- Нестабильность генома неопластических клеток. Лекция 5

Содержание
- 2. Нестабильность генома (мутации и эпигенетические изменения) Нечувствительность к рост-ингибирующим сигналам (приобретение бессмертия и др.) Постоянная инициация
- 3. Онкогенез – многоступенчатый процесс отбора клеток с рядом генетических изменений, определяющих свойства злокачественной клетки
- 4. Легко обнаруживаемые проявления нестабильности генома опухолевых клеток: (SKY – многоцветное спектральное кариотипирование) Множественные изменения кариотипа Bat25
- 5. В первых исследованиях геномов и транскриптомов опухолевых тканей (Wood еt al, Science, 2007; Jones et al.,
- 7. Многочисленные изменения геномов/транскриптомов опухолевых клеток являются отражением их генетической нестабильности и клонального разнообразия формируемых ими новообразований
- 8. Хромотрипсис Одномоментный клеточный кризис, ведущий к образованию сложных комплексных хромосомных перестроек (10-100) в одной или нескольких
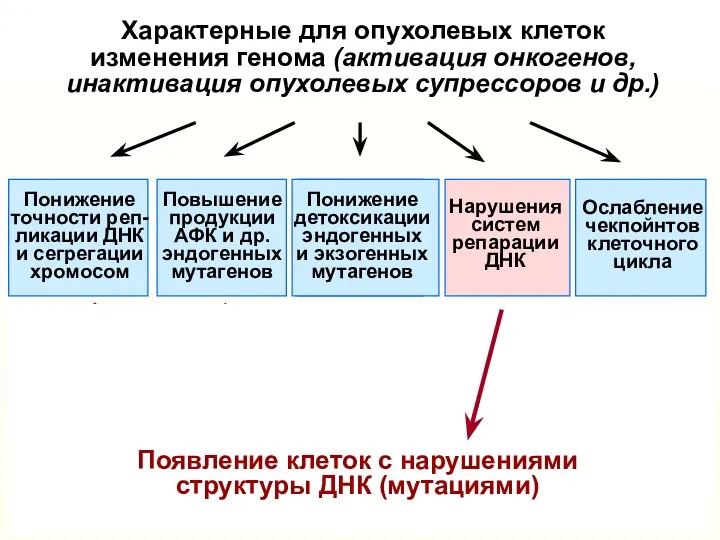
- 9. Характерные для опухолевых клеток изменения генома (активация онкогенов, инактивация опухолевых супрессоров и др.) Понижение точности реп-
- 10. Характерные для опухолевых клеток изменения генома (активация онкогенов, инактивация опухолевых супрессоров и др.) Понижение точности реп-
- 11. Мутагенные факторы: Эндогенные – а) ошибки репликации ДНК и митоза; б) внутриклеточные свободные радикалы (активные формы
- 12. Нарушения экспрессии онкогенов и опухолевых супрессоров увеличивают частоту возникновения генетических ошибок Proofreading by polymerase δ Гетерозиготные
- 13. Нарушения экспрессии онкогенов и опухолевых супрессоров увеличивают частоту возникновения генетических ошибок Неправильная сегрегация хромосом Изменения числа
- 14. Трехполюсный митоз Амплификация центросом Норма p21Cip1 CycE/ Cdk2 Многократная репликация центросом p53 p21Cip1 CycE/ Cdk2 Однократная
- 15. Анеуплоидия (~ 90% опухолей)
- 16. Эндогенные мутагенные факторы: а) ошибки репликации ДНК и митоза; б) внутриклеточные свободные радикалы (активные формы кислорода,
- 17. Активные формы кислорода (АФК) образуются в ходе постепенного восстановления молекул кислорода, как побочные продукты ряда внутриклеточных
- 18. Эндогенные источники, воспаление, радиация, хим. агенты О2- Н2О2 Н2О + О2 -ОН + ОН- Повреждения Cu+/Fe2+
- 19. АФК – не только повреждающие, но и важные сигнальные молекулы Окисляя компоненты различных сигнальных путей (фосфатазы
- 20. АФК Окисление ДНК Окисление компонентов сигнальных путей Массивные повреждения ДНК и др. макромолекул Сильное повышение Умеренное
- 21. Окисление оснований в ДНК
- 22. Умеренное повышение содержания активных форм кислорода Активация RAS, МYC и др. Инактивация р53 Инфекция HBV, HCV
- 23. Активированные онкогены RAS Энзиматический синтез супероксидного радикала Повышение содержания активных форм кислорода Подавление антиоксидантной защиты PI3K-Rac-NADPH
- 24. Сигнальный путь Nrf2–Keap1 Sporn and Liby, 2012 Nat Rev Cancer; 12(8): doi:10.1038/nrc3278 Nrf2 – транскрипционный фактор,
- 25. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis Perera and Bardeesy, Nature, 2011, 43-44 When antioxidants
- 26. Контекст-зависимая роль Nrf2 в канцерогенезе Sporn and Liby, 2012. Nat Rev Cancer; 12(8): doi:10.1038/nrc3278
- 27. Нарушения Nrf2–Keap1 сигнализации в опухолях
- 28. Повышение содержания NO Активация RAS, МYC и др. Инактивация р53 Инфекция HBV, HCV и др. Увеличенный
- 29. Характерные для опухолевых клеток изменения генома (активация онкогенов, инактивация опухолевых супрессоров и др.) Понижение точности реп-
- 30. Во многих опухолях понижена экспрессия глютатион-S- трансферазы, детоксифицирующей реактивные соединения путем переноса на них SH-группы глютатиона
- 31. Характерные для опухолевых клеток изменения генома (активация онкогенов, инактивация опухолевых супрессоров и др.) Возникновение различных модификаций
- 32. Повреждение ДНК Белки-сенсоры аномальных структур ДНК Восстановление структуры ДНК Белки репарационных систем Остановка деления, апоптоз Передатчики
- 33. Эксцизионная репарация ДНК
- 34. Репарация двуцепочечных разрывов ДНК Гомологичная рекомбинация (HDR) – поздняя S – G2 фаза Лигирование негомологичных концов
- 35. Репарация двунитевых разрывов ДНК Эксцизионная репарация Ошибки репликации γ-облучение УФ-облучение Распознавание Процессинг Лигирование негомологичных концов ИЛИ
- 36. – MSH2,6, MLH1, PMS2 неспаренных оснований Нарушения репарации ДНК – XPG, XPE, XPA и др. нуклеотидов
- 37. – NBS1, LIG4 Двунитевых разрывов – FANCA, FANCG и др. Межнитевых сшивок Ниймегенский синдром, лимфоидные новообразования
- 38. Характерные для опухолевых клеток изменения генома Генетическая нестабильность Понижение точности реп- ликации ДНК и сегрегации хромосом
- 39. Повреждение ДНК Белки-сенсоры аномальных структур ДНК Восстановление структуры ДНК Белки репарационных систем Остановка деления, апоптоз Передатчики
- 40. Гомологичная рекомбинация Остановка в G2/M (?) Остановка в G1, G2 Остановка в G1, S Двуцепочечный разрыв
- 41. Изменения в делящихся фибробластах мышей c дефектным BRCA2 (brca2-mut/brca2-mut) Метафаза с разрывами и рекомбинациями хромосом Дефектный
- 42. G1 S G2 Митоз Нарушения структуры ДНК p53 Механизмы подавления активности циклин-зависимых киназ и остановки клеточного
- 43. ATM Aтаксия-телеангиоэктазия, лимфолейкозы, лимфомы BRCA1, BRCA2 Рак молочной железы, яичника p53, CHK2 Синдром Ли-Фраумени – различные
- 44. Характерные для опухолевых клеток изменения генома Генетическая нестабильность Понижение точности реп- ликации ДНК и сегрегации хромосом
- 45. Chin and Yeong, Mol. Cell.Biol., 2010: 22–32 Антефазный чекпойнт (Antephase checkpoint) Aurora kinase A, Polo-like kinase
- 46. Метафазный чекпойнт (Spindle checkpoint) Kim and Yu, Semin Cell Dev Biol. 2011, 22(6):551-558. APC/C - Anaphase
- 47. Метафазный чекпойнт (Spindle checkpoint)
- 48. G1 S G2 Митоз Нарушения структуры ДНК Нарушения веретена деления Bub1, Bub3 BubR1, Mad1/2 p53 p21Cip1
- 49. 20-40% различных опухолей (рак желудка, носоглотки и др.) Гиперэкспрессия AURKA Многие новообразования Мутации MAD2 Bub1, Bub3,
- 50. Характерные для опухолевых клеток изменения генома Понижение точности реп- ликации ДНК и сегрегации хромосом Генетическая нестабильность
- 51. Повышенная изменчивость неопластических клеток Увеличение вероятности появления клонов клеток с изменениями структуры и/или копийности генов Увеличение
- 52. Метилирование ДНК – важный механизм тканеспецифичной регуляции генной транскрипции Осуществляется метилтрансферазами (Dnmt1, Dnmt3, Dnmt3a) по 5-му
- 53. Метилирование опухолевых супрессоров в новообразованиях Метилирование промотора гена p16INK4a
- 54. Метилирование CpG-островков в промоторах генов Для опухолевых клеток характерно: Деметилирование промоторов регуляторных генов и одиночных CpG-динуклеотидов
- 55. активация онкогенов RAS инактивация опухолевого супрессора р53 Понижение точности репликации ДНК Повышение уровня эндогенных мутагенов (активные
- 57. Скачать презентацию
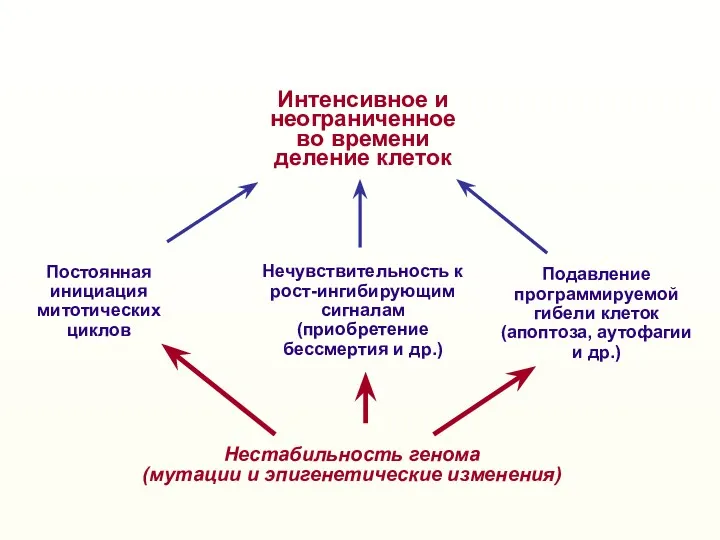
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
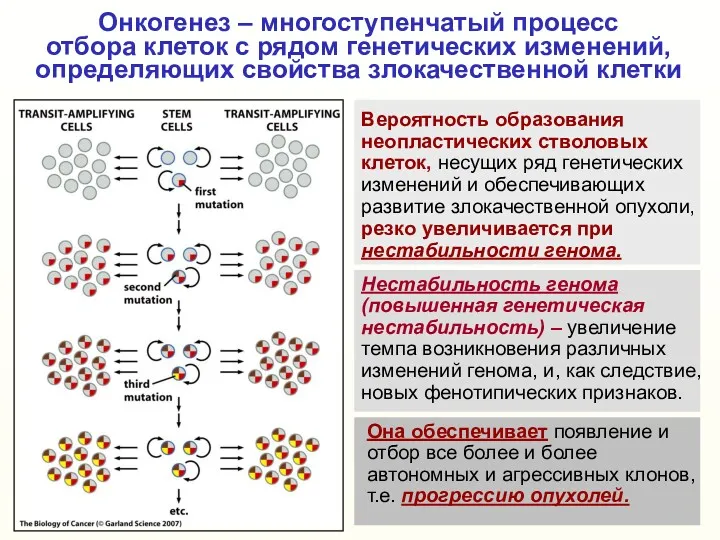
Онкогенез – многоступенчатый процесс
отбора клеток с рядом генетических изменений,
определяющих свойства злокачественной
Онкогенез – многоступенчатый процесс
отбора клеток с рядом генетических изменений,
определяющих свойства злокачественной
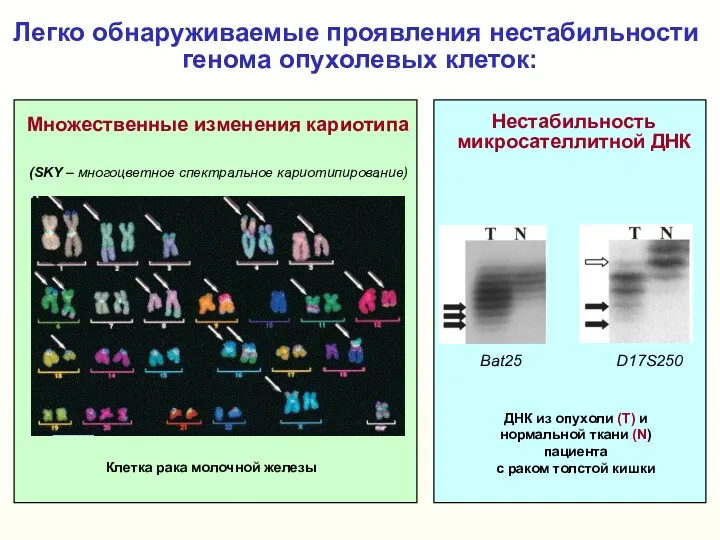
Легко обнаруживаемые проявления нестабильности
генома опухолевых клеток:
(SKY – многоцветное спектральное кариотипирование)
Множественные
Легко обнаруживаемые проявления нестабильности
генома опухолевых клеток:
(SKY – многоцветное спектральное кариотипирование)
Множественные
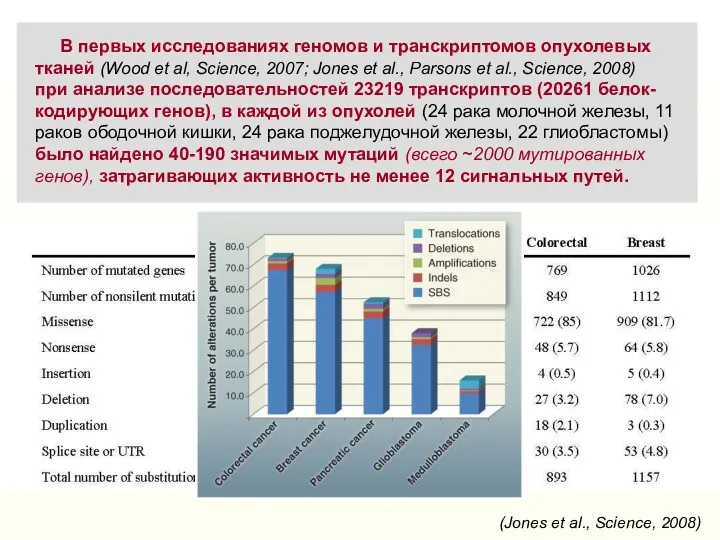
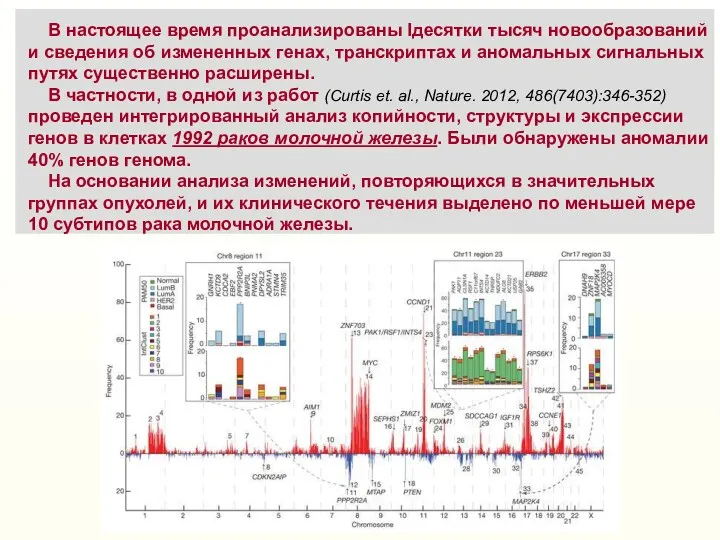
В первых исследованиях геномов и транскриптомов опухолевых тканей (Wood еt
В первых исследованиях геномов и транскриптомов опухолевых тканей (Wood еt
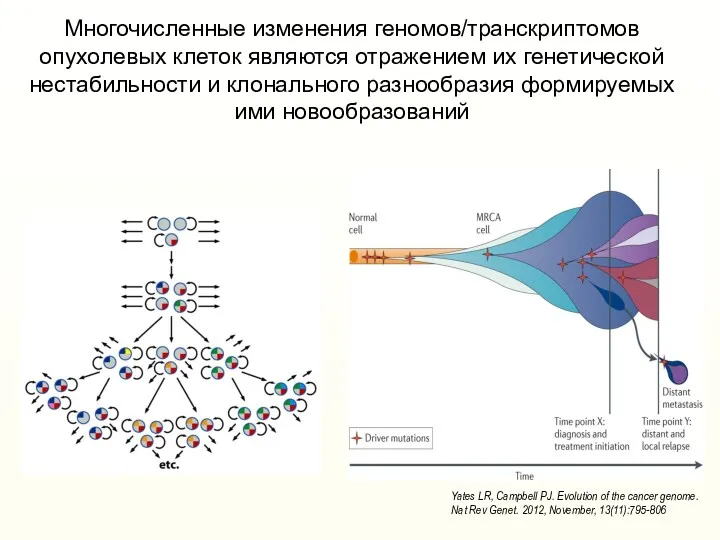
Многочисленные изменения геномов/транскриптомов
опухолевых клеток являются отражением их генетической нестабильности и
Многочисленные изменения геномов/транскриптомов
опухолевых клеток являются отражением их генетической нестабильности и
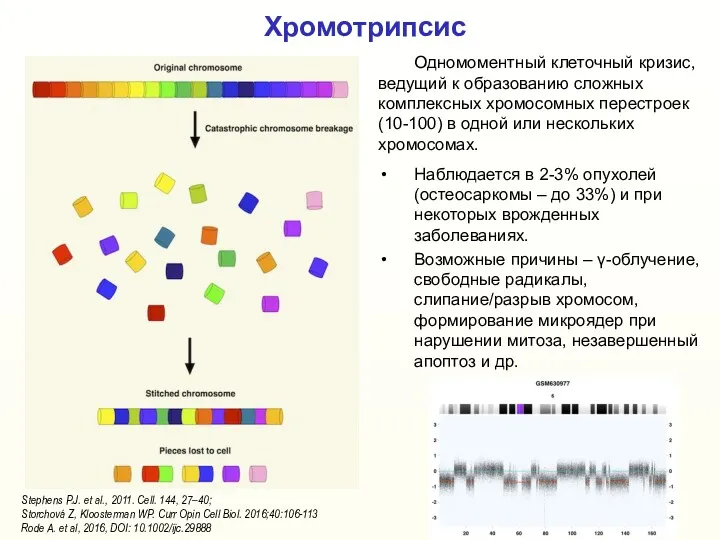
Хромотрипсис
Одномоментный клеточный кризис, ведущий к образованию сложных комплексных хромосомных перестроек (10-100)
Хромотрипсис
Одномоментный клеточный кризис, ведущий к образованию сложных комплексных хромосомных перестроек (10-100)
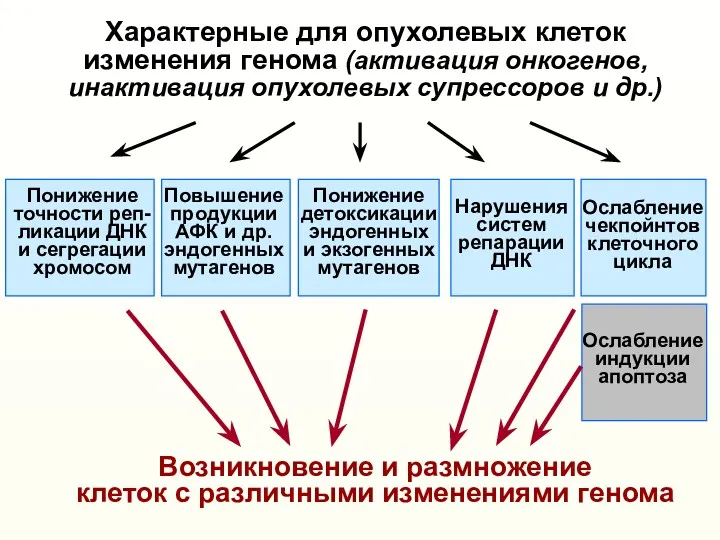
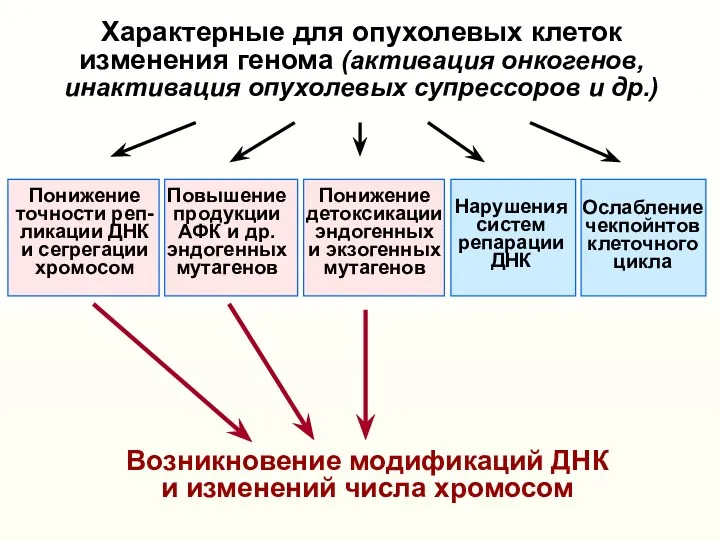
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение

Мутагенные факторы:
Эндогенные – а) ошибки репликации ДНК и митоза;
б) внутриклеточные
Мутагенные факторы:
Эндогенные – а) ошибки репликации ДНК и митоза;
б) внутриклеточные
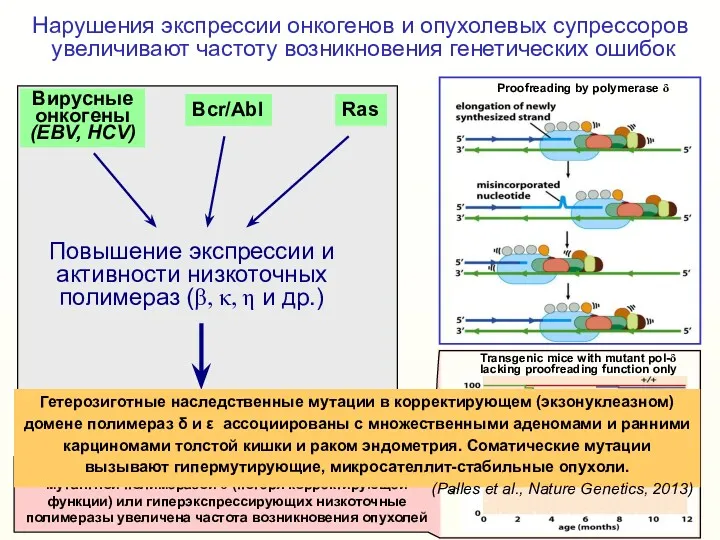
Нарушения экспрессии онкогенов и опухолевых супрессоров
увеличивают частоту возникновения генетических ошибок
Proofreading
Нарушения экспрессии онкогенов и опухолевых супрессоров
увеличивают частоту возникновения генетических ошибок
Proofreading
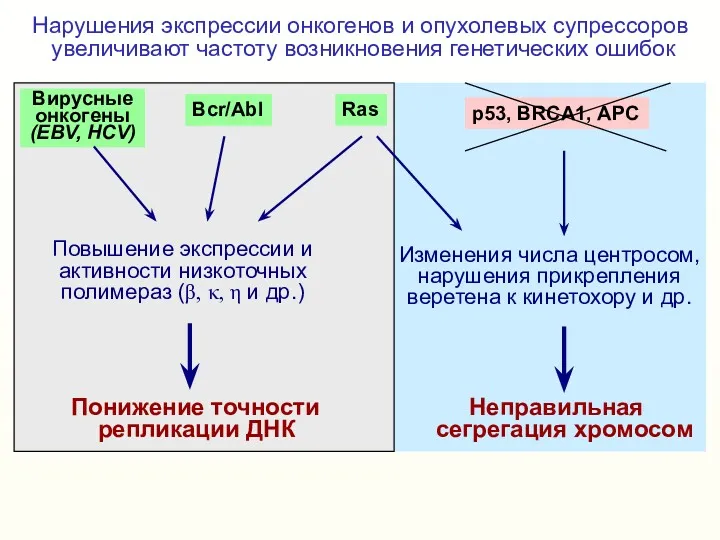
Нарушения экспрессии онкогенов и опухолевых супрессоров
увеличивают частоту возникновения генетических ошибок
Нарушения экспрессии онкогенов и опухолевых супрессоров
увеличивают частоту возникновения генетических ошибок
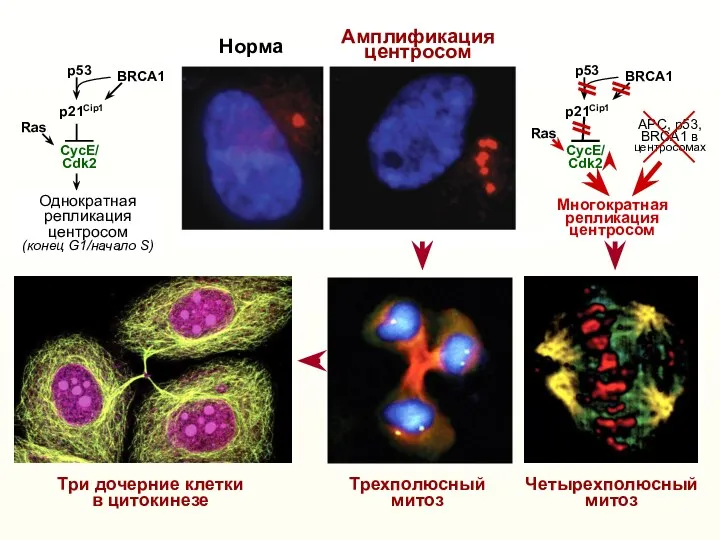
Трехполюсный
митоз
Амплификация
центросом
Норма
p21Cip1
CycE/
Cdk2
Многократная
репликация
центросом
p53
p21Cip1
CycE/
Cdk2
Однократная
репликация
центросом
(конец G1/начало S)
BRCA1
p53
BRCA1
АPC, p53,
BRCA1 в
центросомах
Ras
Ras
Четырехполюсный
митоз
Три дочерние клетки
в цитокинезе
Трехполюсный
митоз
Амплификация
центросом
Норма
p21Cip1
CycE/
Cdk2
Многократная
репликация
центросом
p53
p21Cip1
CycE/
Cdk2
Однократная
репликация
центросом
(конец G1/начало S)
BRCA1
p53
BRCA1
АPC, p53,
BRCA1 в
центросомах
Ras
Ras
Четырехполюсный
митоз
Три дочерние клетки
в цитокинезе
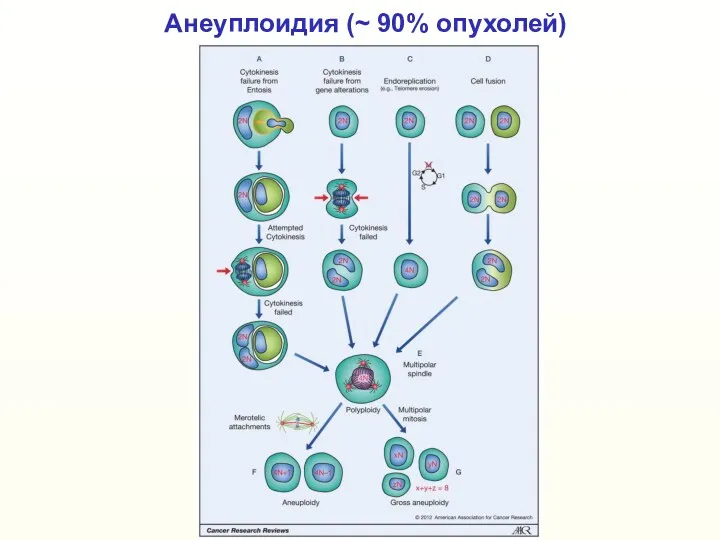
Анеуплоидия (~ 90% опухолей)
Анеуплоидия (~ 90% опухолей)

Эндогенные мутагенные факторы:
а) ошибки репликации ДНК и митоза;
б) внутриклеточные свободные радикалы
Эндогенные мутагенные факторы:
а) ошибки репликации ДНК и митоза;
б) внутриклеточные свободные радикалы

Активные формы кислорода (АФК) образуются в ходе постепенного восстановления молекул
Активные формы кислорода (АФК) образуются в ходе постепенного восстановления молекул
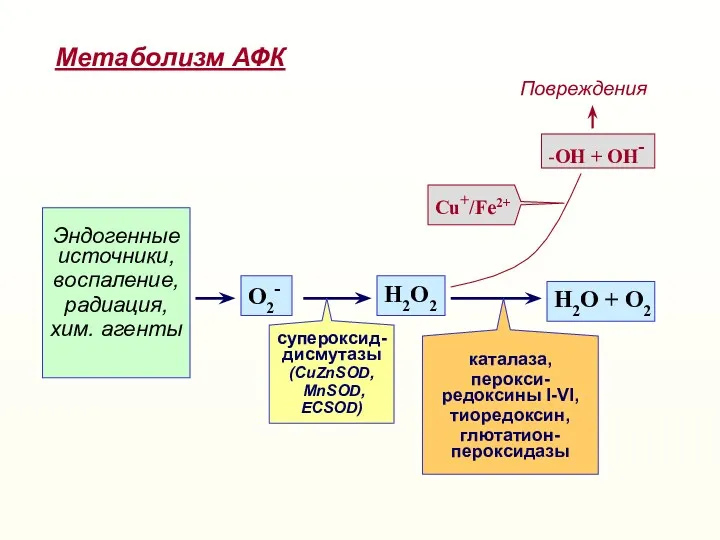
Эндогенные источники,
воспаление,
радиация,
хим. агенты
О2-
Н2О2
Н2О + О2
-ОН + ОН-
Повреждения
Cu+/Fe2+
супероксид-дисмутазы
(CuZnSOD,
MnSOD,
ECSOD)
каталаза,
перокси-редоксины I-VI,
тиоредоксин,
глютатион-пероксидазы
Метаболизм АФК
Эндогенные источники,
воспаление,
радиация,
хим. агенты
О2-
Н2О2
Н2О + О2
-ОН + ОН-
Повреждения
Cu+/Fe2+
супероксид-дисмутазы
(CuZnSOD,
MnSOD,
ECSOD)
каталаза,
перокси-редоксины I-VI,
тиоредоксин,
глютатион-пероксидазы
Метаболизм АФК

АФК – не только повреждающие, но
и важные сигнальные молекулы
Окисляя компоненты
АФК – не только повреждающие, но
и важные сигнальные молекулы
Окисляя компоненты
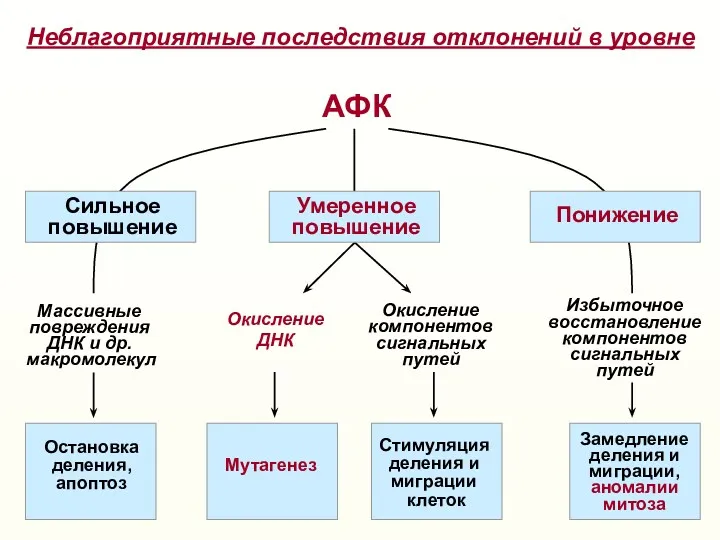
АФК
Окисление
ДНК
Окисление
компонентов
сигнальных
путей
Массивные
повреждения
ДНК и др.
макромолекул
Сильное
повышение
Умеренное
повышение
Понижение
Избыточное
восстановление
компонентов
сигнальных
путей
Неблагоприятные последствия отклонений в уровне
Остановка
деления,
апоптоз
Мутагенез
Стимуляция
деления и
миграции
клеток
Замедление
деления
АФК
Окисление
ДНК
Окисление
компонентов
сигнальных
путей
Массивные
повреждения
ДНК и др.
макромолекул
Сильное
повышение
Умеренное
повышение
Понижение
Избыточное
восстановление
компонентов
сигнальных
путей
Неблагоприятные последствия отклонений в уровне
Остановка
деления,
апоптоз
Мутагенез
Стимуляция
деления и
миграции
клеток
Замедление
деления
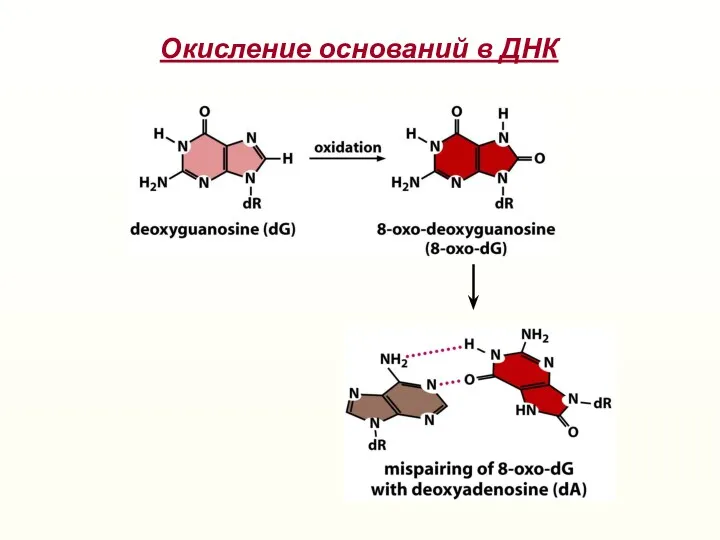
Окисление оснований в ДНК
Окисление оснований в ДНК
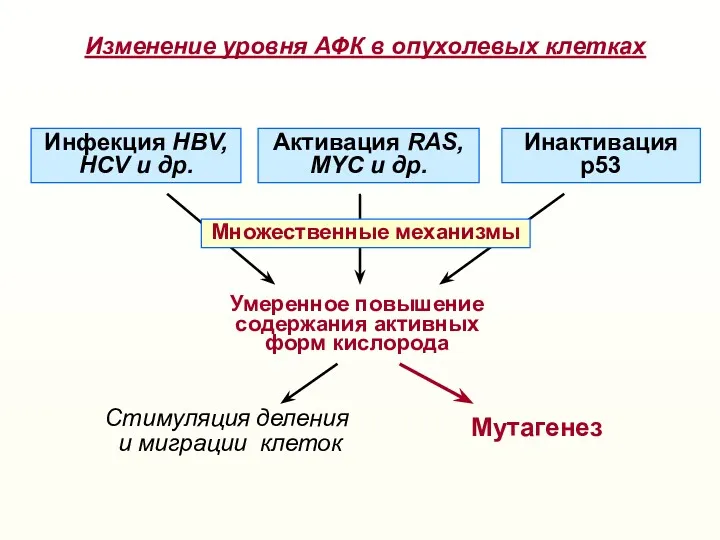
Умеренное повышение содержания активных
форм кислорода
Активация RAS,
МYC и др.
Инактивация р53
Умеренное повышение содержания активных
форм кислорода
Активация RAS,
МYC и др.
Инактивация р53
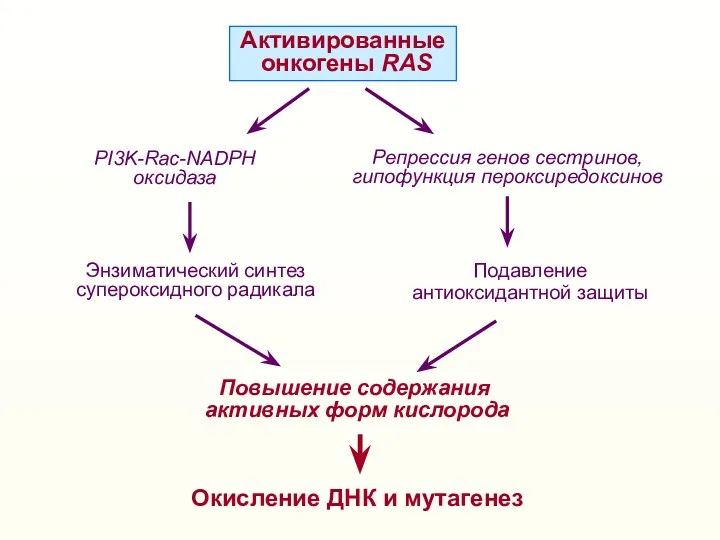
Активированные
онкогены RAS
Энзиматический синтез супероксидного радикала
Повышение содержания
активных форм кислорода
Активированные
онкогены RAS
Энзиматический синтез супероксидного радикала
Повышение содержания
активных форм кислорода
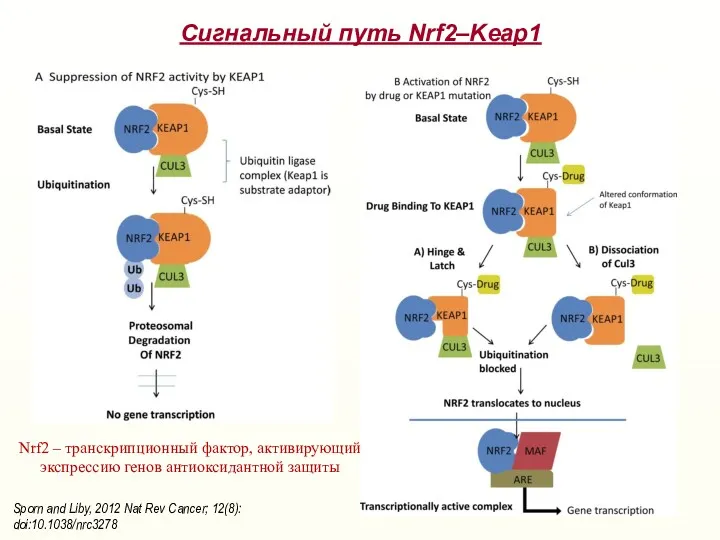
Сигнальный путь Nrf2–Keap1
Sporn and Liby, 2012 Nat Rev Cancer; 12(8): doi:10.1038/nrc3278
Nrf2
Сигнальный путь Nrf2–Keap1
Sporn and Liby, 2012 Nat Rev Cancer; 12(8): doi:10.1038/nrc3278
Nrf2
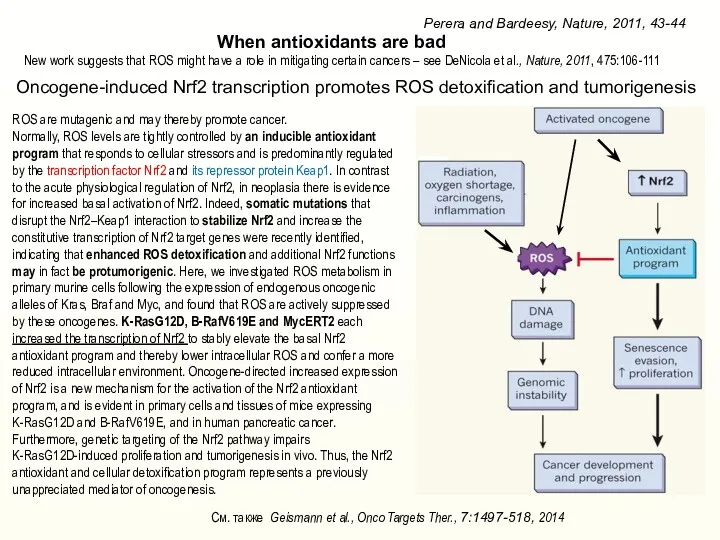
Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis
Perera and Bardeesy, Nature,
Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis
Perera and Bardeesy, Nature,
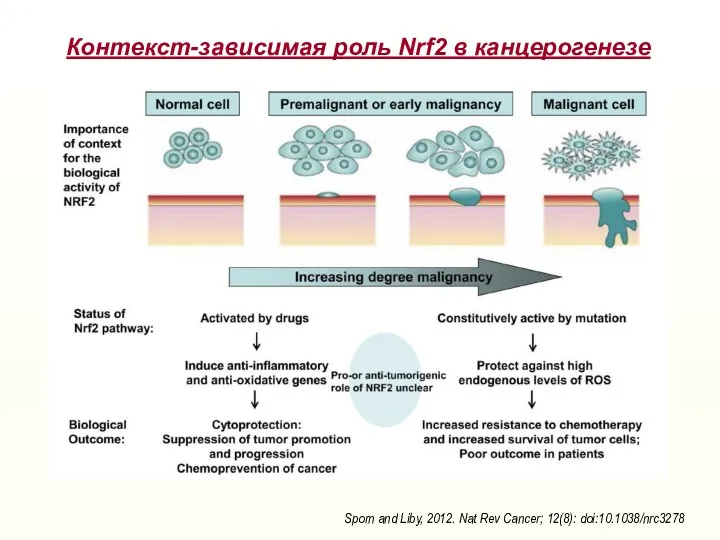
Контекст-зависимая роль Nrf2 в канцерогенезе
Sporn and Liby, 2012. Nat Rev Cancer;
Контекст-зависимая роль Nrf2 в канцерогенезе
Sporn and Liby, 2012. Nat Rev Cancer;
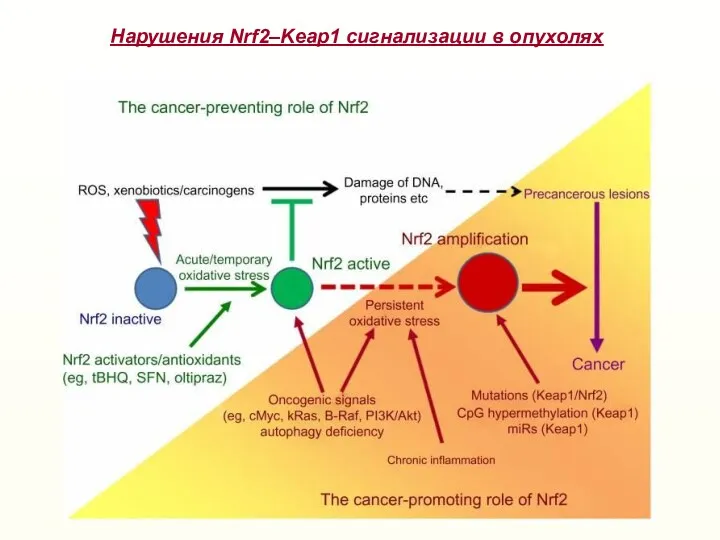
Нарушения Nrf2–Keap1 сигнализации в опухолях
Нарушения Nrf2–Keap1 сигнализации в опухолях
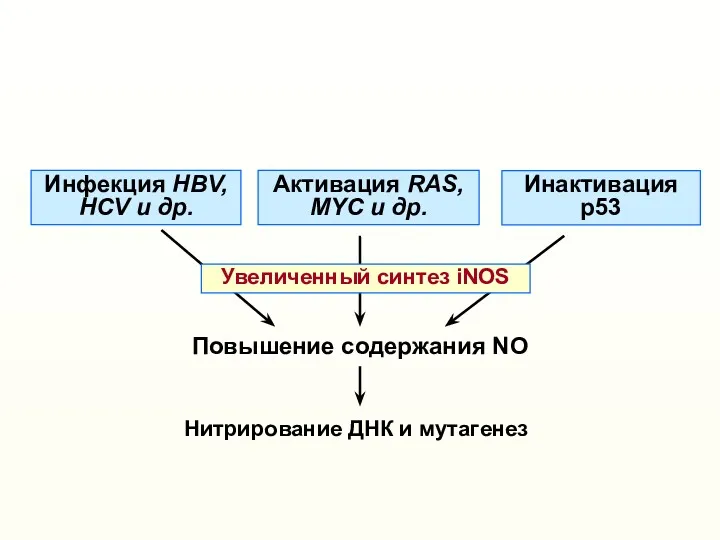
Повышение содержания NO
Активация RAS,
МYC и др.
Инактивация р53
Инфекция HBV,
HCV
Повышение содержания NO
Активация RAS,
МYC и др.
Инактивация р53
Инфекция HBV,
HCV
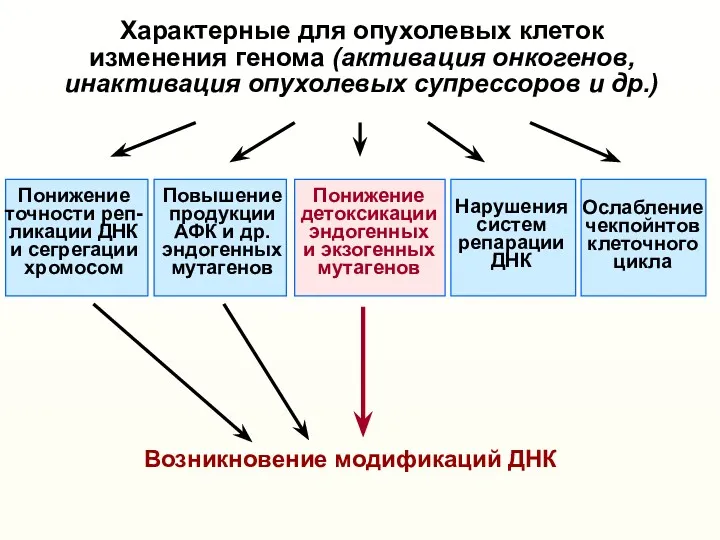
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Понижение
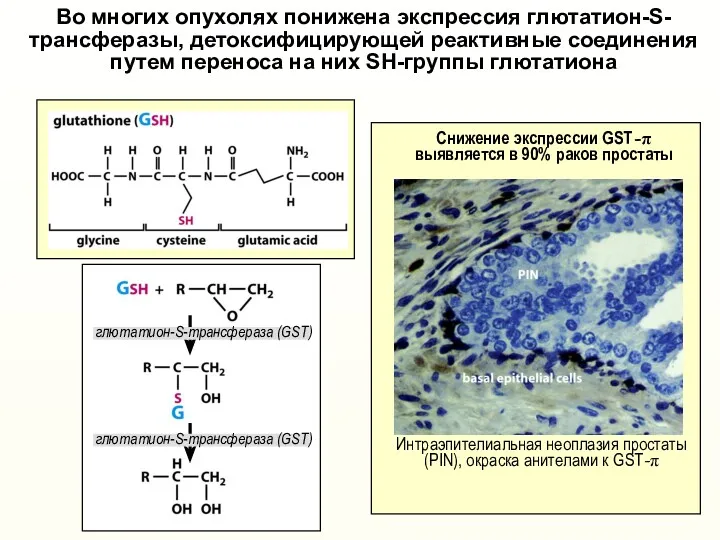
Во многих опухолях понижена экспрессия глютатион-S-
трансферазы, детоксифицирующей реактивные соединения
путем переноса на
Во многих опухолях понижена экспрессия глютатион-S-
трансферазы, детоксифицирующей реактивные соединения
путем переноса на
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Возникновение
Характерные для опухолевых клеток
изменения генома (активация онкогенов,
инактивация опухолевых супрессоров и др.)
Возникновение
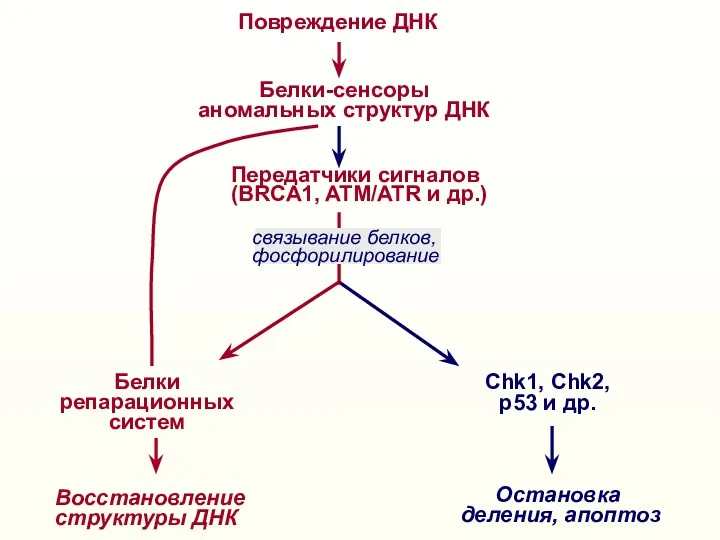
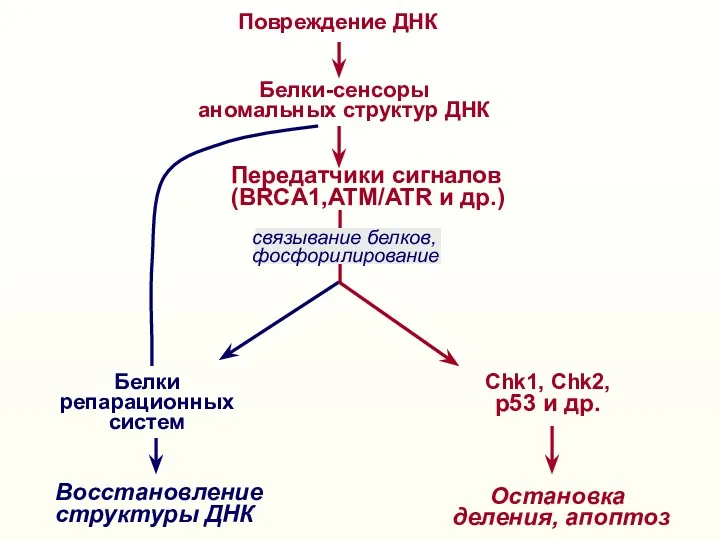
Повреждение ДНК
Белки-сенсоры
аномальных структур ДНК
Восстановление
структуры ДНК
Белки
репарационных
систем
Остановка
деления, апоптоз
Передатчики сигналов
(BRCA1, ATM/ATR
Повреждение ДНК
Белки-сенсоры
аномальных структур ДНК
Восстановление
структуры ДНК
Белки
репарационных
систем
Остановка
деления, апоптоз
Передатчики сигналов
(BRCA1, ATM/ATR
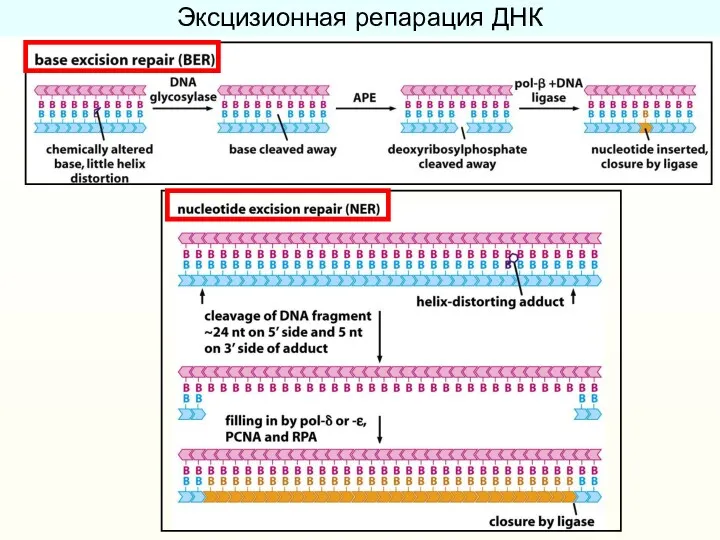
Эксцизионная репарация ДНК
Эксцизионная репарация ДНК
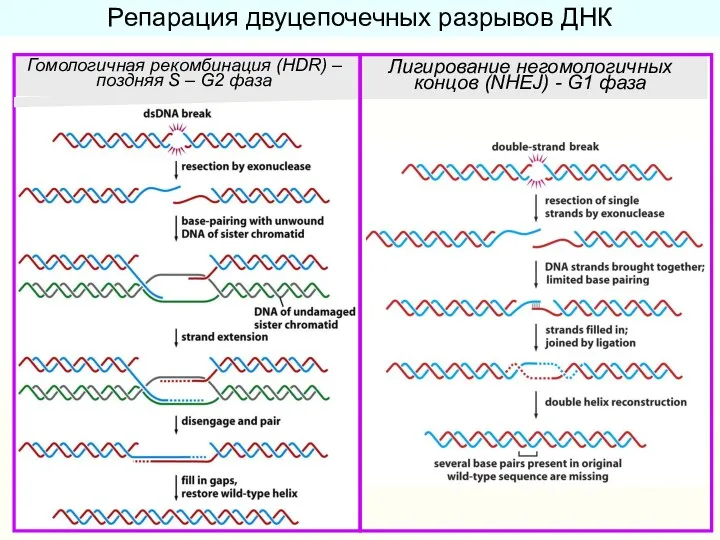
Репарация двуцепочечных разрывов ДНК
Гомологичная рекомбинация (HDR) – поздняя S – G2
Репарация двуцепочечных разрывов ДНК
Гомологичная рекомбинация (HDR) – поздняя S – G2
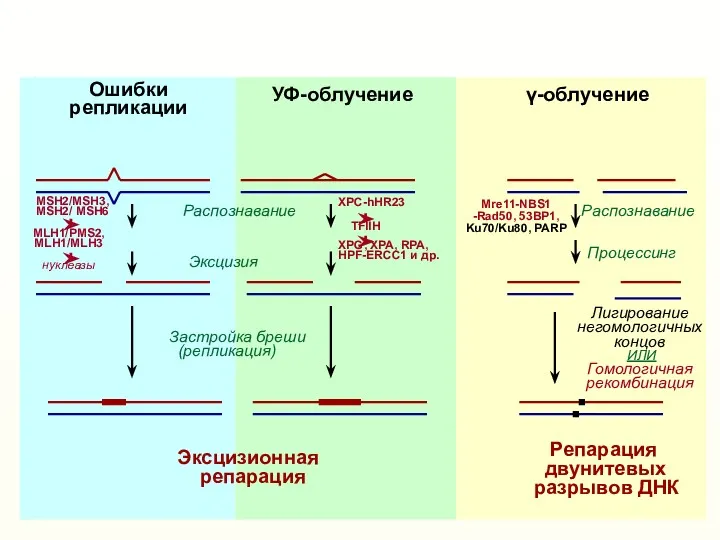
Репарация
двунитевых
разрывов ДНК
Эксцизионная
репарация
Ошибки
репликации
γ-облучение
УФ-облучение
Распознавание
Процессинг
Лигирование
негомологичных
концов
ИЛИ
Гомологичная
рекомбинация
Распознавание
Эксцизия
Застройка бреши
(репликация)
MSH2/MSH3,
MSH2/
Репарация
двунитевых
разрывов ДНК
Эксцизионная
репарация
Ошибки
репликации
γ-облучение
УФ-облучение
Распознавание
Процессинг
Лигирование
негомологичных
концов
ИЛИ
Гомологичная
рекомбинация
Распознавание
Эксцизия
Застройка бреши
(репликация)
MSH2/MSH3,
MSH2/
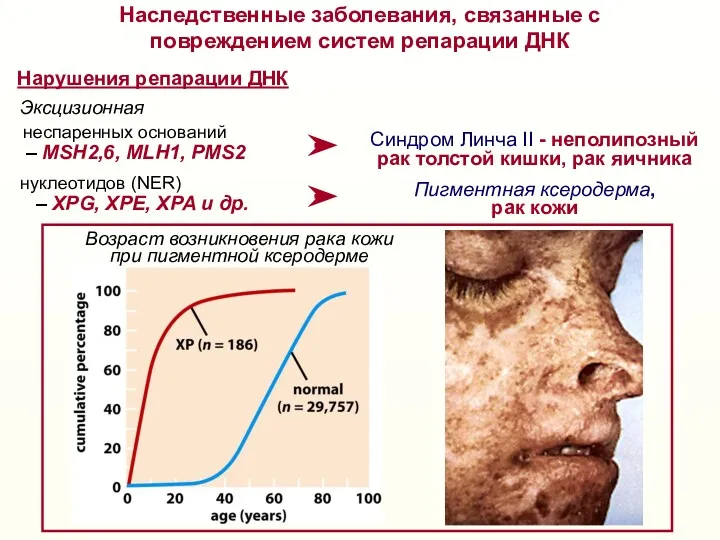
– MSH2,6, MLH1, PMS2
неспаренных оснований
Нарушения репарации ДНК
– XPG, XPE,
– MSH2,6, MLH1, PMS2
неспаренных оснований
Нарушения репарации ДНК
– XPG, XPE,

– NBS1, LIG4
Двунитевых разрывов
– FANCA, FANCG и др.
Межнитевых сшивок
Ниймегенский
– NBS1, LIG4
Двунитевых разрывов
– FANCA, FANCG и др.
Межнитевых сшивок
Ниймегенский
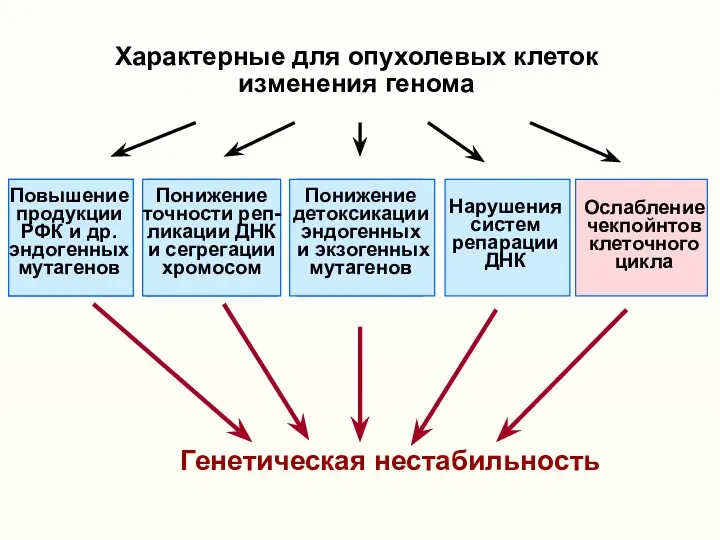
Характерные для опухолевых клеток
изменения генома
Генетическая нестабильность
Понижение
точности реп-
ликации ДНК
и сегрегации
Характерные для опухолевых клеток
изменения генома
Генетическая нестабильность
Понижение
точности реп-
ликации ДНК
и сегрегации
Повреждение ДНК
Белки-сенсоры
аномальных структур ДНК
Восстановление
структуры ДНК
Белки
репарационных
систем
Остановка
деления, апоптоз
Передатчики сигналов
(BRCA1,ATM/ATR и
Повреждение ДНК
Белки-сенсоры
аномальных структур ДНК
Восстановление
структуры ДНК
Белки
репарационных
систем
Остановка
деления, апоптоз
Передатчики сигналов
(BRCA1,ATM/ATR и
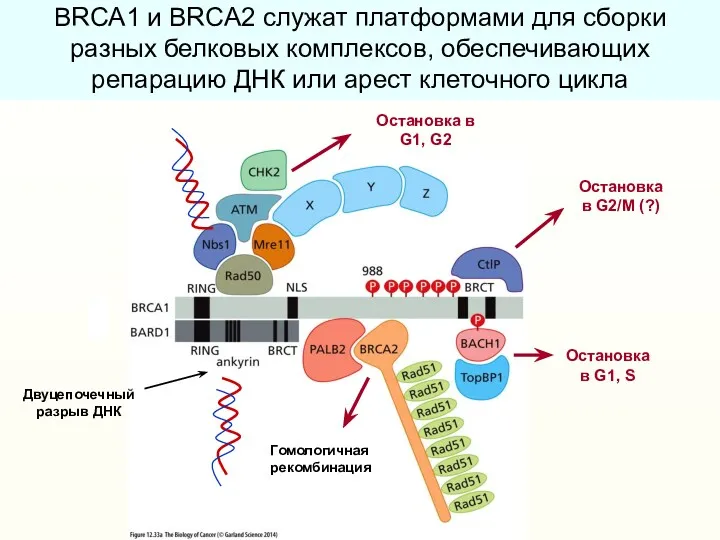
Гомологичная
рекомбинация
Остановка
в G2/M (?)
Остановка в G1, G2
Остановка в G1, S
Двуцепочечный
разрыв ДНК
BRCA1 и
Гомологичная
рекомбинация
Остановка
в G2/M (?)
Остановка в G1, G2
Остановка в G1, S
Двуцепочечный
разрыв ДНК
BRCA1 и
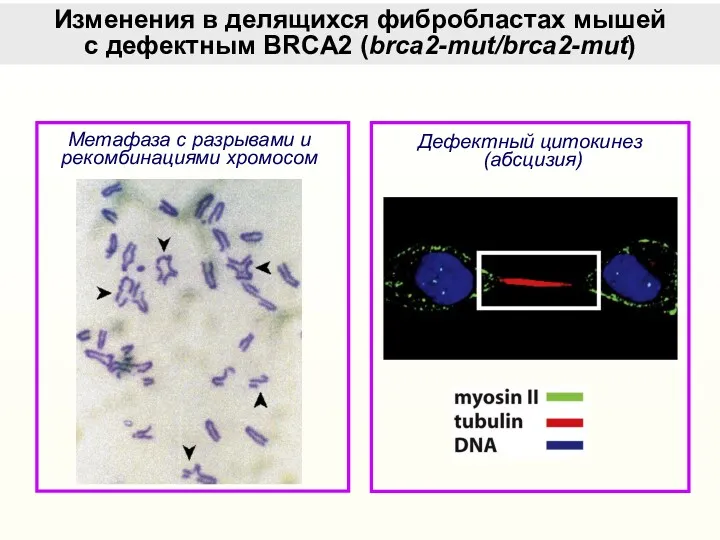
Изменения в делящихся фибробластах мышей
c дефектным BRCA2 (brca2-mut/brca2-mut)
Метафаза с разрывами
Изменения в делящихся фибробластах мышей
c дефектным BRCA2 (brca2-mut/brca2-mut)
Метафаза с разрывами
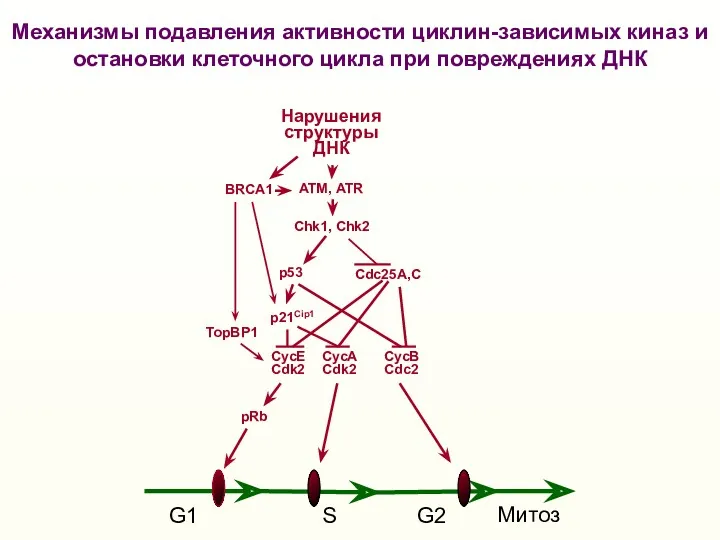
G1
S
G2
Митоз
Нарушения
структуры
ДНК
p53
Механизмы подавления активности циклин-зависимых киназ и остановки клеточного цикла при повреждениях
G1
S
G2
Митоз
Нарушения
структуры
ДНК
p53
Механизмы подавления активности циклин-зависимых киназ и остановки клеточного цикла при повреждениях

ATM
Aтаксия-телеангиоэктазия,
лимфолейкозы, лимфомы
BRCA1, BRCA2
Рак молочной железы, яичника
p53, CHK2
Синдром Ли-Фраумени –
различные новообразования
Сигнализация
ATM
Aтаксия-телеангиоэктазия,
лимфолейкозы, лимфомы
BRCA1, BRCA2
Рак молочной железы, яичника
p53, CHK2
Синдром Ли-Фраумени –
различные новообразования
Сигнализация
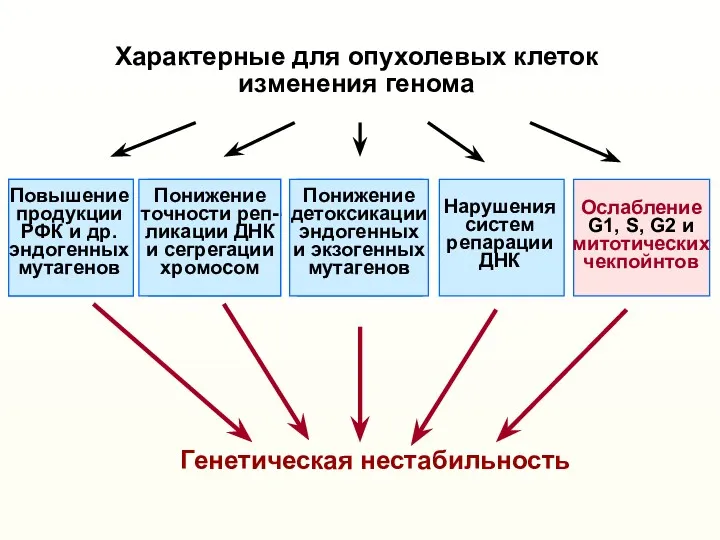
Характерные для опухолевых клеток
изменения генома
Генетическая нестабильность
Понижение
точности реп-
ликации ДНК
и сегрегации
Характерные для опухолевых клеток
изменения генома
Генетическая нестабильность
Понижение
точности реп-
ликации ДНК
и сегрегации
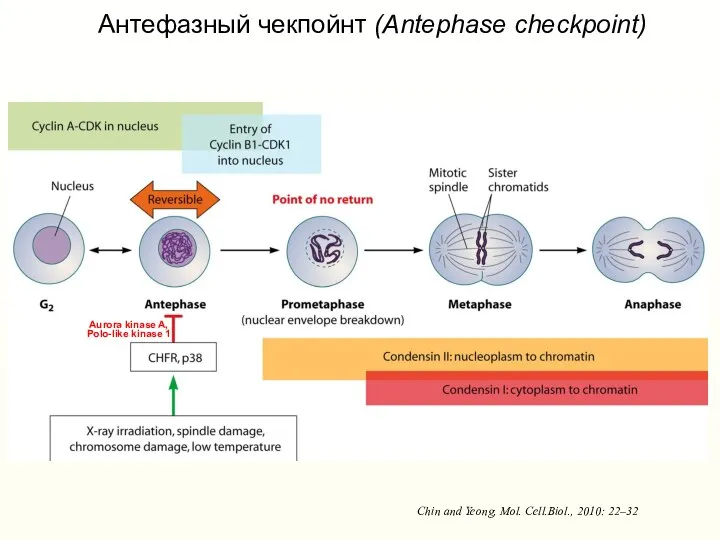
Chin and Yeong, Mol. Cell.Biol., 2010: 22–32
Антефазный чекпойнт (Antephase checkpoint)
Aurora kinase
Chin and Yeong, Mol. Cell.Biol., 2010: 22–32
Антефазный чекпойнт (Antephase checkpoint)
Aurora kinase
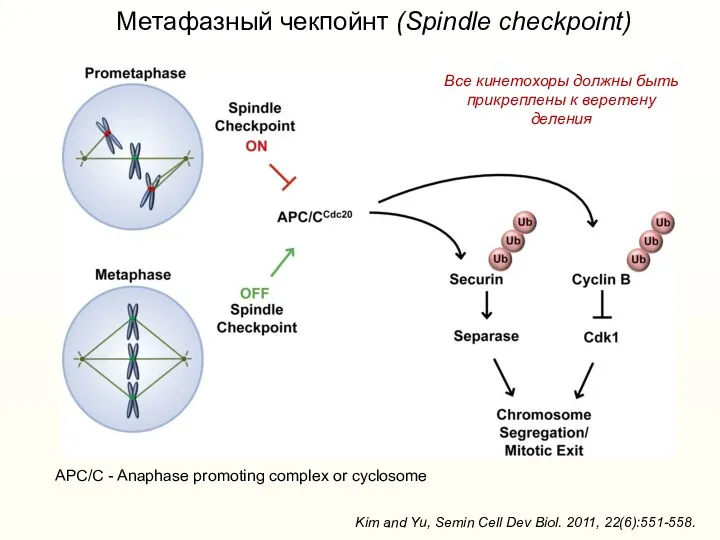
Метафазный чекпойнт (Spindle checkpoint)
Kim and Yu, Semin Cell Dev Biol. 2011,
Метафазный чекпойнт (Spindle checkpoint)
Kim and Yu, Semin Cell Dev Biol. 2011,
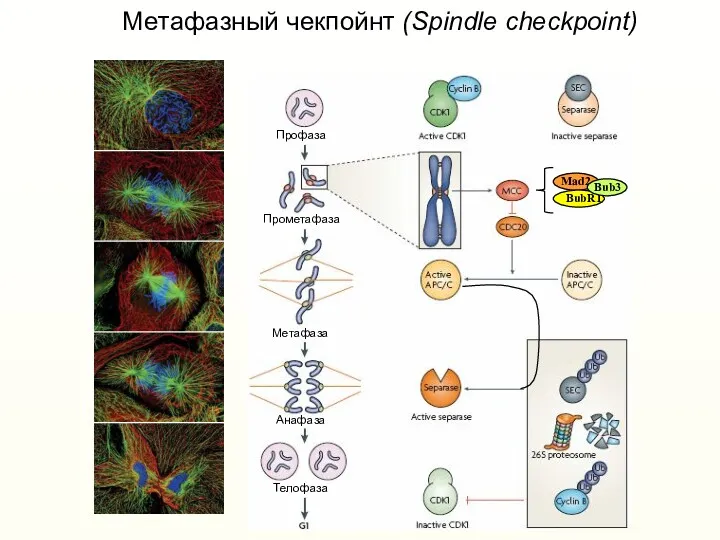
Метафазный чекпойнт (Spindle checkpoint)
Метафазный чекпойнт (Spindle checkpoint)
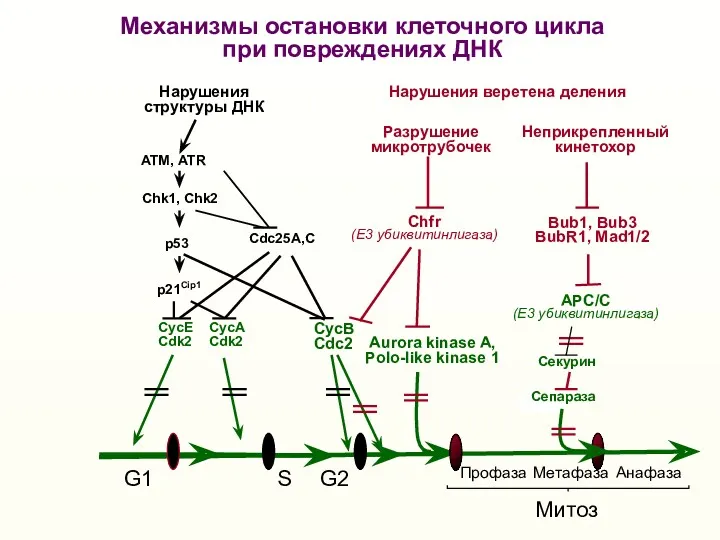
G1
S
G2
Митоз
Нарушения
структуры ДНК
Нарушения веретена деления
Bub1, Bub3
BubR1, Mad1/2
p53
p21Cip1
CycE
Cdk2
CycA
Cdk2
CycB
Cdc2
Cdc25A,C
ATM, ATR
Chk1, Chk2
Метафаза
Анафаза
Профаза
APC/C
(Е3 убиквитинлигаза)
Chfr
(Е3 убиквитинлигаза)
Неприкрепленный
кинетохор
Разрушение
микротрубочек
Aurora kinase
G1
S
G2
Митоз
Нарушения
структуры ДНК
Нарушения веретена деления
Bub1, Bub3
BubR1, Mad1/2
p53
p21Cip1
CycE
Cdk2
CycA
Cdk2
CycB
Cdc2
Cdc25A,C
ATM, ATR
Chk1, Chk2
Метафаза
Анафаза
Профаза
APC/C
(Е3 убиквитинлигаза)
Chfr
(Е3 убиквитинлигаза)
Неприкрепленный
кинетохор
Разрушение
микротрубочек
Aurora kinase

20-40% различных опухолей
(рак желудка, носоглотки и др.)
Гиперэкспрессия AURKA
Многие новообразования
Мутации MAD2
Bub1, Bub3,
20-40% различных опухолей
(рак желудка, носоглотки и др.)
Гиперэкспрессия AURKA
Многие новообразования
Мутации MAD2
Bub1, Bub3,
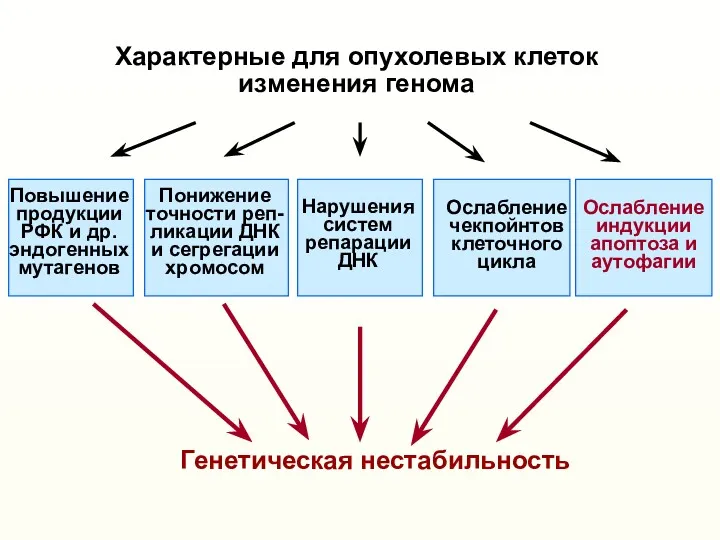
Характерные для опухолевых клеток
изменения генома
Понижение
точности реп-
ликации ДНК
и сегрегации
хромосом
Генетическая
Характерные для опухолевых клеток
изменения генома
Понижение
точности реп-
ликации ДНК
и сегрегации
хромосом
Генетическая
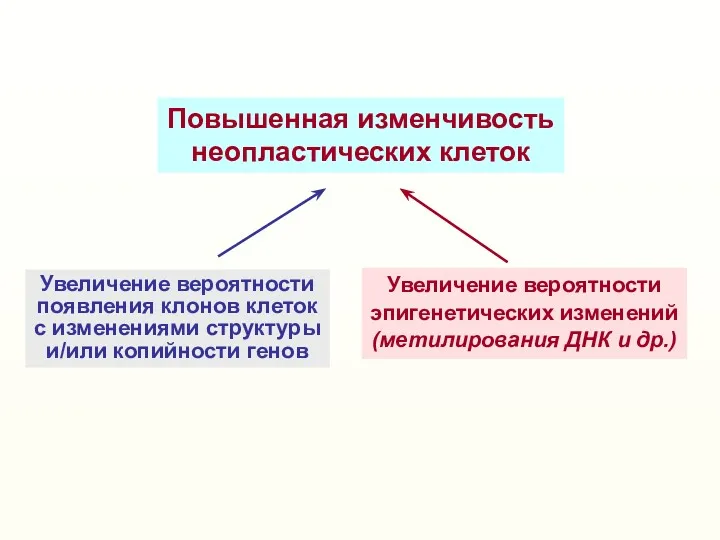
Повышенная изменчивость
неопластических клеток
Увеличение вероятности
появления клонов клеток
с изменениями структуры
и/или копийности генов
Увеличение
Повышенная изменчивость
неопластических клеток
Увеличение вероятности
появления клонов клеток
с изменениями структуры
и/или копийности генов
Увеличение
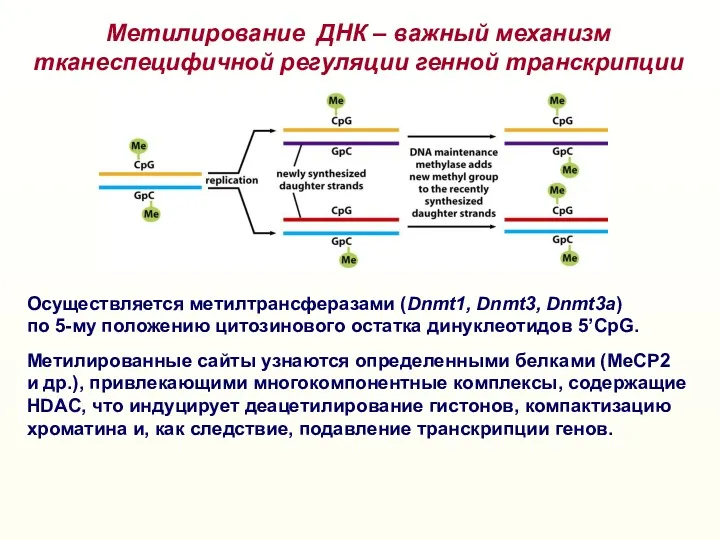
Метилирование ДНК – важный механизм тканеспецифичной регуляции генной транскрипции
Осуществляется метилтрансферазами (Dnmt1,
Метилирование ДНК – важный механизм тканеспецифичной регуляции генной транскрипции
Осуществляется метилтрансферазами (Dnmt1,
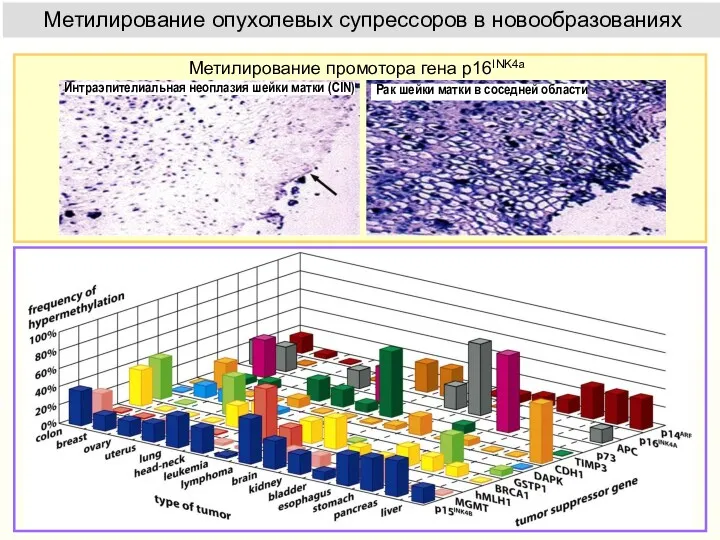
Метилирование опухолевых супрессоров в новообразованиях
Метилирование промотора гена p16INK4a
Метилирование опухолевых супрессоров в новообразованиях
Метилирование промотора гена p16INK4a
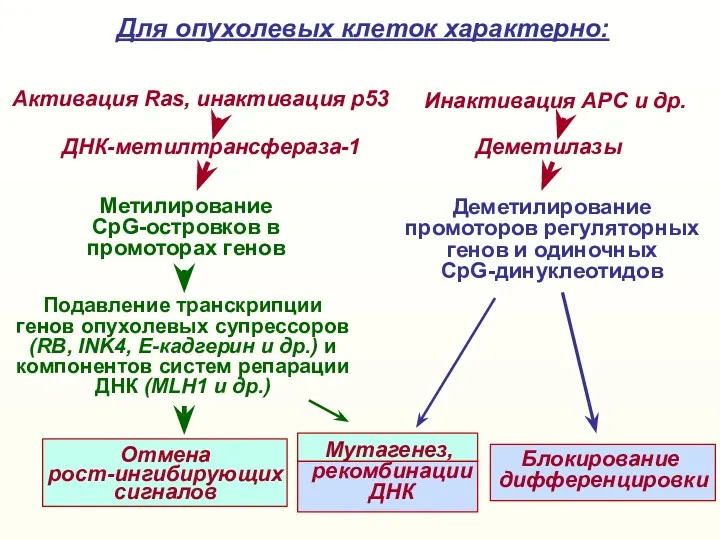
Метилирование
CpG-островков в
промоторах генов
Для опухолевых клеток характерно:
Деметилирование промоторов регуляторных генов и
Метилирование
CpG-островков в
промоторах генов
Для опухолевых клеток характерно:
Деметилирование промоторов регуляторных генов и
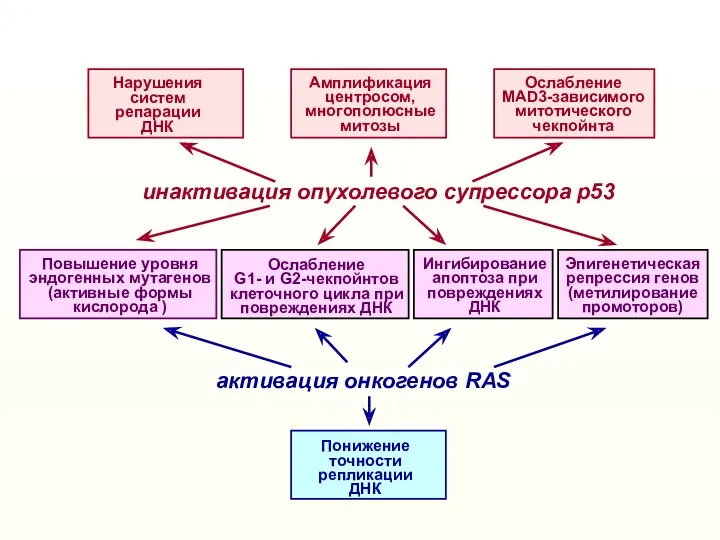
активация онкогенов RAS
инактивация опухолевого супрессора р53
Понижение
точности
репликации
ДНК
Повышение уровня
эндогенных мутагенов
(активные формы
кислорода )
Ослабление
G1-
активация онкогенов RAS
инактивация опухолевого супрессора р53
Понижение
точности
репликации
ДНК
Повышение уровня
эндогенных мутагенов
(активные формы
кислорода )
Ослабление
G1-
 Фармацевтическая биоэтика
Фармацевтическая биоэтика Профессиональное выгорание среди медицинских работников: связь с ошибками и качеством медицинской помощи
Профессиональное выгорание среди медицинских работников: связь с ошибками и качеством медицинской помощи Медицина народов Средневекового Востока (2)
Медицина народов Средневекового Востока (2) Infection. Forms of infection. Immunity. Types and forms of umminity. Factors specific and nonspecific defense
Infection. Forms of infection. Immunity. Types and forms of umminity. Factors specific and nonspecific defense Лечение в Израиле
Лечение в Израиле Синдром “супер жінки”
Синдром “супер жінки” Туберкулинодиагностика
Туберкулинодиагностика Лабораторная диагностика болезней желудочно-кишечного тракта
Лабораторная диагностика болезней желудочно-кишечного тракта Заболевания паращитовидных желез
Заболевания паращитовидных желез Витамин Д3
Витамин Д3 Влияние алкоголя на здоровье человека
Влияние алкоголя на здоровье человека Ультразвуковая диапевтика в урологии
Ультразвуковая диапевтика в урологии Дифференциальный диагноз заболеваний толстой кишки
Дифференциальный диагноз заболеваний толстой кишки Кишечные вирусные инфекции
Кишечные вирусные инфекции Тістердің жоғары қажалуының емі
Тістердің жоғары қажалуының емі Лучевая терапия и радиохирургия
Лучевая терапия и радиохирургия Вакцинопрофилактика
Вакцинопрофилактика Оказание первичной медицинской помощи при перфорации половых органов на уровне ПМСП
Оказание первичной медицинской помощи при перфорации половых органов на уровне ПМСП Догляд за хворими з хірургічними захворюваннями прямої кишки
Догляд за хворими з хірургічними захворюваннями прямої кишки Балалардағы Крон ауруы
Балалардағы Крон ауруы Клиническая фармакология диуретиков
Клиническая фармакология диуретиков Транквилизаторы
Транквилизаторы Гелиотерапия
Гелиотерапия Укрепление здоровья семьи, планирование семьи
Укрепление здоровья семьи, планирование семьи ҚРда және жақын шетелде медициналық
ҚРда және жақын шетелде медициналық Поражения слизистой оболочки полости рта при дерматозах
Поражения слизистой оболочки полости рта при дерматозах Тики (тикозные гиперкинезы)
Тики (тикозные гиперкинезы) Врачебный контроль в спортивной медицине
Врачебный контроль в спортивной медицине