- Моторизованный микроскоп. Съемка живых клеток

Содержание
- 2. Моторизованный микроскоп Высокопроизводительная микроскопия предполагает использование моторизованного микроскопа. В полностью моторизованном микроскопе следующие основные функции управляются
- 3. Моторизованный столик Управление по осям Х и У осуществляется с помощью сервомоторов (через джойстик). Точность перемещения
- 4. Работа моторизованного микроскопа Для съемки нескольких полей зрения (чашка Петри, многолуночный планшет) производится настройка фокусировки в
- 5. Установка постоянной фокусировки Две системы: 1. Через фокусировку камеры (максимальная резкость кадра). 2. Специальная система (perfect
- 6. Тестовый объект Кожица лука, окрашенная DiO C6(3) в концентрации около 0,5 мкг/мл (время окрашивания – 10-30
- 7. Фототоксичность и фотообесцвечивание Фотообесцвечивание и фототоксичность представляют собой два разных ограничения флуоресцентной микроскопии живых клеток. Фотообесцвечивание
- 8. Фототоксичность – формы АФК
- 9. Основные источники АФК в клетках животных Около 95 % от всего кислорода восстанавливается в митохондриях до
- 10. Основные мишени фотоповреждения в клетках 1. Повреждение ДНК и РНК. Повреждение ДНК сразу активирует систему репарации.
- 11. Фотопротекция клеток Для уменьшения фотообесцвечивания и снижения фототоксичности наиболее эффективным средством является фермент оксираза (Oxyrase Inc.).
- 12. Фотообесцвечивание Оксираза высокоэффективна при использовании низкомолекулярных органических красителей (Су-3, Су-5), но сравнительно малоэффективна при использовании флуоресцентных
- 13. Определение допустимой мощности излучения Эффект фототоксичности уменьшается с ростом длины волны возбуждающего света (Горгидзе, Ошемкова, Воробьев,
- 14. Приложения флуоресцентной микроскопии к исследованию живых клеток 1. FRAP (fluorescence recovery after photobleaching) – анализ динамики
- 15. FRAP и FLIP FRAP – динамические наблюдения внутри облученного района FRAP (одномоментное облучение) – кинетика восстановления
- 16. Методы FRAP и iFRAP
- 17. Пример – кинетика миозина III Example of a FRAP experiment to show that monomeric GFP can
- 18. Анализ методом FRAP - теория
- 19. Anatomy of a typical FRAP curve. (A) From the initial (pre-bleach) fluorescence intensity (Ii), the signal
- 20. Анализ методом FRAP - практика Для измерения желательно использовать четыре региона: (1) собственно ROI (красная линия),
- 21. Пример эксперимента iFRAP (A) Fluorescence images of a Cos7 cell micro-injected with pre-mRNAs before and after
- 22. Интерпретация данных, полученных методом FRAP Кривая FRAP содержит две компоненты: восстановление флуоресценции в результате диффузии молекул
- 23. FLIP – fluorescence loss in photobleaching Метод FLIP используется для анализа связи между органеллами, ядерно-цитоплазматического транспорта
- 24. Комбинирование FLIP и FRAP
- 25. FLIP и FRAP Пример для расчета FRAP и FLIP - экспрессия GFP с цитоплазматическим белком. FLIP
- 26. Приложение FLIP
- 28. Скачать презентацию

Моторизованный микроскоп
Высокопроизводительная микроскопия предполагает использование моторизованного микроскопа. В полностью моторизованном микроскопе
Моторизованный микроскоп
Высокопроизводительная микроскопия предполагает использование моторизованного микроскопа. В полностью моторизованном микроскопе

Моторизованный столик
Управление по осям Х и У осуществляется с помощью сервомоторов
Моторизованный столик
Управление по осям Х и У осуществляется с помощью сервомоторов

Работа моторизованного микроскопа
Для съемки нескольких полей зрения (чашка Петри, многолуночный планшет)
Работа моторизованного микроскопа
Для съемки нескольких полей зрения (чашка Петри, многолуночный планшет)

Установка постоянной фокусировки
Две системы:
1. Через фокусировку камеры (максимальная резкость кадра).
2.
Установка постоянной фокусировки
Две системы:
1. Через фокусировку камеры (максимальная резкость кадра).
2.

Тестовый объект
Кожица лука, окрашенная DiO C6(3) в концентрации около 0,5
Тестовый объект
Кожица лука, окрашенная DiO C6(3) в концентрации около 0,5

Фототоксичность и фотообесцвечивание
Фотообесцвечивание и фототоксичность представляют собой два разных ограничения флуоресцентной
Фототоксичность и фотообесцвечивание
Фотообесцвечивание и фототоксичность представляют собой два разных ограничения флуоресцентной
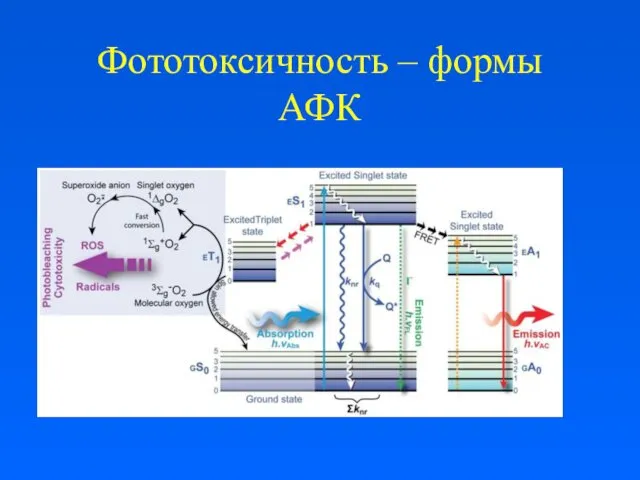
Фототоксичность – формы АФК
Фототоксичность – формы АФК
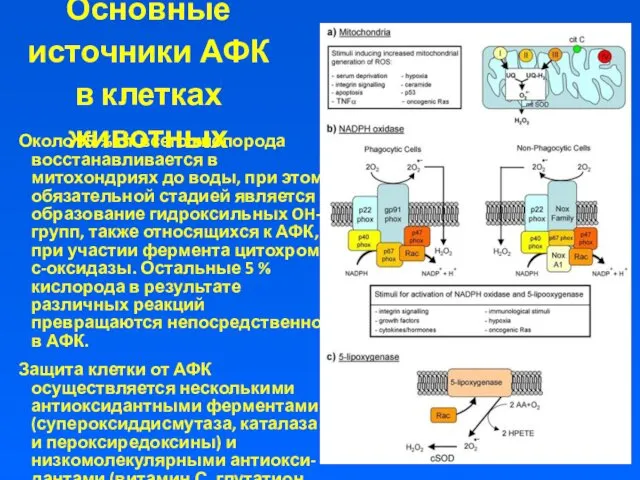
Основные источники АФК в клетках животных
Около 95 % от всего
Основные источники АФК в клетках животных
Около 95 % от всего

Основные мишени фотоповреждения в клетках
1. Повреждение ДНК и РНК. Повреждение ДНК
Основные мишени фотоповреждения в клетках
1. Повреждение ДНК и РНК. Повреждение ДНК

Фотопротекция клеток
Для уменьшения фотообесцвечивания и снижения фототоксичности наиболее эффективным средством является
Фотопротекция клеток
Для уменьшения фотообесцвечивания и снижения фототоксичности наиболее эффективным средством является

Фотообесцвечивание
Оксираза высокоэффективна при использовании низкомолекулярных органических красителей (Су-3, Су-5), но сравнительно
Фотообесцвечивание
Оксираза высокоэффективна при использовании низкомолекулярных органических красителей (Су-3, Су-5), но сравнительно

Определение допустимой мощности излучения
Эффект фототоксичности уменьшается с ростом длины волны возбуждающего
Определение допустимой мощности излучения
Эффект фототоксичности уменьшается с ростом длины волны возбуждающего

Приложения флуоресцентной микроскопии к исследованию живых клеток
1. FRAP (fluorescence recovery after
Приложения флуоресцентной микроскопии к исследованию живых клеток
1. FRAP (fluorescence recovery after

FRAP и FLIP
FRAP – динамические наблюдения внутри облученного района
FRAP (одномоментное
FRAP и FLIP
FRAP – динамические наблюдения внутри облученного района
FRAP (одномоментное
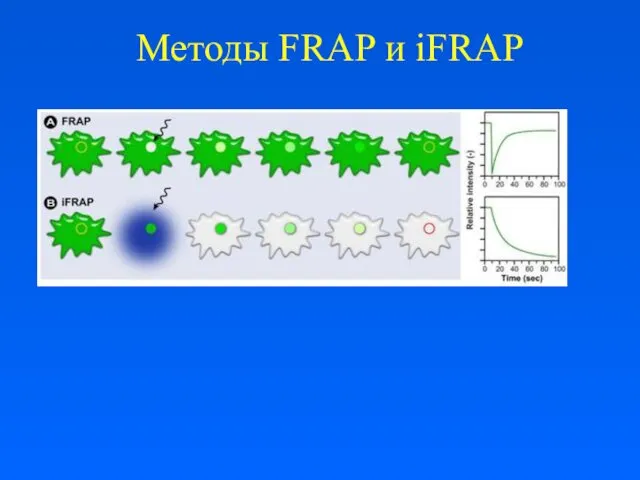
Методы FRAP и iFRAP
Методы FRAP и iFRAP
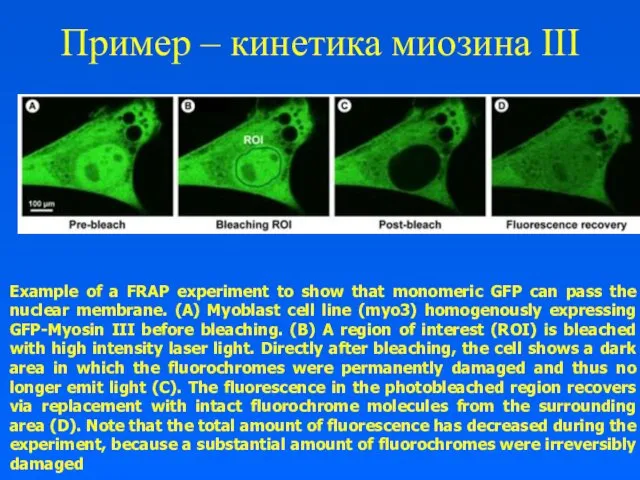
Пример – кинетика миозина III
Example of a FRAP experiment to show
Пример – кинетика миозина III
Example of a FRAP experiment to show
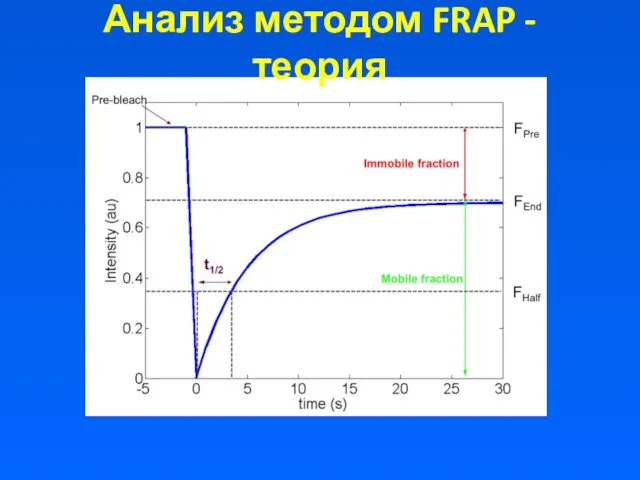
Анализ методом FRAP - теория
Анализ методом FRAP - теория
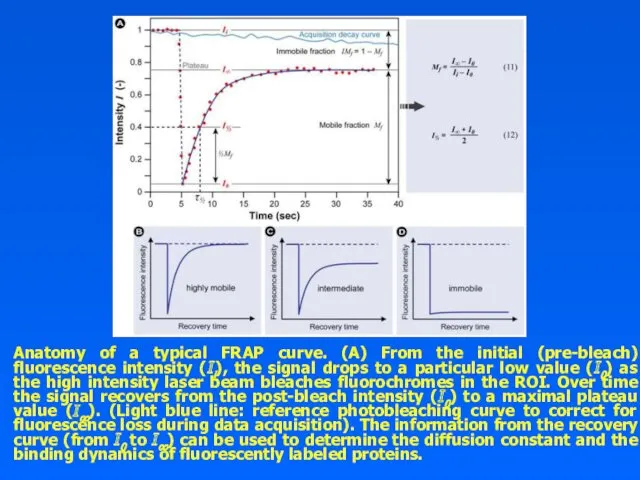
Anatomy of a typical FRAP curve. (A) From the initial (pre-bleach)
Anatomy of a typical FRAP curve. (A) From the initial (pre-bleach)
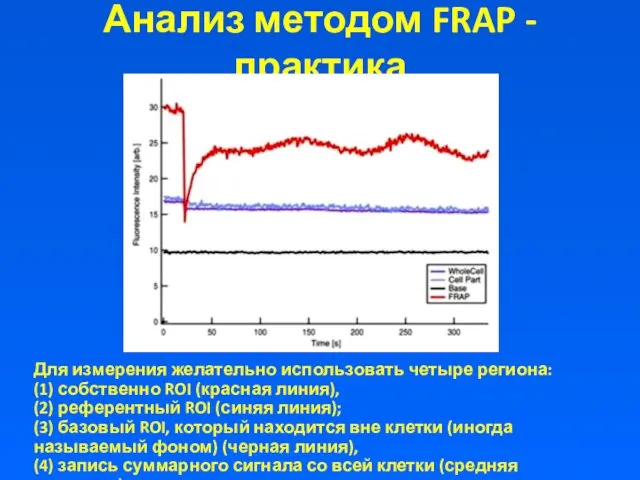
Анализ методом FRAP - практика
Для измерения желательно использовать четыре региона:
(1)
Анализ методом FRAP - практика
Для измерения желательно использовать четыре региона:
(1)
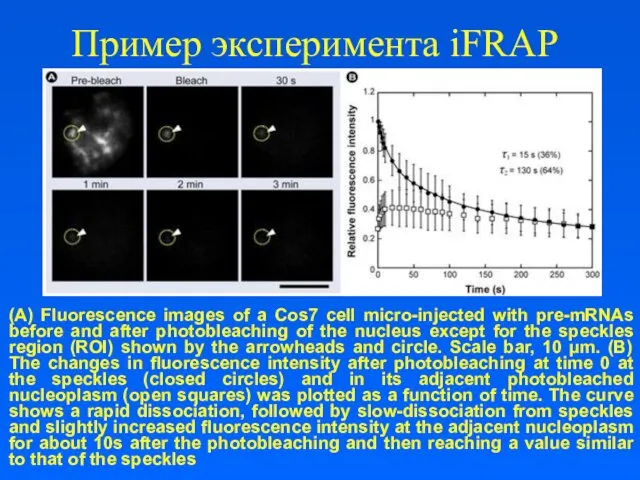
Пример эксперимента iFRAP
(A) Fluorescence images of a Cos7 cell micro-injected with
Пример эксперимента iFRAP
(A) Fluorescence images of a Cos7 cell micro-injected with

Интерпретация данных, полученных методом FRAP
Кривая FRAP содержит две компоненты: восстановление флуоресценции
Интерпретация данных, полученных методом FRAP
Кривая FRAP содержит две компоненты: восстановление флуоресценции
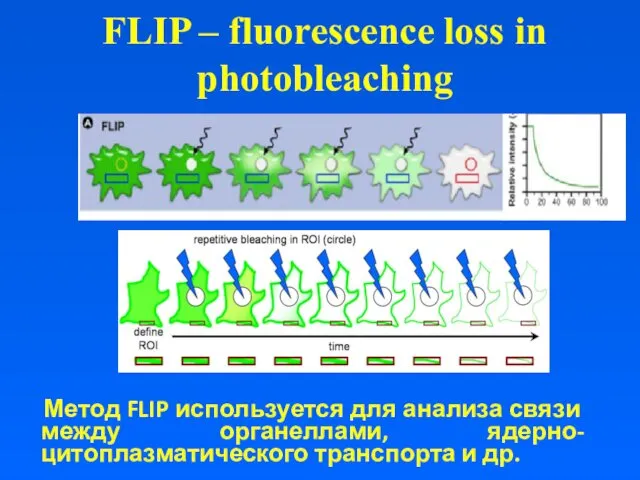
FLIP – fluorescence loss in photobleaching
Метод FLIP используется для анализа
FLIP – fluorescence loss in photobleaching
Метод FLIP используется для анализа
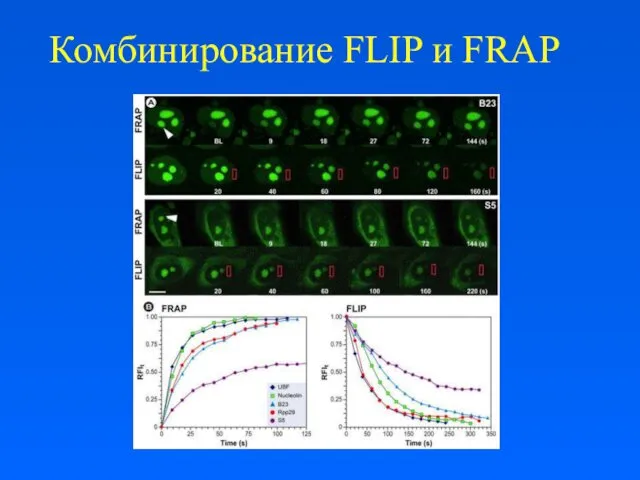
Комбинирование FLIP и FRAP
Комбинирование FLIP и FRAP
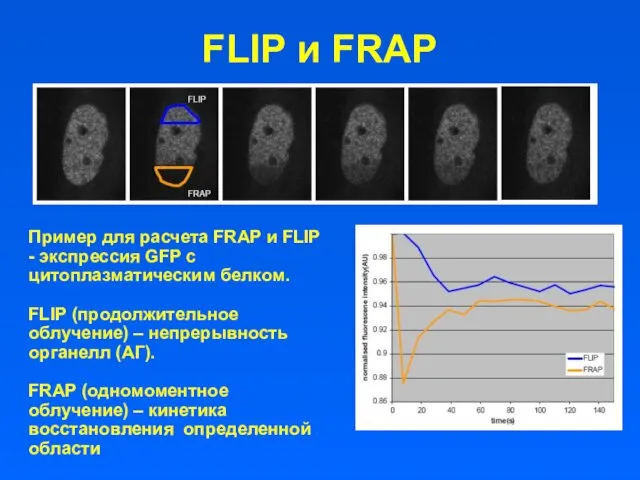
FLIP и FRAP
Пример для расчета FRAP и FLIP - экспрессия GFP
FLIP и FRAP
Пример для расчета FRAP и FLIP - экспрессия GFP
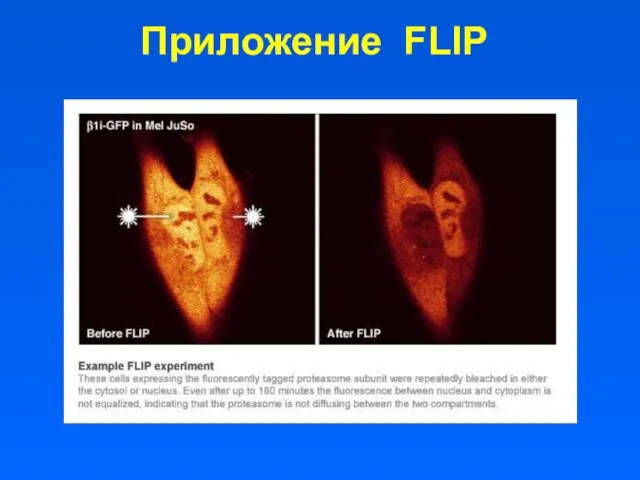
Приложение FLIP
Приложение FLIP
 Назначение, конструкция, технические характеристики, принцип действия, основные неисправности тележки
Назначение, конструкция, технические характеристики, принцип действия, основные неисправности тележки Модели простых сплошных сред
Модели простых сплошных сред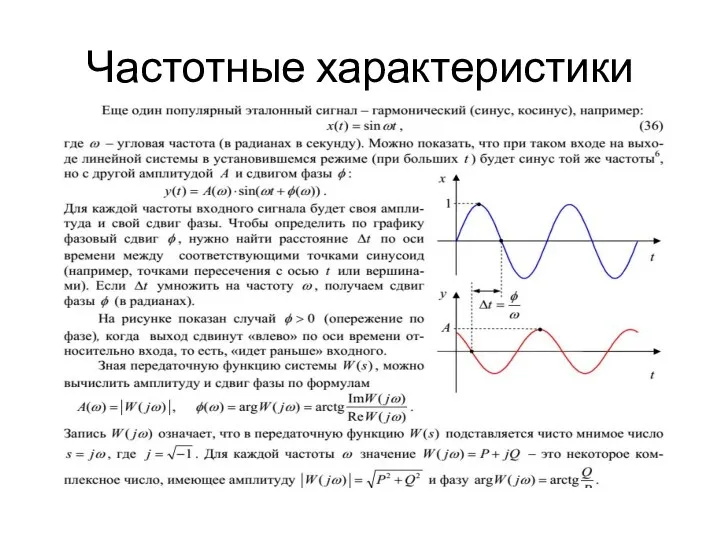 Частотные характеристики. ТАУ 3-1
Частотные характеристики. ТАУ 3-1 Уровень жидкости в сосудах
Уровень жидкости в сосудах Изучение электрической цепи.
Изучение электрической цепи. Что изучает физика. Междисциплинарные связи
Что изучает физика. Междисциплинарные связи Механізоване зварювання, наплавлення
Механізоване зварювання, наплавлення Разработка цифрового устройства для измерения технологических переменных электролизной лабораторной установки
Разработка цифрового устройства для измерения технологических переменных электролизной лабораторной установки Масс-спектрометрия в протеомных исследованиях. Часть 1: Масс-спектрометрия
Масс-спектрометрия в протеомных исследованиях. Часть 1: Масс-спектрометрия Опиливание металла. Размерная слесарная обработка
Опиливание металла. Размерная слесарная обработка Электромагнитная индукция. 11 класс
Электромагнитная индукция. 11 класс Резенке пластмассалар полихлоринилді оқшауламалы өткізгіштермен кабельдер
Резенке пластмассалар полихлоринилді оқшауламалы өткізгіштермен кабельдер Тушение пожара. Проект ORGLAG-01 (Мобильная платформа с системой огнетушения)
Тушение пожара. Проект ORGLAG-01 (Мобильная платформа с системой огнетушения) Абсолютная температура как мера средней кинетической энергии частиц
Абсолютная температура как мера средней кинетической энергии частиц Физика и музыка
Физика и музыка Уравнения Максвелла
Уравнения Максвелла Урок по теме Расчёт пути и времени движения 7 класс
Урок по теме Расчёт пути и времени движения 7 класс Сила Ампера
Сила Ампера Атмосферное давление. Вес воздуха
Атмосферное давление. Вес воздуха Явление тяготения. Сила тяжести
Явление тяготения. Сила тяжести КПП переднеприводного автомобиля
КПП переднеприводного автомобиля Основные положения. Механизм и машина
Основные положения. Механизм и машина Электронное пособие к обобщающему уроку физики в 8 классе по теме Магнитное поле.
Электронное пособие к обобщающему уроку физики в 8 классе по теме Магнитное поле. Презентация Давление жидкостей и газов для 7 класса
Презентация Давление жидкостей и газов для 7 класса Перспективы атомной энергетики. (Лекция 13)
Перспективы атомной энергетики. (Лекция 13) презентация к уроку физики по теме Кипение и конденсация
презентация к уроку физики по теме Кипение и конденсация Формирование учебно-познавательной компетенции учащихся второй ступени обучения на уроках физики
Формирование учебно-познавательной компетенции учащихся второй ступени обучения на уроках физики Кулачковые механизмы
Кулачковые механизмы