- Лекция 19. Биологическая продуктивность биоценозов

Содержание
- 2. Биологическая продуктивность - свойство экосистемы создавать в процессе своего функционирования органическое (Биологическая) продукция – органическое вещество,
- 3. Все органическое вещество, создаваемое автотрофными организмами в процессах фотосинтеза или хемосинтеза, называется валовой первичной продукцией (ВПП).
- 4. Органическое вещество, создаваемое гетеротрофными организмами, находящимися на втором и последующих трофических уровнях, называется вторичной продукцией. В
- 5. Количество органического вещества, покидающего тем или иным способом экосистему (сбор урожая, вылов рыбы, вылет наземных насекомых
- 6. Общее количество живого вещества в экосистеме называется биомассой. Биомасса экосистемы образована всеми населяющими ее живыми организмами.
- 7. Биомасса является статическим показателем, который не имеет размерности времени. Напротив, продукция – динамический показатель, она является
- 8. Значения Р/B-коэффициентов рассчитываются для определенного периода времени, обычно за сутки, месяц, сезон вегетации, год и т.д..
- 9. Методы определения первичной продукции Чистая первичная продукция наземных экосистем обычно оценивают по годовому приросту биомассы растений.
- 10. Однако этот метод можно применить лишь к популяциям растений, в которых самоизреживание отсутствует либо незначительно. К
- 11. Основным методом определения первичной продукции в водных экосистемах является «метод светлых и темных склянок». Он был
- 12. Пример Начальное содержание кислорода в воде (контроль) равно 6 мгО2·л-1, в конце опыта в светлой склянке
- 13. В балансовых уравнениях фотосинтеза и дыхания соотношение молекулярной массы одной молекулы глюкозы и шести молекул кислорода
- 14. С другой стороны по величине первичной продукции растений, т.е. прироста органической массы растений (без учета воды
- 15. Радиоуглеродный метод определения первичной продукции фитоплнктона основан на том, что живые организмы в равных пропорциях поглощают
- 16. Калибровочная кривая, характеризующая скорость распада 14С.
- 17. Пример. В склянку с фитопланктоном внесли 100 мг смеси [(14CO2 + 12CO2], в которой соотношение Cr/Сs
- 18. Расчет первичной продукции по содержанию хлорофилла а в фитопланктоне. Для этого фитопланктон из известного объема воды
- 19. Пример. В водоеме взята проба воды объемом (V), равным 0,5 л. Фотосинтетические пигменты экстрагированы в 5
- 20. Для расчета первичной продукции по содержанию хлорофилла необходимо знать ассимиляционное число (АЧ). АЧ - отношение массы
- 21. Основные методы расчета продукция гетеротрофных организмов Выбор методов расчета вторичной продукции в значительной степени определяется особенностями
- 22. I тип. Популяции, состоящие из одновозрастных особей. К ним относятся популяции моноцикличных видов с непродолжительным (до
- 23. Эти виды значительную часть года проводят на стадии покоящихся яиц. Выход личинок из покоящихся яиц семян
- 24. Размерная структура таких популяций выражается одновершинной гистограммой с узким основанием. Ее модальный класс в течение вегетационного
- 25. II Тип. Популяции с дискретным распределением разных возрастных групп Характерны для животных и растений с длительным
- 26. Размеры тела, Доля в популяции, %% О+ 1+ 2+ 3+ 4+ и более Размерная структура популяции
- 27. III Тип. Популяции с непрерывным размножением в течение сезона вегетации. Подобный тип популяций характерен для планктонных
- 28. Динамика размерной структуры популяции Paramysis lacustris в Таганрогском заливе в первую половину сезона вегетации
- 29. IV Тип. Популяции с незначительными различиями размеров тела особей. К этому типу относятся популяции одноклеточных организмов
- 30. Продукция популяций второго типа может быть рассчитана по формуле: P = (W2 – W1)N = W2N
- 31. Таким образом, при отсутствии смертности продукция популяции (Р) за определенный период времени равна разности между конечной
- 32. С учетом смертности расчет продукции несколько усложнится: Р = Ве + В2 – В1, где Ве
- 33. Графическая иллюстрация расчета продукции гипотетической популяции второго типа по зависимости численности особей от их массы. Если
- 34. Таким образом, продукция популяции второго типа соответствует площади под кривой зависимости изменения численности особей в популяции
- 35. Продукция популяций первого типа может быть рассчитана способами, аналогичными приведенными для популяций второго типа. За весь
- 36. Допустим, снижение численности популяции первого типа (Nw) с увеличением средней массы ее особей (W) соответствует экспоненциальному
- 37. Графический пример расчета продукции гипотетической популяции первого типа по зависимости снижения численности когорты с увеличением средней
- 38. Биомасса элиминированных особей (Ве) численно равна площади под кривой экспоненциального уравнения). Ее легко рассчитать, взяв определенный
- 39. Пример. Функциональная связь между численностью когорты (N, экз·м-2) и средней массой ее особей (W, мг) следует
- 40. Продукция популяций третьего типа рассчитывается следующим образом. 1. Через определенные промежутки времени из природной популяции отбираются
- 41. Кривая роста и изменение абсолютного прироста особей с возрастом
- 42. Суммарный суточный прирост массы тела в одной размерной группы особей в популяции со средней массой Wi,
- 43. Суточная продукция одной размерной группы особей в популяции со средней массой Wi, равна: Умножив и разделив
- 44. Расчет суточной продукции популяции веслоногого рака Cyclоps colensis в озере Нарочь Исходя из данных таблицы, суточный
- 45. Продукцию популяции (ΣР) за периоды времени (t2 – t1), превышающие одни сутки, определяют согласно ΣР =
- 46. Продукции популяций четвертого типа. Продукция одноклеточных организмов, размножающихся делением надвое, за единицу времени определяется следующим образом:
- 47. Произведение NW представляет биомассу популяции (B). Тогда продукция за единицу времени, соответствующую размерности r (час, если
- 48. Пусть начальная биомасса популяции в момент времени t = 0 равна Во. Поскольку одноклеточные организмы размножаются
- 49. Физиологический метод расчета продукции Коловратки размножаются продуцированием яиц, число которых в кладке обычно больше одного. По
- 50. Легко доказать, что Р = Т У большинства видов значения К2 на организменном уровне снижаются с
- 52. Скачать презентацию

Биологическая продуктивность - свойство экосистемы создавать в процессе своего функционирования
Биологическая продуктивность - свойство экосистемы создавать в процессе своего функционирования

Все органическое вещество, создаваемое автотрофными организмами в процессах фотосинтеза или
Все органическое вещество, создаваемое автотрофными организмами в процессах фотосинтеза или

Органическое вещество, создаваемое гетеротрофными организмами, находящимися на втором и последующих
Органическое вещество, создаваемое гетеротрофными организмами, находящимися на втором и последующих

Количество органического вещества, покидающего тем или иным способом экосистему (сбор
Количество органического вещества, покидающего тем или иным способом экосистему (сбор

Общее количество живого вещества в экосистеме называется биомассой. Биомасса экосистемы
Общее количество живого вещества в экосистеме называется биомассой. Биомасса экосистемы

Биомасса является статическим показателем, который не имеет размерности времени.
Биомасса является статическим показателем, который не имеет размерности времени.

Значения Р/B-коэффициентов рассчитываются для определенного периода времени, обычно за сутки,
Значения Р/B-коэффициентов рассчитываются для определенного периода времени, обычно за сутки,

Методы определения первичной продукции
Чистая первичная продукция наземных экосистем обычно оценивают по
Методы определения первичной продукции
Чистая первичная продукция наземных экосистем обычно оценивают по

Однако этот метод можно применить лишь к популяциям растений,
Однако этот метод можно применить лишь к популяциям растений,

Основным методом определения первичной продукции в водных экосистемах является
Основным методом определения первичной продукции в водных экосистемах является

Пример
Начальное содержание кислорода в воде (контроль) равно 6
Пример
Начальное содержание кислорода в воде (контроль) равно 6

В балансовых уравнениях фотосинтеза и дыхания соотношение молекулярной массы одной
В балансовых уравнениях фотосинтеза и дыхания соотношение молекулярной массы одной

С другой стороны по величине первичной продукции растений, т.е. прироста
С другой стороны по величине первичной продукции растений, т.е. прироста

Радиоуглеродный метод определения первичной продукции фитоплнктона основан на том, что
Радиоуглеродный метод определения первичной продукции фитоплнктона основан на том, что
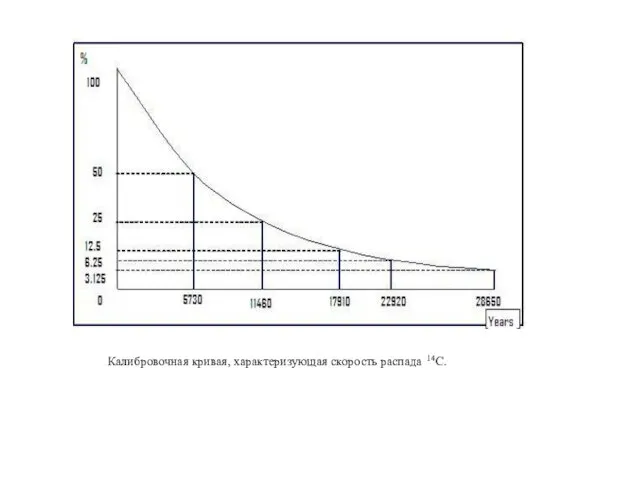
Калибровочная кривая, характеризующая скорость распада 14С.
Калибровочная кривая, характеризующая скорость распада 14С.

Пример.
В склянку с фитопланктоном внесли 100 мг смеси
Пример.
В склянку с фитопланктоном внесли 100 мг смеси

Расчет первичной продукции по содержанию хлорофилла а в фитопланктоне.
Расчет первичной продукции по содержанию хлорофилла а в фитопланктоне.

Пример.
В водоеме взята проба воды объемом (V), равным
Пример.
В водоеме взята проба воды объемом (V), равным

Для расчета первичной продукции по содержанию хлорофилла необходимо знать ассимиляционное число

Основные методы расчета продукция гетеротрофных организмов
Выбор методов расчета вторичной
Основные методы расчета продукция гетеротрофных организмов
Выбор методов расчета вторичной

I тип. Популяции, состоящие из одновозрастных особей.
К ним
I тип. Популяции, состоящие из одновозрастных особей.
К ним

Эти виды значительную часть года проводят на стадии покоящихся яиц.
Эти виды значительную часть года проводят на стадии покоящихся яиц.

Размерная структура таких популяций выражается одновершинной гистограммой с узким основанием.
Размерная структура таких популяций выражается одновершинной гистограммой с узким основанием.

II Тип. Популяции с дискретным распределением разных возрастных групп
II Тип. Популяции с дискретным распределением разных возрастных групп
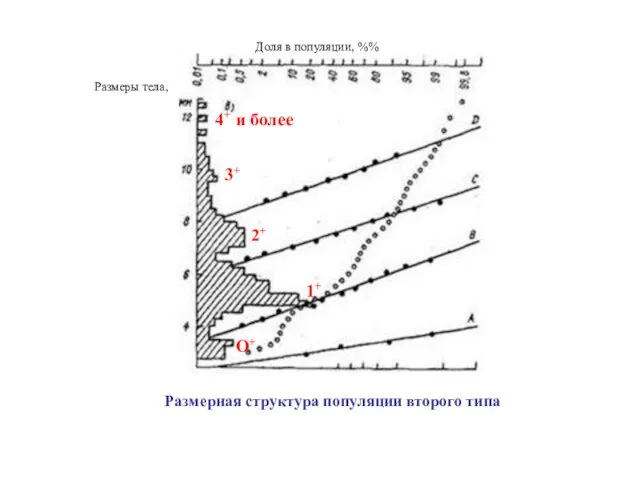
Размеры тела,
Доля в популяции, %%
О+
1+
2+
3+
4+ и более
Размерная структура популяции второго
Размеры тела,
Доля в популяции, %%
О+
1+
2+
3+
4+ и более
Размерная структура популяции второго

III Тип. Популяции с непрерывным размножением в течение сезона вегетации.
III Тип. Популяции с непрерывным размножением в течение сезона вегетации.

Динамика размерной структуры популяции Paramysis lacustris в Таганрогском заливе в первую
Динамика размерной структуры популяции Paramysis lacustris в Таганрогском заливе в первую
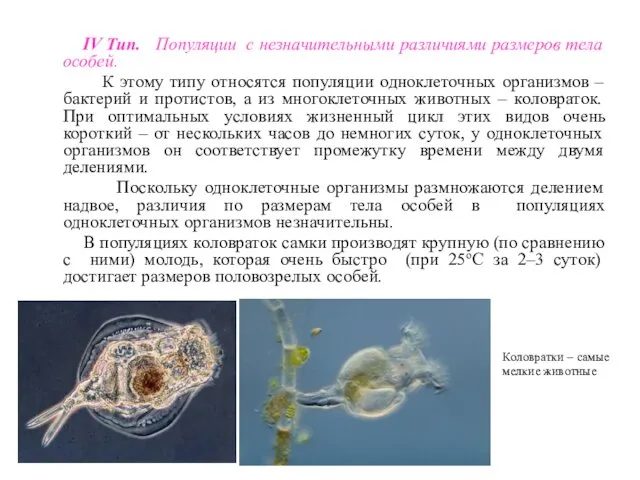
IV Тип. Популяции с незначительными различиями размеров тела особей.
IV Тип. Популяции с незначительными различиями размеров тела особей.

Продукция популяций второго типа может быть рассчитана по формуле:
P
Продукция популяций второго типа может быть рассчитана по формуле:
P

Таким образом, при отсутствии смертности продукция популяции (Р) за определенный
Таким образом, при отсутствии смертности продукция популяции (Р) за определенный

С учетом смертности расчет продукции несколько усложнится:
Р = Ве +
С учетом смертности расчет продукции несколько усложнится:
Р = Ве +
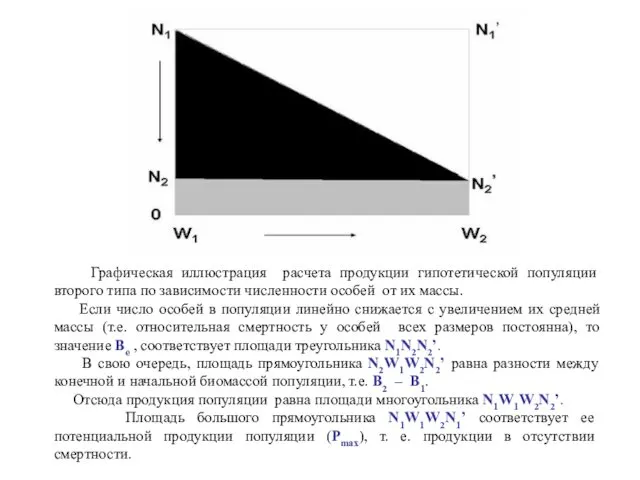
Графическая иллюстрация расчета продукции гипотетической популяции второго типа по зависимости
Графическая иллюстрация расчета продукции гипотетической популяции второго типа по зависимости

Таким образом, продукция популяции второго типа соответствует площади под кривой
Таким образом, продукция популяции второго типа соответствует площади под кривой

Продукция популяций первого типа может быть рассчитана способами, аналогичными приведенными
Продукция популяций первого типа может быть рассчитана способами, аналогичными приведенными

Допустим, снижение численности популяции первого типа (Nw) с увеличением средней
Допустим, снижение численности популяции первого типа (Nw) с увеличением средней
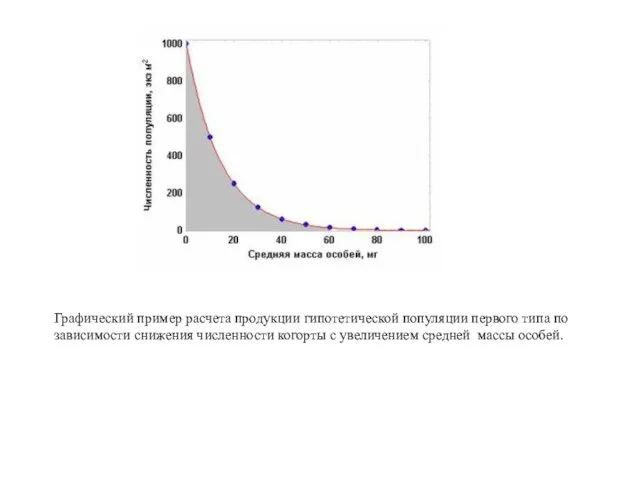
Графический пример расчета продукции гипотетической популяции первого типа по
зависимости снижения
Графический пример расчета продукции гипотетической популяции первого типа по
зависимости снижения

Биомасса элиминированных особей (Ве) численно равна площади под кривой экспоненциального
Биомасса элиминированных особей (Ве) численно равна площади под кривой экспоненциального

Пример.
Функциональная связь между численностью когорты (N, экз·м-2) и
Пример.
Функциональная связь между численностью когорты (N, экз·м-2) и

Продукция популяций третьего типа рассчитывается следующим образом.
1. Через
Продукция популяций третьего типа рассчитывается следующим образом.
1. Через
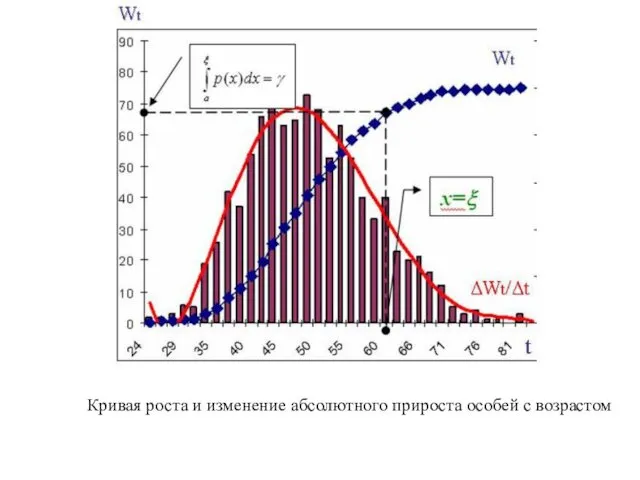
Кривая роста и изменение абсолютного прироста особей с возрастом
Кривая роста и изменение абсолютного прироста особей с возрастом

Суммарный суточный прирост массы тела в одной размерной группы особей
Суммарный суточный прирост массы тела в одной размерной группы особей

Суточная продукция одной размерной группы особей в популяции со средней
Суточная продукция одной размерной группы особей в популяции со средней
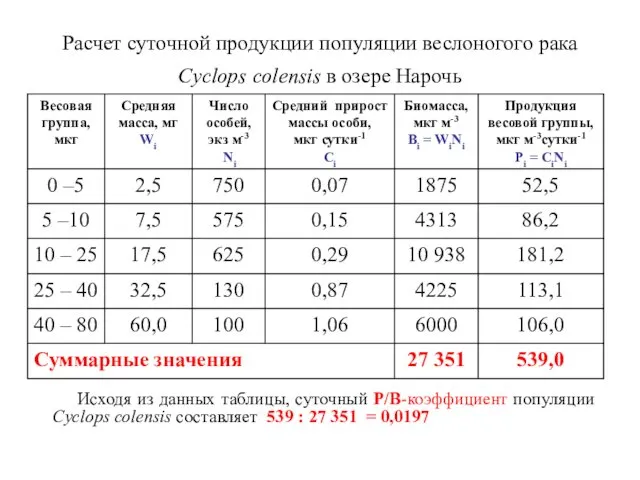
Расчет суточной продукции популяции веслоногого рака Cyclоps colensis в озере Нарочь
Расчет суточной продукции популяции веслоногого рака Cyclоps colensis в озере Нарочь

Продукцию популяции (ΣР) за периоды времени (t2 – t1),
Продукцию популяции (ΣР) за периоды времени (t2 – t1),

Продукции популяций четвертого типа.
Продукция одноклеточных организмов, размножающихся делением
Продукции популяций четвертого типа.
Продукция одноклеточных организмов, размножающихся делением

Произведение NW представляет биомассу популяции (B).
Тогда продукция за
Произведение NW представляет биомассу популяции (B).
Тогда продукция за

Пусть начальная биомасса популяции в момент времени t = 0
Пусть начальная биомасса популяции в момент времени t = 0

Физиологический метод расчета продукции
Коловратки размножаются продуцированием яиц, число
Физиологический метод расчета продукции
Коловратки размножаются продуцированием яиц, число

Легко доказать, что
Р = Т
У большинства видов значения К2 на
Легко доказать, что
Р = Т
У большинства видов значения К2 на
 Ко Дню защиты детей. Мастер-класс по изготовлению открытки
Ко Дню защиты детей. Мастер-класс по изготовлению открытки Многофункциональный scara манипулятор
Многофункциональный scara манипулятор Україна (укр. Україна [ukrɑˈjinɑ]) — держава у Східній Європі
Україна (укр. Україна [ukrɑˈjinɑ]) — держава у Східній Європі Основы малярных работ
Основы малярных работ презентация к уроку Генетическая связь.
презентация к уроку Генетическая связь. Етіологія та епідеміологія черевного тифу
Етіологія та епідеміологія черевного тифу Применение моделей для исследования изменений климата. Лекция 11
Применение моделей для исследования изменений климата. Лекция 11 Святой праведный Иоанн Кронштадтский
Святой праведный Иоанн Кронштадтский Estimation of permissible nitrogen content in the protective gas of the sodium circuit. Rosatom
Estimation of permissible nitrogen content in the protective gas of the sodium circuit. Rosatom Общий альбом визуализаций по интерьеру
Общий альбом визуализаций по интерьеру Кислотноcть и основность органических соединений. Инфракрасная спектроскопия. (Лекция 2)
Кислотноcть и основность органических соединений. Инфракрасная спектроскопия. (Лекция 2) Человеческие расы, их родство и происхождение
Человеческие расы, их родство и происхождение Презентация - Правила дорожного движения: ВНИМАНИЕ - СВЕТОФОР
Презентация - Правила дорожного движения: ВНИМАНИЕ - СВЕТОФОР Гигиена тела хирургического больного
Гигиена тела хирургического больного Методика Профессиональная готовность
Методика Профессиональная готовность История создания Вооруженных сил РФ
История создания Вооруженных сил РФ Дошкольник как субъект деятельности. Установление субъект субъектных отношений
Дошкольник как субъект деятельности. Установление субъект субъектных отношений Russian Wind Projects. Follow Up High Level Meeting
Russian Wind Projects. Follow Up High Level Meeting Презентация ЮНЕСКО
Презентация ЮНЕСКО Воспитательное мероприятие, посвященное 65-летию Великой победы Навеки в памяти людской.
Воспитательное мероприятие, посвященное 65-летию Великой победы Навеки в памяти людской. Физиология питания
Физиология питания Основания. 8класс. (презентация)
Основания. 8класс. (презентация) Развитие скоростно-силовых качеств и быстроты учащихся среднего школьного возраста
Развитие скоростно-силовых качеств и быстроты учащихся среднего школьного возраста презентация Чтобы помнили
презентация Чтобы помнили Непосредственно-образовательная деятельность в средней группе (Эксперимент с воздухом) с презентацией Злой и добрый ветер
Непосредственно-образовательная деятельность в средней группе (Эксперимент с воздухом) с презентацией Злой и добрый ветер Презентация Основные ошибки при создании презентации в программе PowerPoint
Презентация Основные ошибки при создании презентации в программе PowerPoint Самовыражение подростков
Самовыражение подростков Развитие театрального искусства в первой половине 19 в
Развитие театрального искусства в первой половине 19 в