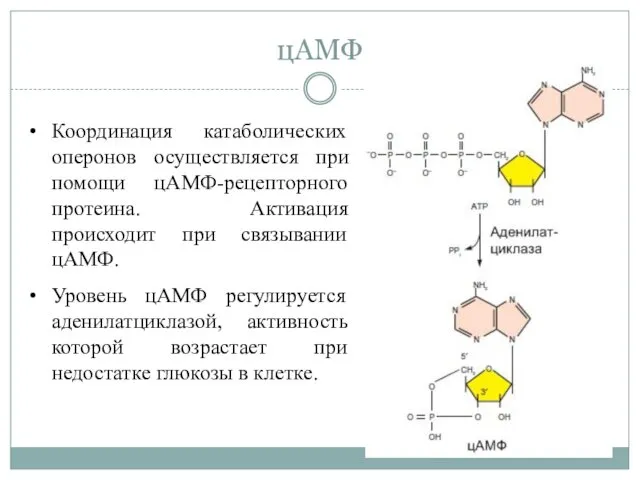
цАМФ
Координация катаболических оперонов осуществляется при помощи цАМФ-рецепторного протеина. Активация происходит при
цАМФ
Координация катаболических оперонов осуществляется при помощи цАМФ-рецепторного протеина. Активация происходит при
Катаболическая репрессия
Катаболические опероны контролируются 2 типами белков:
1) Специфическим для оперона
2) Общий
Катаболическая репрессия
Катаболические опероны контролируются 2 типами белков:
1) Специфическим для оперона
2) Общий
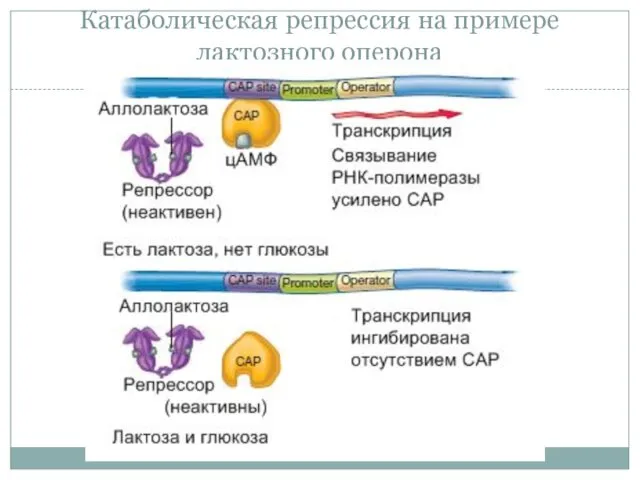
Катаболическая репрессия на примере лактозного оперона
Катаболическая репрессия на примере лактозного оперона
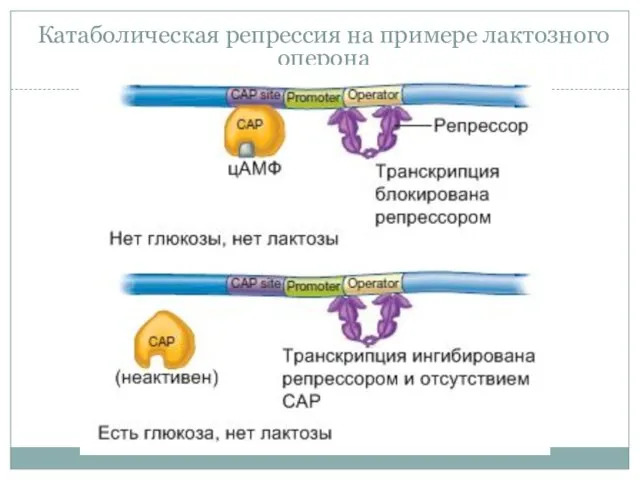
Катаболическая репрессия на примере лактозного оперона
Катаболическая репрессия на примере лактозного оперона

«Ощущение кворума» (Quorum Sensing)
В микробных популяциях большую роль в координации
«Ощущение кворума» (Quorum Sensing)
В микробных популяциях большую роль в координации

«Ощущение кворума» (Quorum Sensing)
Небольшие количества АГЛ в популяциях с низкой
«Ощущение кворума» (Quorum Sensing)
Небольшие количества АГЛ в популяциях с низкой
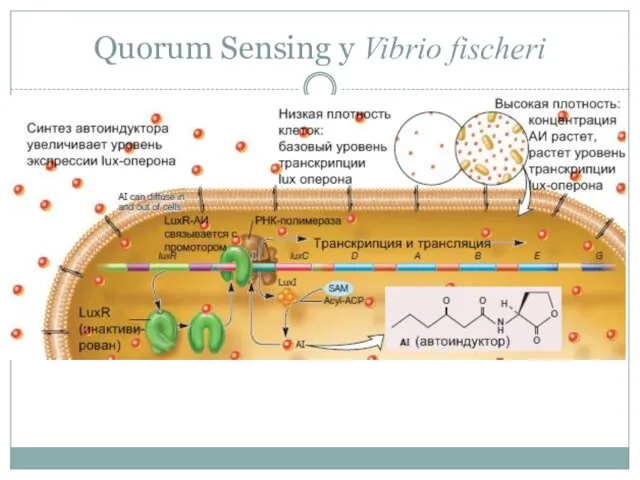
Quorum Sensing у Vibrio fischeri
Quorum Sensing у Vibrio fischeri
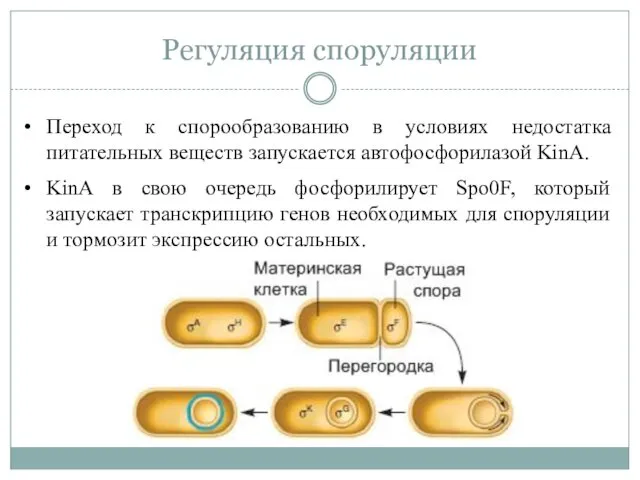
Регуляция споруляции
Переход к спорообразованию в условиях недостатка питательных веществ запускается автофосфорилазой
Регуляция споруляции
Переход к спорообразованию в условиях недостатка питательных веществ запускается автофосфорилазой

Регуляция экспрессии у Эукариот и Архей
Подобно Эубактериям, регуляция у Эукариот и
Регуляция экспрессии у Эукариот и Архей
Подобно Эубактериям, регуляция у Эукариот и
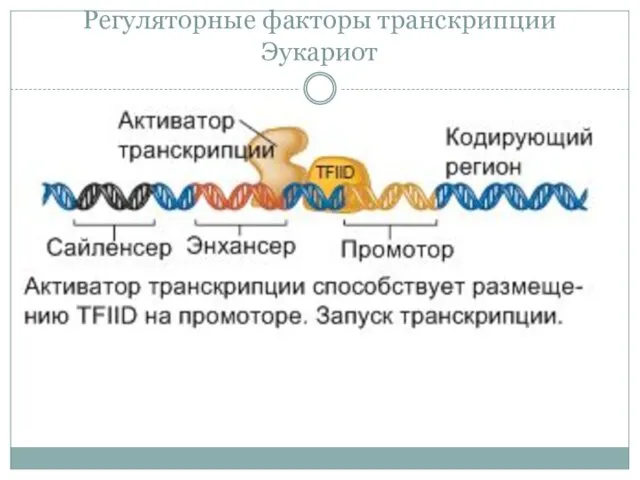
Регуляторные факторы транскрипции Эукариот
Регуляторные факторы транскрипции Эукариот
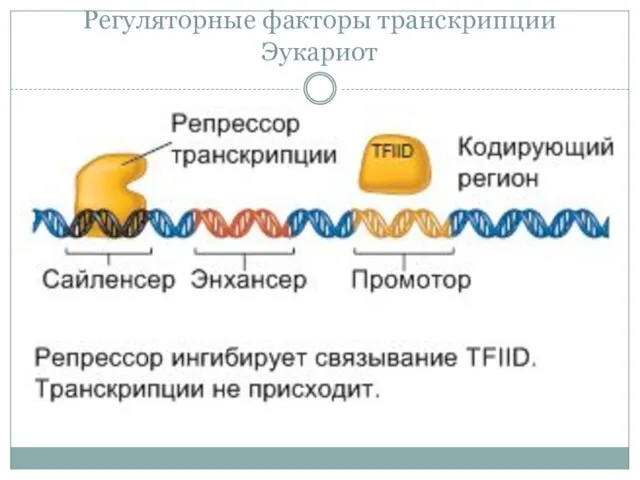
Регуляторные факторы транскрипции Эукариот
Регуляторные факторы транскрипции Эукариот
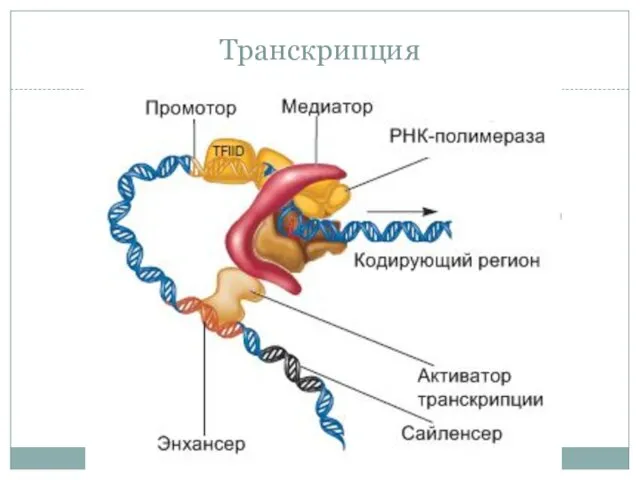
Транскрипция
Транскрипция
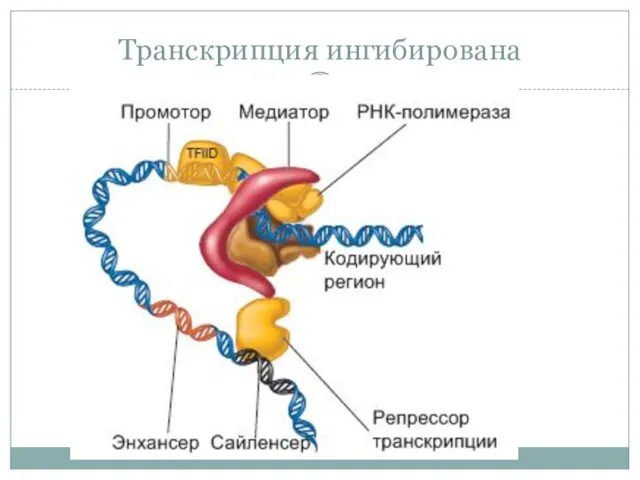
Транскрипция ингибирована
Транскрипция ингибирована

Природа взаимодействий белок-ДНК
Регуляторные белки узнают последовательности ДНК (8-20 т.п.н.) из миллионов
Природа взаимодействий белок-ДНК
Регуляторные белки узнают последовательности ДНК (8-20 т.п.н.) из миллионов
 Обзор мероприятий Месячника Школьных библиотек
Обзор мероприятий Месячника Школьных библиотек Как моя семья пережила Великую Отечественную войну. Вязина Ксения
Как моя семья пережила Великую Отечественную войну. Вязина Ксения Ислам. Пророк Мухаммед
Ислам. Пророк Мухаммед Презентация 1 сентября
Презентация 1 сентября Презентация к сказке В гости к солнышку
Презентация к сказке В гости к солнышку Световые рефлексы глазного дна
Световые рефлексы глазного дна Минутка чистописания. Части речи
Минутка чистописания. Части речи Дополнительная общеразвивающая программа Бу-магия, как средство развития художественно-творческих способностей обучающихся
Дополнительная общеразвивающая программа Бу-магия, как средство развития художественно-творческих способностей обучающихся Microsoft Office. Организация работы с документацией. Word. Excel
Microsoft Office. Организация работы с документацией. Word. Excel Презентация по теме Суша
Презентация по теме Суша доклад 1 Пульсар, Гравитация и Пульсар продолжение публикации из новой Конструктивной Теории Всего
доклад 1 Пульсар, Гравитация и Пульсар продолжение публикации из новой Конструктивной Теории Всего открытка
открытка Аспекты изучения простого предложения
Аспекты изучения простого предложения Сырье для производства строительных материалов. Виды сырья
Сырье для производства строительных материалов. Виды сырья Қытай мен Қазақстанның байланысы
Қытай мен Қазақстанның байланысы Клеточные механизмы иммунного ответа
Клеточные механизмы иммунного ответа Александр Дюма, роман Три мушкетера. Вопросы на знание текста
Александр Дюма, роман Три мушкетера. Вопросы на знание текста Презентация Бурятский национальный костюм
Презентация Бурятский национальный костюм Наслідки промислової революції
Наслідки промислової революції Введение в HTML и CSS. Текст и списки. Позиционирование
Введение в HTML и CSS. Текст и списки. Позиционирование Компания Talivenda. Производство нашей продукции
Компания Talivenda. Производство нашей продукции Паломническая поездка в Грузию
Паломническая поездка в Грузию Трудовое право
Трудовое право How AI* affects NPT-regime? And other emerging technologies
How AI* affects NPT-regime? And other emerging technologies Организация проектной деятельности в преподавании курса ОРКСЭ
Организация проектной деятельности в преподавании курса ОРКСЭ Согласные звуки и буквы
Согласные звуки и буквы Применение генной инженерии в селекции пробиотических микроорганизмов
Применение генной инженерии в селекции пробиотических микроорганизмов Организация Массолит в романе М.А. Булгакова Мастер и Маргарита
Организация Массолит в романе М.А. Булгакова Мастер и Маргарита