- Биомакромолекулы. Нуклеиновые кислоты

Содержание
- 2. Транскрипция Репликация
- 5. Происхождение ДНК
- 7. Нуклеиновые кислоты Химически нуклеиновые кислоты представляют собой биополимеры, состоящие из мономерных звеньев – нуклеотидов. Каждый нуклеотид
- 8. Азотистые основания Гетероциклические основания, входящие в состав нуклеиновых кислот (нуклеиновые основания), - это гидрокси- и аминопроизводные
- 9. Таутомерия азотистых оснований Все оксопроизводные азотистых оснований могут существовать в лактимной (енольной) и лактамной (кетонной) форме.
- 10. Минорные основания нуклеозидов Минорные нуклеозиды обнаружены практически во всех нуклеиновых кислотах. Наиболее высокое содержание минорных нуклеозидов
- 11. Минорные основания нуклеозидов К пуриновым минорным основаниям относятся такие: инозин, N6-метиладенин, N2- метилгуанин, ксантин, гипоксантин, 7-метилгуанин
- 12. Нуклеозиды Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой) Уридин Нуклеозиды лучше растворимы в воде,
- 13. Нуклеозиды. Углеводы. Углеводы (рибоза и дезоксирибоза ) в молекулах ДНК и РНК находятся в β-D-рибофуранозной форме:
- 14. Нуклеозиды Азотистые основания не участвуют в образовании никаких других ковалентных связей, помимо связывающей их с остатками
- 15. Гидролиз нуклеозидов аденозин рибоза аденин
- 16. Различные конформации сахара При этом нуклеотидная единица с 3′-эндоконформацией углеводного остатка имеет меньшую длину, чем 2′-эндоизомер.
- 17. Конформации
- 18. Конформации нуклеотидов Конформации основания угол вращения вокруг гликозидной связи C1’-N - ориентация основания относительно рибозы χ
- 19. Конформации рибозного кольца
- 20. Конформации Рибоза-основание Взаимодействие (-С5’H2- ) группы рибозы с основанием определяет область разрешенных значений Р, χ
- 21. Строение нуклеозида и нуклеотидов Нуклеотиды – фосфорные эфиры нуклеозидов.
- 22. Нуклеотиды Нуклеотиды - фосфорные эфиры нуклеозидов гликозидная связь сложноэфирная связь
- 23. Аденозин-5’-фосфат Аденозинмонофосфат (АМФ) 5’-адениловая кислота Гуанозин-5’-фосфат Гуанозинмонофосфат (ГМФ) 5’-гуаниловая кислота Номенклатура нуклеотидов
- 24. Нуклеотиды
- 25. Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам. У других нуклеотидов в сокращённом названии указывают положение
- 26. Нуклеотиды У рибонуклеотидов остаток фосфорной кислоты может находиться в положениях 2′ и 3′ и 5′, у
- 27. Конформации Фосфатная группа Три оптимальных состояния : (g-,g-), (g-,t), (t,g-)
- 28. Биохимические функции нуклеотидов 1) являются строительными блоками нуклеиновых кислот (ДНК и РНК); участвуют в молекулярных механизмах,
- 29. Модифицированные нуклеозиды Основное достоинство ацикловира заключается в том, что незараженные клетки человека не могут достаточно эффективно
- 30. цAMФ Циклические нуклеотиды Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль вторичного посредника
- 31. Нуклеиновые кислоты Нуклеиновые кислоты хорошо растворимы в воде, практически не растворимы в органических растворителях. Очень чувствительны
- 32. Гидролиз АТФ по стадиям
- 33. Нуклеиновые кислоты ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания: пуриновые — гуанин (G), аденин (A),пиримидиновые
- 34. Первичная структура ДНК и РНК
- 35. Первичная структура ДНК и РНК Первичная структура НК - последовательность нуклеотидов 5’- конец ТГАЦТААГТАЦЦ 3’-конец (ф-конец)
- 36. Гидролиз нуклеотидов АМФ аденозин рибоза аденин
- 37. Вторичная структура ДНК
- 38. Вторичная структура ДНК
- 39. Вторичная структура ДНК
- 40. Вторичная структура ДНК
- 41. Вторичная структура Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953) Две цепи антипараллельны друг
- 42. Вторичная структура ДНК Значительное количество доноров и акцепторов водородной связи атомов оснований и рибозы допускает большое
- 43. Образование связей между основаниями Каждое основание на одной из цепей связывается с одним определённым основанием на
- 44. Взаимодействия копланарных оснований
- 45. Канонические пары оснований – комплиментарные пары Энергии копланарных взаимодействий оснований для 20 конфигураций пар, образующих две
- 46. Не канонические пары оснований
- 47. Комплементарные пары нуклеиновых оснований Правила Чаргаффа Пра́вила Ча́ргаффа — система эмпирически выявленных правил, описывающих количественные соотношения
- 48. Стэкинг-взаимодействие Основания в цепи ДНК лежат друг над другом в стопке, что обеспечивает дополнительную стабилизацию цепи
- 49. Стэкинг-взаимодействие При этом оказывается, что стэкинг-взаимодействие между основаниями довольно специфично: полярные заместители одного основания, -NH2, =N-,
- 50. Вторичная структура ДНК Диаметр спирали постоянен вдоль всей её длины и равен 2,0 нм. Пуриновые и
- 51. Схематическое изображение струтуры молекулы ДНК 1.Малая борозда; 2.Большая борозда; 3.Углеводно-фосфатный остов; 4.Азотистые основания; 5.Водородные связи между
- 52. Вторичная структура ДНК
- 53. Спиральные параметры
- 54. Регулярные формы двойной спирали: А, В, Z Молекулы ДНК могут находиться в различных конформационных со- стояниях
- 55. Формы двойной спирали: А, В, Z
- 56. А-форма двойной спирали Правозакрученные спирали образуют два семейства: А-семейство и В-семейство. А-семейство ДНК представлено так называемой
- 57. В-форма двойной спирали Для В-форм ДНК характерно структурное разнообразие. Формы ДНК со случайными последовательностями могут находиться
- 58. Формы двойной спирали: А, В, Z
- 59. Z-форма ДНК Z-форма ДНК имеет левозакрученную спираль и обнаружена у полинуклеотида с чередующейся последовательностью (dG-dC). Особенность
- 60. Z-форма ДНК Область перехода перемещается вдоль спирали в виде небольшой петли. Переход В-формы ДНК в Z-форму
- 62. Регулярные формы двойной спирали: А, В, Z Значения спиральных параметров и структурные особенности
- 63. Средние значения конформационных параметров форм ДНК Средние значения конформационных параметров рибозо-фосфатного остова Средние значения спиральных параметров
- 64. Конформации нуклеотидов наблюдаемые в кристаллах ДНК дуплексов
- 65. Зависимость стабильности двойной спирали от контекстного состава
- 66. Сверхспирализация ДНК. (Суперскрученность) Если взяться за концы верёвки и начать скручивать их в разные стороны, она
- 67. Физико-химические свойства ДНК ДНК – довольно сильная многоосновная кислота, полностью ионизированная при рН 4,0. Фосфатные группы
- 68. Физико-химические свойства ДНК У большинства вирусов ДНК представляет собой двойную спираль, линейную или замкнутую в кольцо.
- 69. Спасибо за внимание!
- 76. В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует
- 77. Нуклеиновые кислоты
- 79. Скачать презентацию

Транскрипция
Репликация
Транскрипция
Репликация


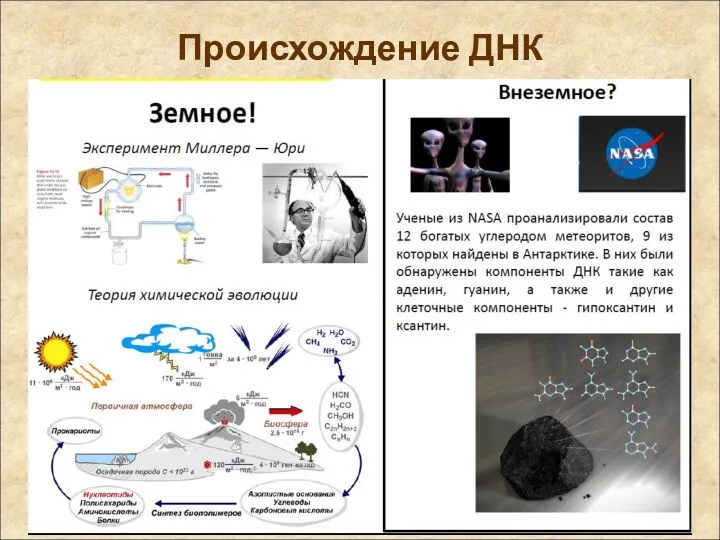
Происхождение ДНК
Происхождение ДНК
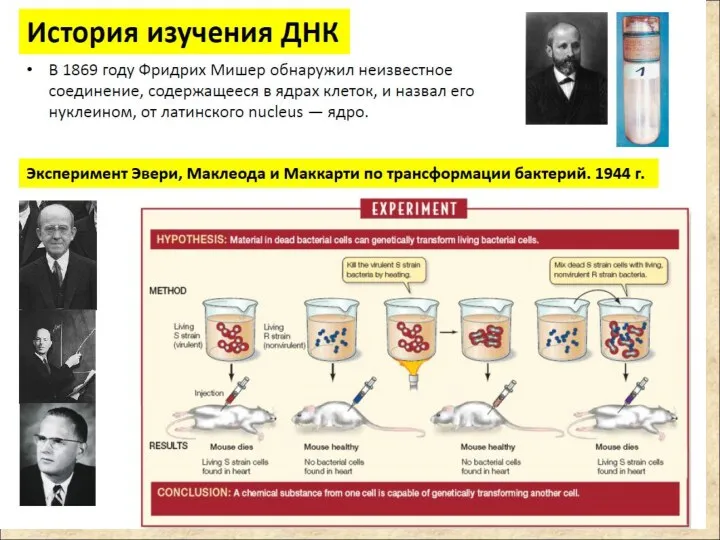
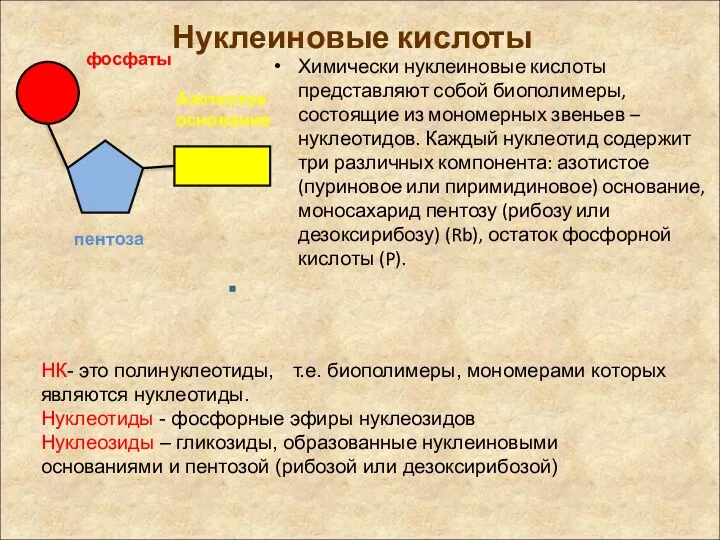
Нуклеиновые кислоты
Химически нуклеиновые кислоты представляют собой биополимеры, состоящие из мономерных звеньев
Нуклеиновые кислоты
Химически нуклеиновые кислоты представляют собой биополимеры, состоящие из мономерных звеньев
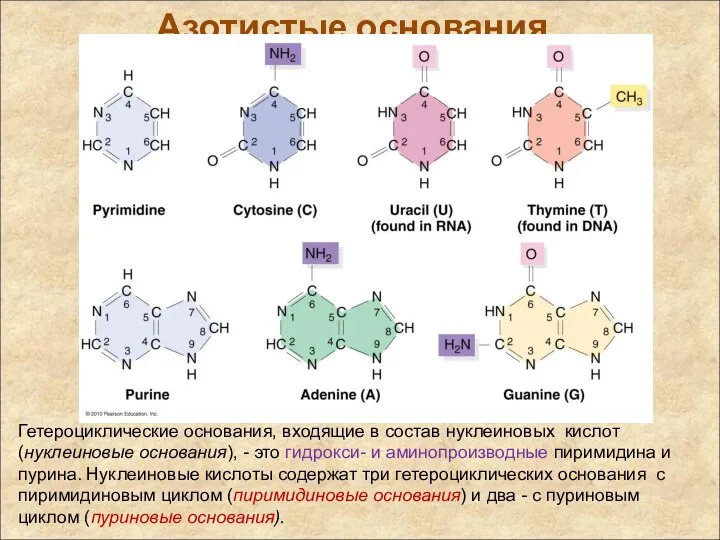
Азотистые основания
Гетероциклические основания, входящие в состав нуклеиновых кислот (нуклеиновые основания), -
Азотистые основания
Гетероциклические основания, входящие в состав нуклеиновых кислот (нуклеиновые основания), -
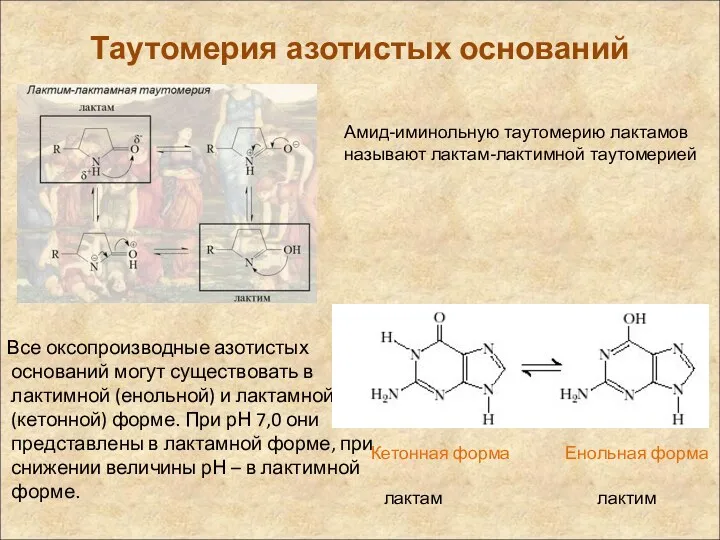
Таутомерия азотистых оснований
Все оксопроизводные азотистых оснований могут существовать в лактимной
Таутомерия азотистых оснований
Все оксопроизводные азотистых оснований могут существовать в лактимной
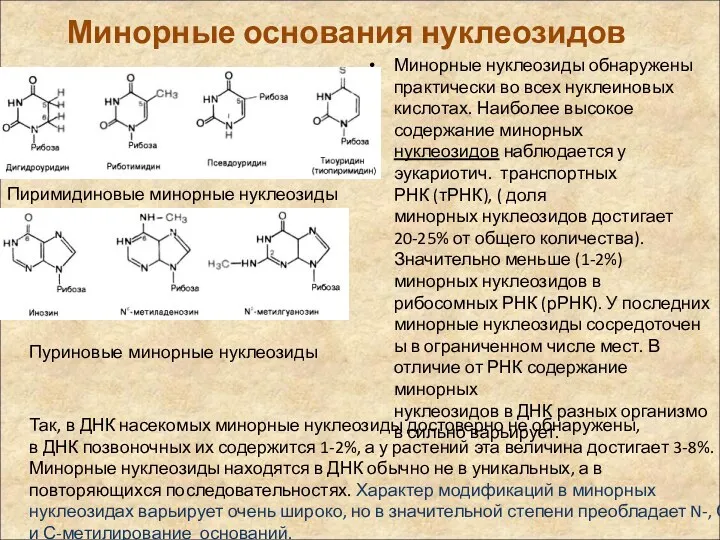
Минорные основания нуклеозидов
Минорные нуклеозиды обнаружены практически во всех нуклеиновых кислотах. Наиболее высокое
Минорные основания нуклеозидов
Минорные нуклеозиды обнаружены практически во всех нуклеиновых кислотах. Наиболее высокое

Минорные основания нуклеозидов
К пуриновым минорным основаниям относятся такие: инозин, N6-метиладенин,
Минорные основания нуклеозидов
К пуриновым минорным основаниям относятся такие: инозин, N6-метиладенин,
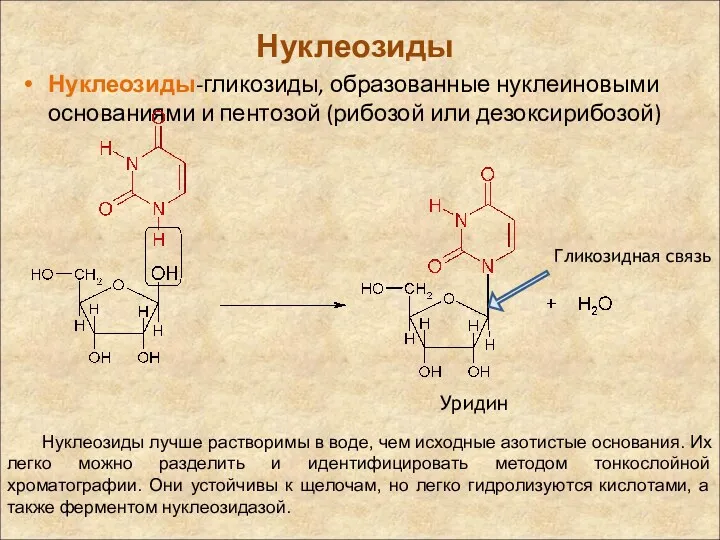
Нуклеозиды
Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой)
Уридин
Нуклеозиды лучше растворимы
Нуклеозиды
Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой)
Уридин
Нуклеозиды лучше растворимы
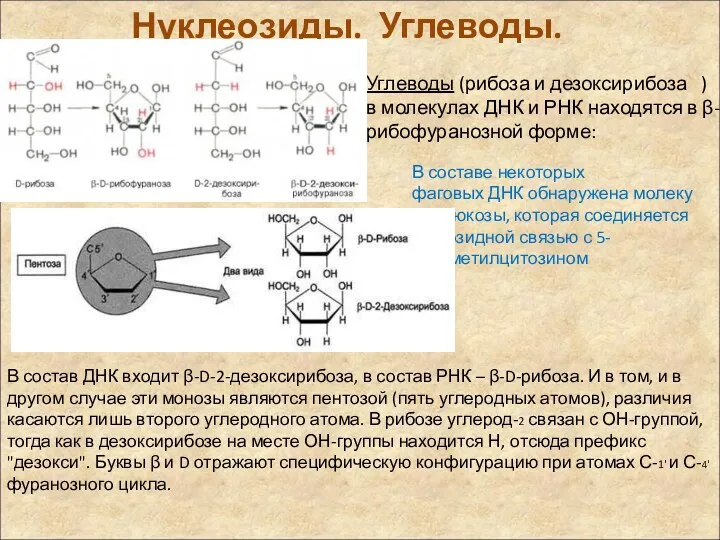
Нуклеозиды. Углеводы.
Углеводы (рибоза и дезоксирибоза ) в молекулах ДНК и РНК находятся в β-D-рибофуранозной форме:
В составе некоторых фаговых ДНК обнаружена молекула глюкозы, которая
Нуклеозиды. Углеводы.
Углеводы (рибоза и дезоксирибоза ) в молекулах ДНК и РНК находятся в β-D-рибофуранозной форме:
В составе некоторых фаговых ДНК обнаружена молекула глюкозы, которая
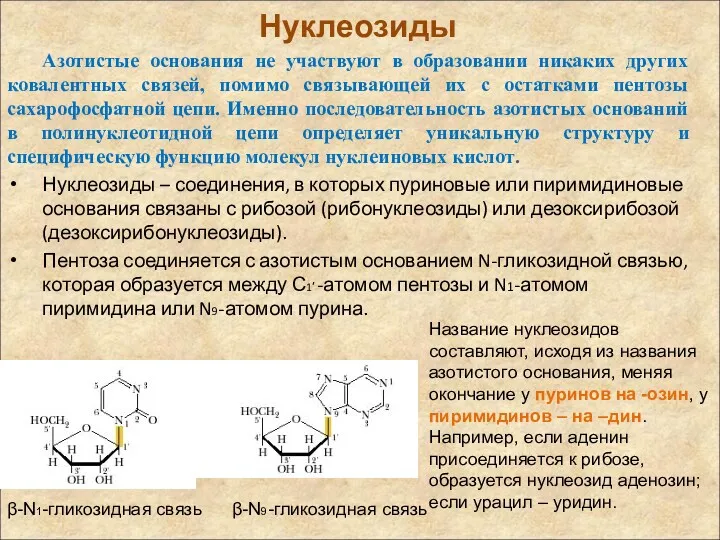
Нуклеозиды
Азотистые основания не участвуют в образовании никаких других ковалентных связей, помимо
Нуклеозиды
Азотистые основания не участвуют в образовании никаких других ковалентных связей, помимо
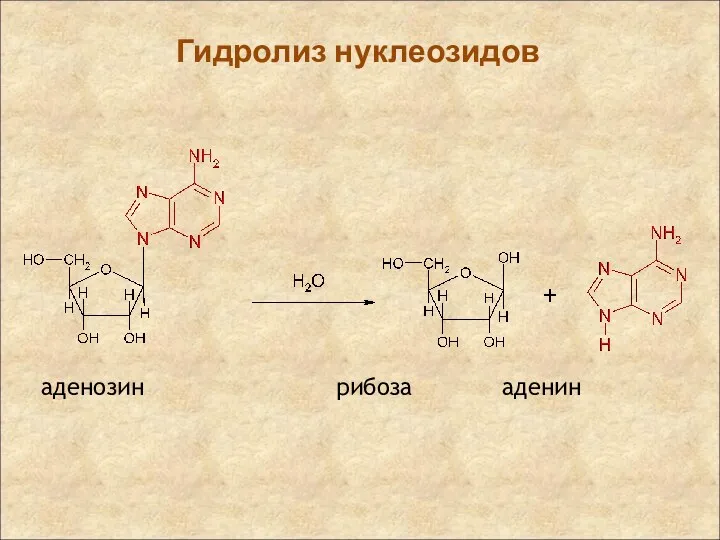
Гидролиз нуклеозидов
аденозин рибоза аденин
Гидролиз нуклеозидов
аденозин рибоза аденин

Различные конформации сахара
При этом нуклеотидная единица с 3′-эндоконформацией углеводного остатка имеет
Различные конформации сахара
При этом нуклеотидная единица с 3′-эндоконформацией углеводного остатка имеет
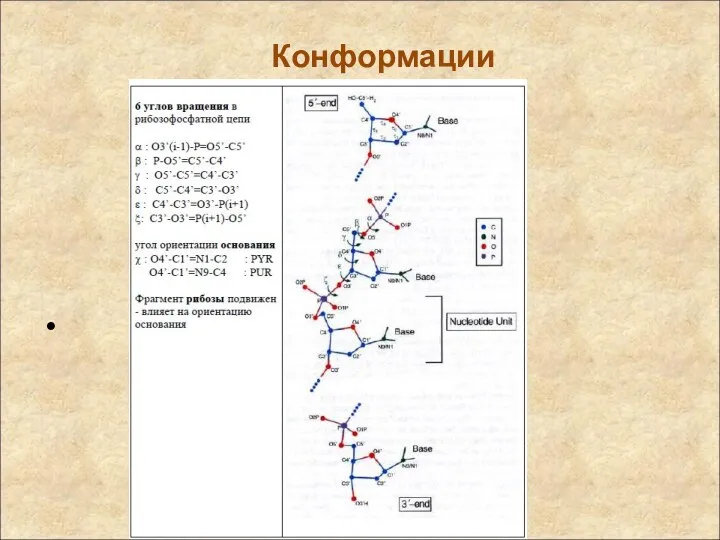
Конформации
Конформации
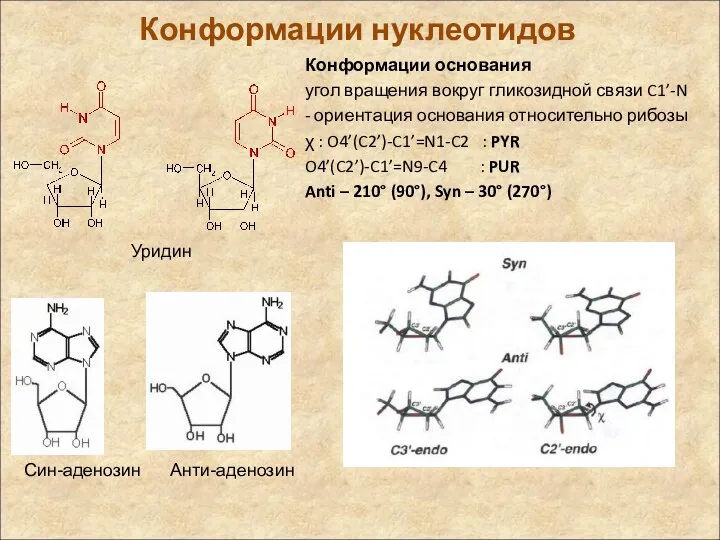
Конформации нуклеотидов
Конформации основания
угол вращения вокруг гликозидной связи C1’-N
- ориентация основания относительно
Конформации нуклеотидов
Конформации основания
угол вращения вокруг гликозидной связи C1’-N
- ориентация основания относительно

Конформации рибозного кольца
Конформации рибозного кольца
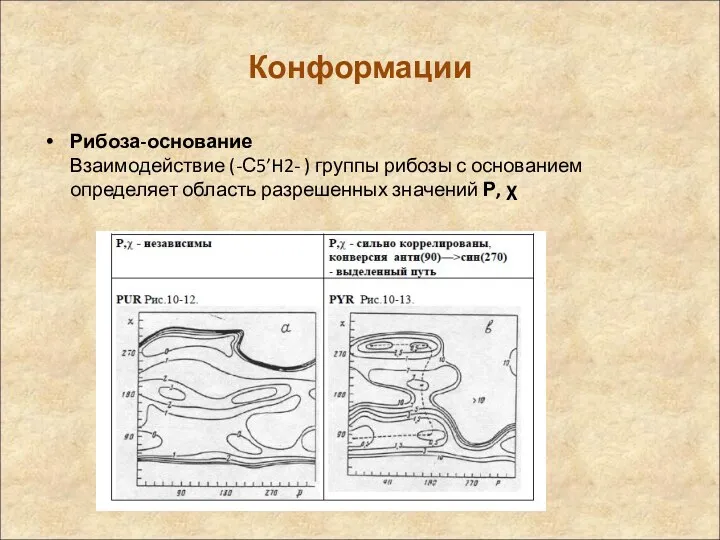
Конформации
Рибоза-основание
Взаимодействие (-С5’H2- ) группы рибозы с основанием определяет область разрешенных значений
Конформации
Рибоза-основание Взаимодействие (-С5’H2- ) группы рибозы с основанием определяет область разрешенных значений
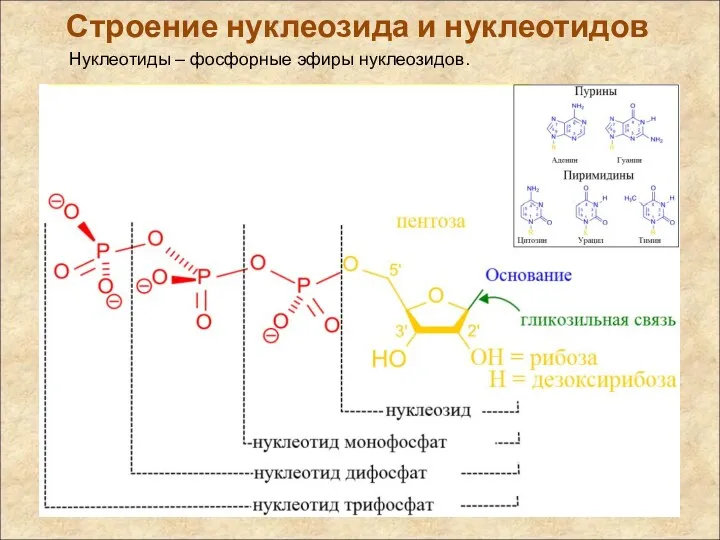
Строение нуклеозида и нуклеотидов
Нуклеотиды – фосфорные эфиры нуклеозидов.
Строение нуклеозида и нуклеотидов
Нуклеотиды – фосфорные эфиры нуклеозидов.
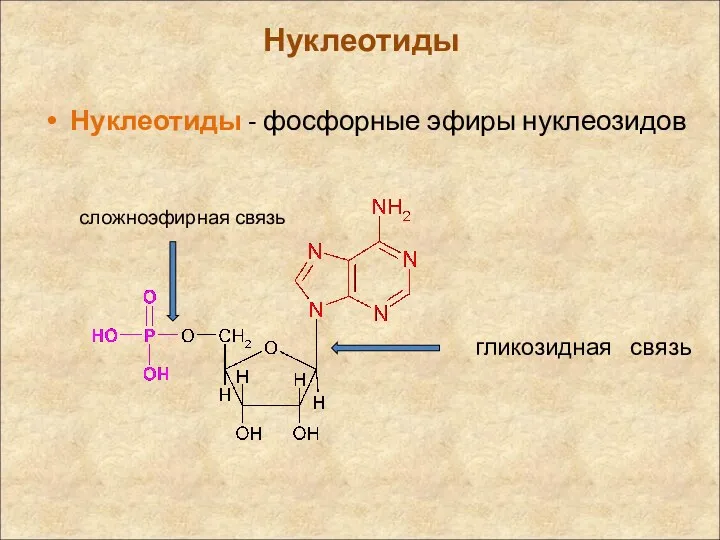
Нуклеотиды
Нуклеотиды - фосфорные эфиры нуклеозидов
гликозидная связь
сложноэфирная связь
Нуклеотиды
Нуклеотиды - фосфорные эфиры нуклеозидов
гликозидная связь
сложноэфирная связь
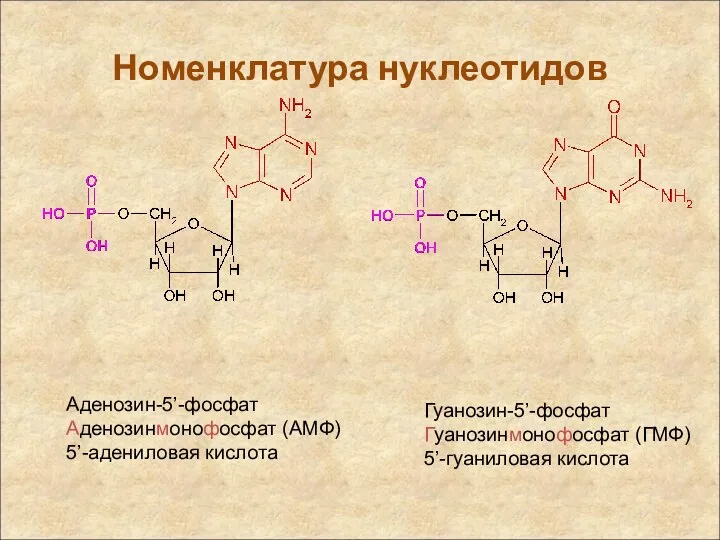
Аденозин-5’-фосфат
Аденозинмонофосфат (АМФ)
5’-адениловая кислота
Гуанозин-5’-фосфат
Гуанозинмонофосфат (ГМФ)
5’-гуаниловая кислота
Номенклатура нуклеотидов
Аденозин-5’-фосфат
Аденозинмонофосфат (АМФ)
5’-адениловая кислота
Гуанозин-5’-фосфат
Гуанозинмонофосфат (ГМФ)
5’-гуаниловая кислота
Номенклатура нуклеотидов

Нуклеотиды
Нуклеотиды
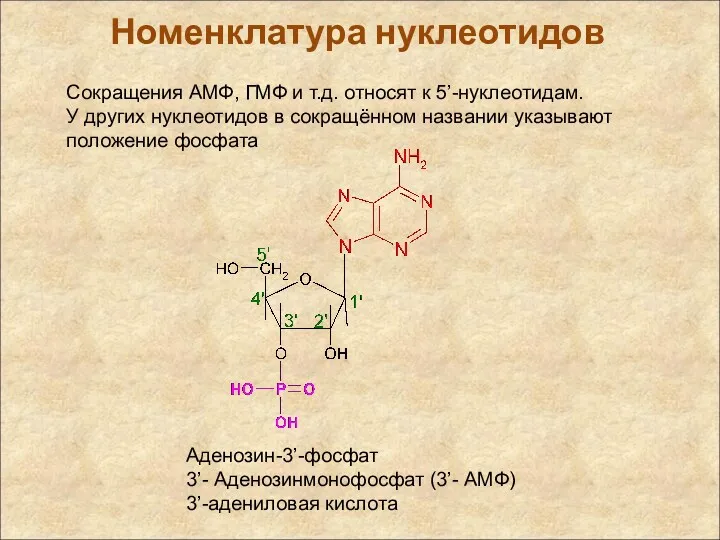
Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам.
У других нуклеотидов в
Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам.
У других нуклеотидов в
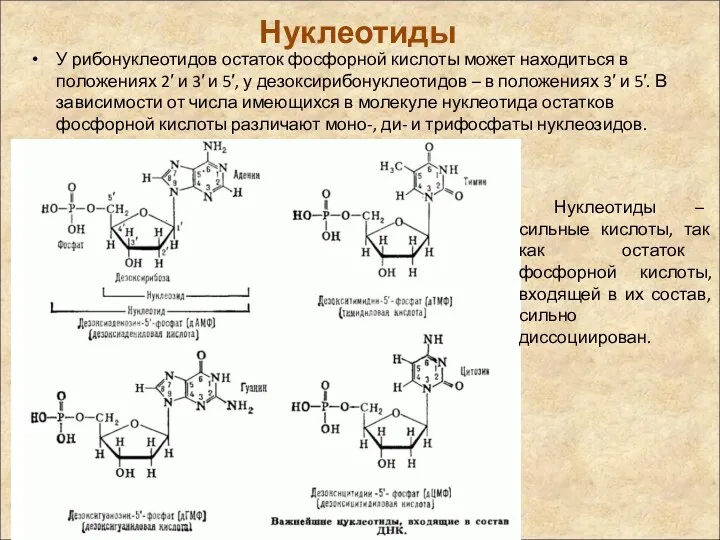
Нуклеотиды
У рибонуклеотидов остаток фосфорной кислоты может находиться в положениях 2′ и
Нуклеотиды
У рибонуклеотидов остаток фосфорной кислоты может находиться в положениях 2′ и
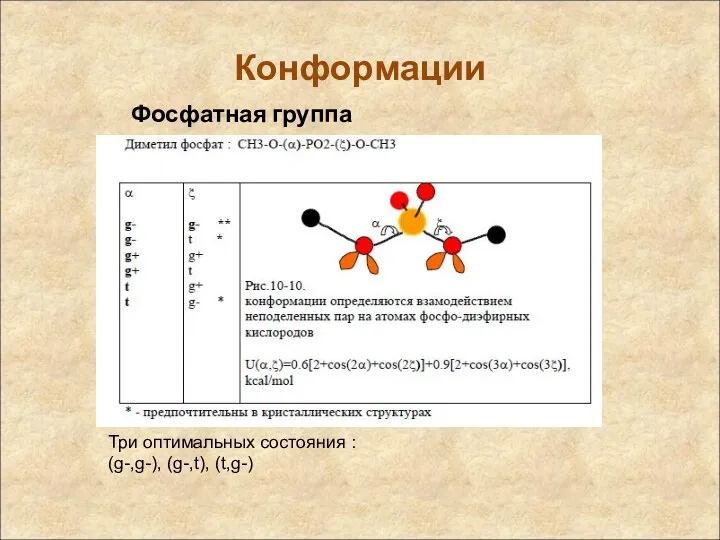
Конформации
Фосфатная группа
Три оптимальных состояния :
(g-,g-), (g-,t), (t,g-)
Конформации
Фосфатная группа
Три оптимальных состояния :
(g-,g-), (g-,t), (t,g-)

Биохимические функции нуклеотидов
1) являются строительными блоками нуклеиновых кислот (ДНК и РНК);
Биохимические функции нуклеотидов
1) являются строительными блоками нуклеиновых кислот (ДНК и РНК);
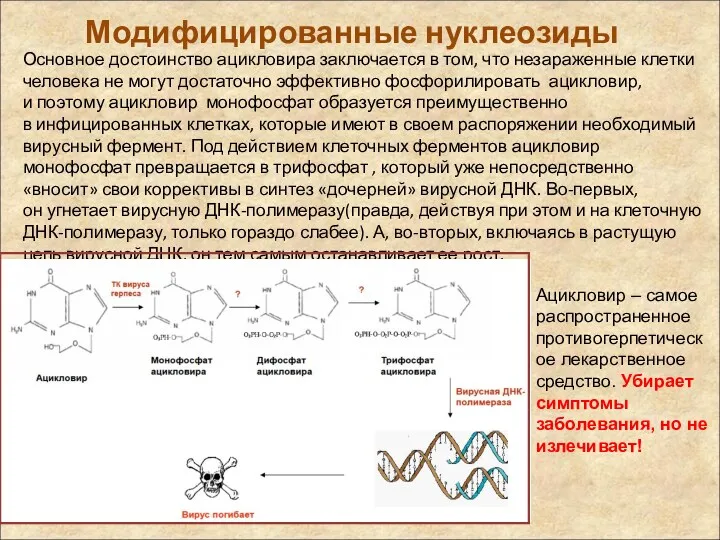
Модифицированные нуклеозиды
Основное достоинство ацикловира заключается в том, что незараженные клетки человека не могут
Модифицированные нуклеозиды
Основное достоинство ацикловира заключается в том, что незараженные клетки человека не могут
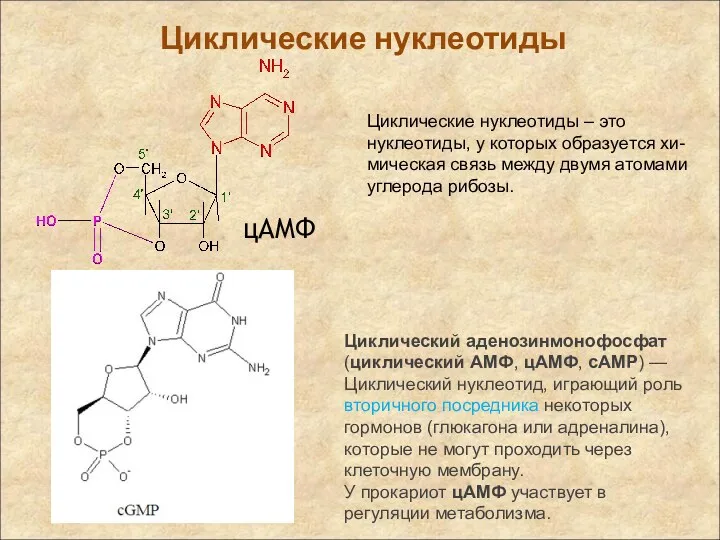
цAMФ
Циклические нуклеотиды
Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль
цAMФ
Циклические нуклеотиды
Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль

Нуклеиновые кислоты
Нуклеиновые кислоты хорошо растворимы в воде, практически не растворимы в
Нуклеиновые кислоты
Нуклеиновые кислоты хорошо растворимы в воде, практически не растворимы в
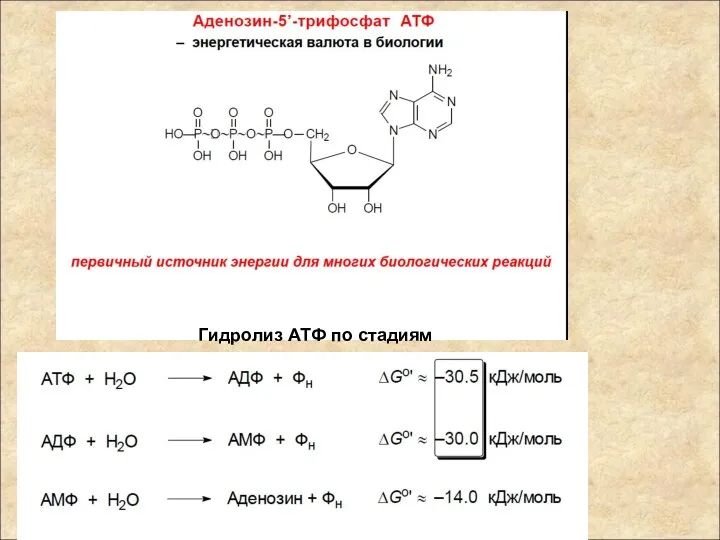
Гидролиз АТФ по стадиям
Гидролиз АТФ по стадиям

Нуклеиновые кислоты
ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания: пуриновые —
Нуклеиновые кислоты
ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания: пуриновые —

Первичная структура ДНК и РНК
Первичная структура ДНК и РНК
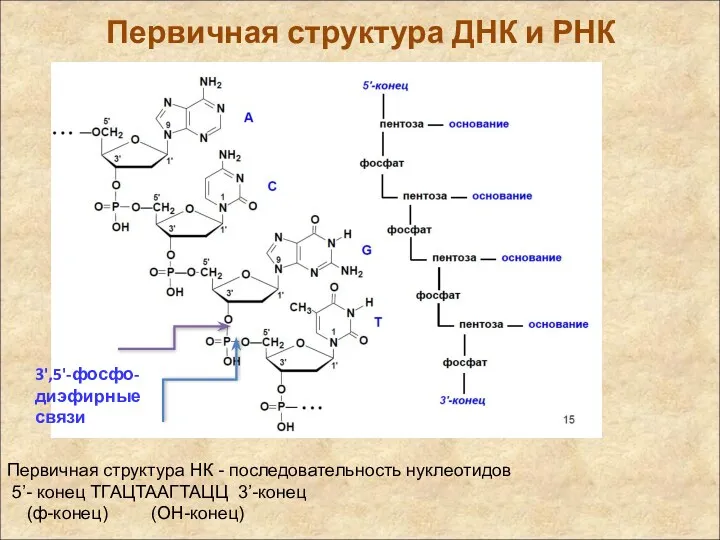
Первичная структура ДНК и РНК
Первичная структура НК - последовательность нуклеотидов
5’-
Первичная структура ДНК и РНК
Первичная структура НК - последовательность нуклеотидов
5’-
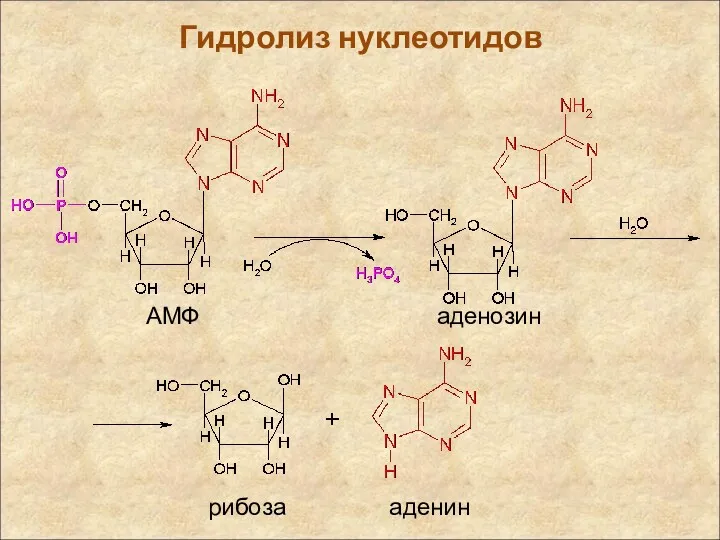
Гидролиз нуклеотидов
АМФ
аденозин
рибоза
аденин
Гидролиз нуклеотидов
АМФ
аденозин
рибоза
аденин

Вторичная структура ДНК
Вторичная структура ДНК

Вторичная структура ДНК
Вторичная структура ДНК
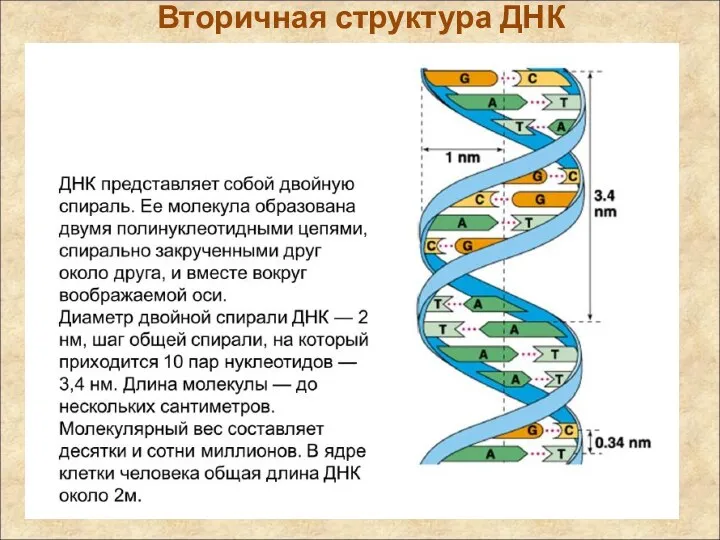
Вторичная структура ДНК
Вторичная структура ДНК
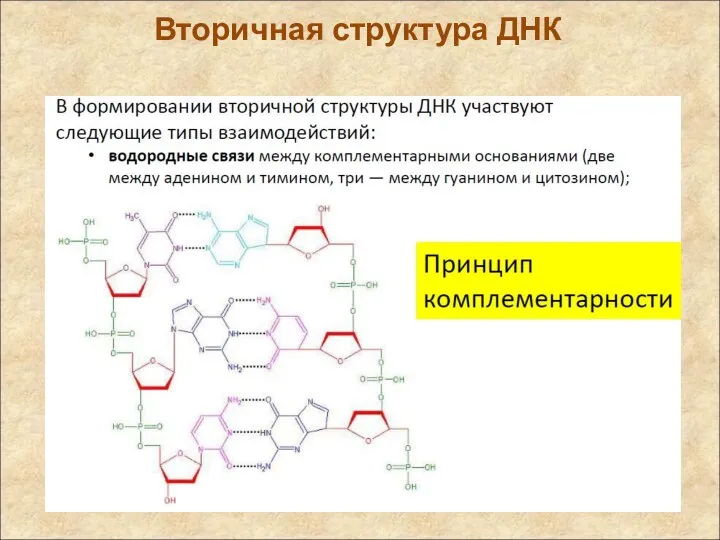
Вторичная структура ДНК
Вторичная структура ДНК

Вторичная структура
Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953)
Две
Вторичная структура
Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953)
Две
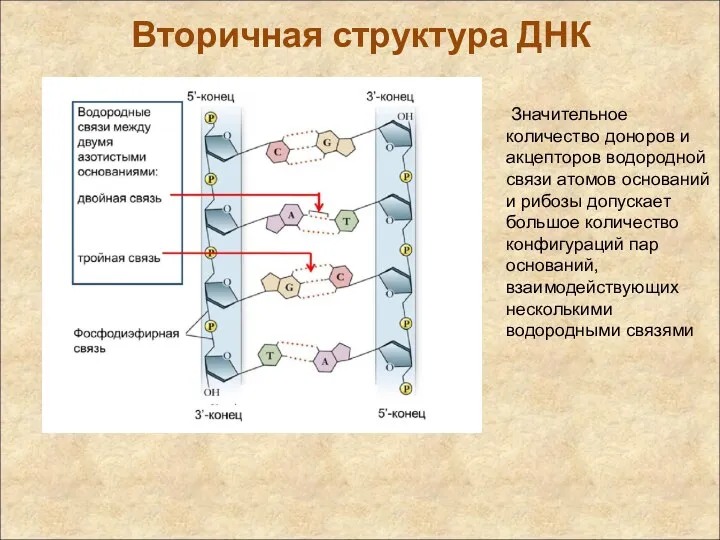
Вторичная структура ДНК
Значительное количество доноров и акцепторов водородной связи атомов
Вторичная структура ДНК
Значительное количество доноров и акцепторов водородной связи атомов

Образование связей между основаниями
Каждое основание на одной из цепей связывается
Образование связей между основаниями
Каждое основание на одной из цепей связывается

Взаимодействия копланарных оснований
Взаимодействия копланарных оснований
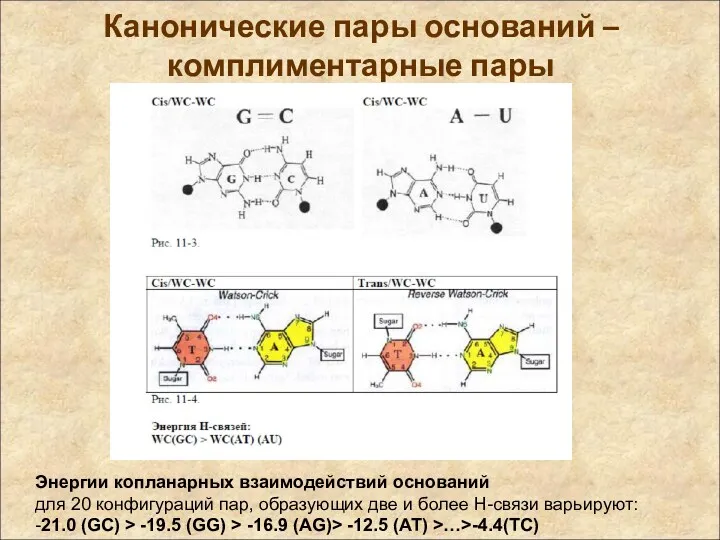
Канонические пары оснований – комплиментарные пары
Энергии копланарных взаимодействий оснований
для 20 конфигураций
Канонические пары оснований – комплиментарные пары
Энергии копланарных взаимодействий оснований
для 20 конфигураций
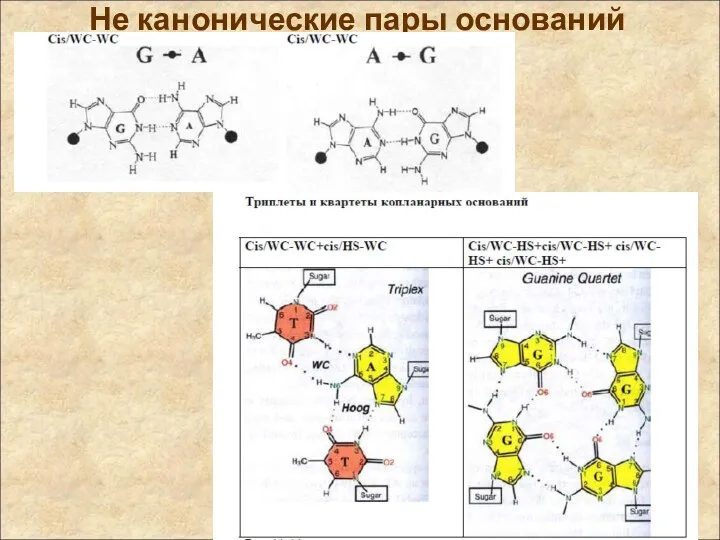
Не канонические пары оснований
Не канонические пары оснований
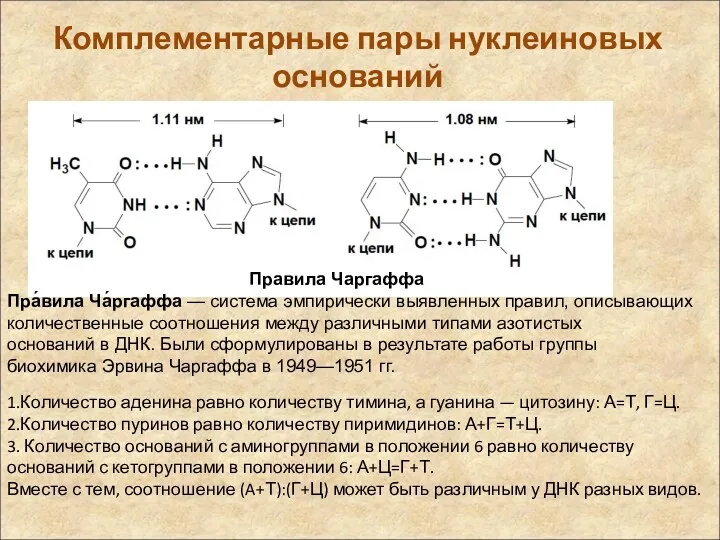
Комплементарные пары нуклеиновых оснований
Правила Чаргаффа
Пра́вила Ча́ргаффа — система эмпирически выявленных
Комплементарные пары нуклеиновых оснований
Правила Чаргаффа
Пра́вила Ча́ргаффа — система эмпирически выявленных
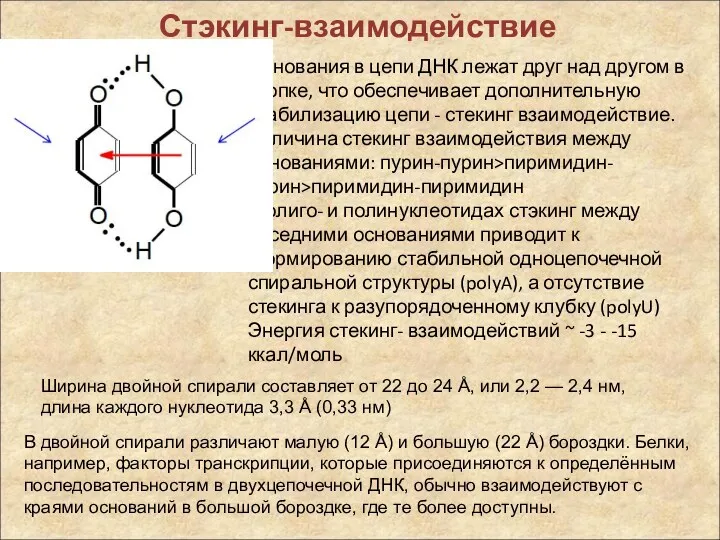
Стэкинг-взаимодействие
Основания в цепи ДНК лежат друг над другом в стопке, что
Стэкинг-взаимодействие
Основания в цепи ДНК лежат друг над другом в стопке, что

Стэкинг-взаимодействие
При этом оказывается, что стэкинг-взаимодействие между основаниями довольно специфично: полярные
Стэкинг-взаимодействие
При этом оказывается, что стэкинг-взаимодействие между основаниями довольно специфично: полярные
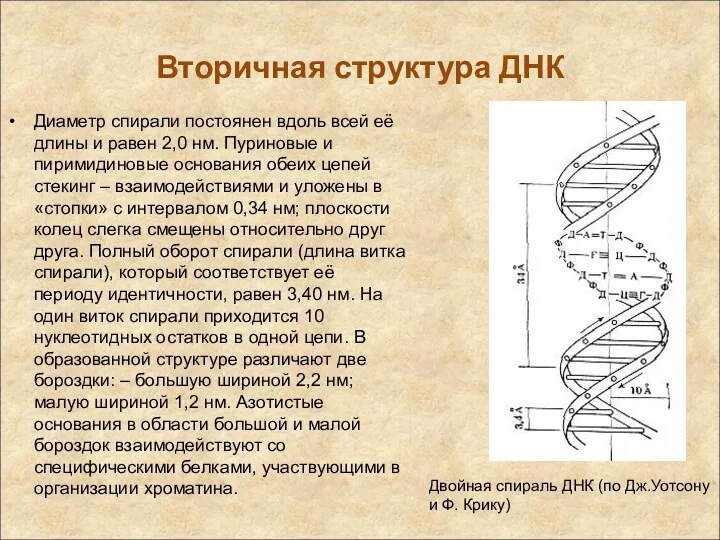
Вторичная структура ДНК
Диаметр спирали постоянен вдоль всей её длины и равен
Вторичная структура ДНК
Диаметр спирали постоянен вдоль всей её длины и равен
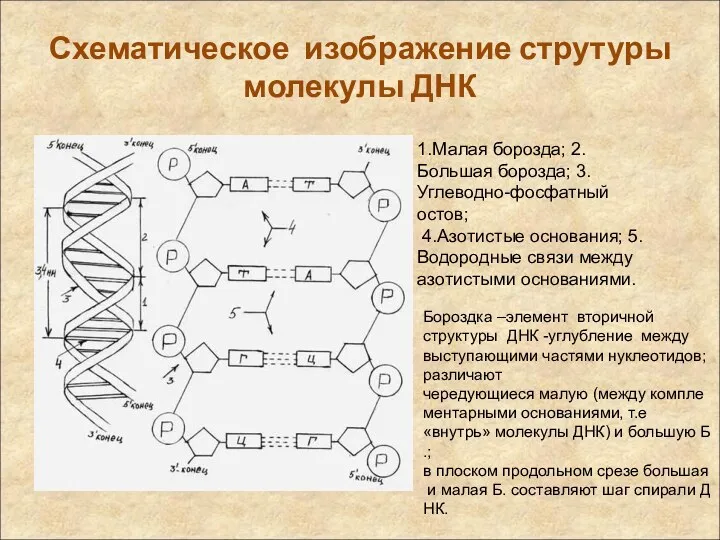
Схематическое изображение струтуры молекулы ДНК
1.Малая борозда; 2.Большая борозда; 3.Углеводно-фосфатный остов;
4.Азотистые
Схематическое изображение струтуры молекулы ДНК
1.Малая борозда; 2.Большая борозда; 3.Углеводно-фосфатный остов;
4.Азотистые
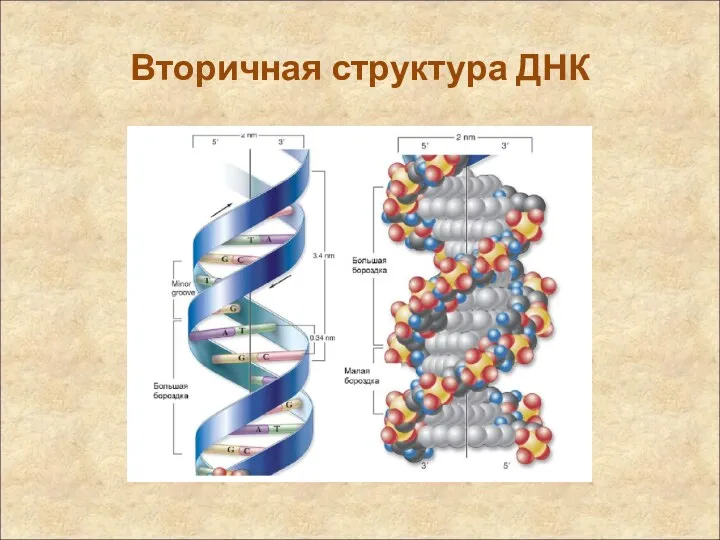
Вторичная структура ДНК
Вторичная структура ДНК
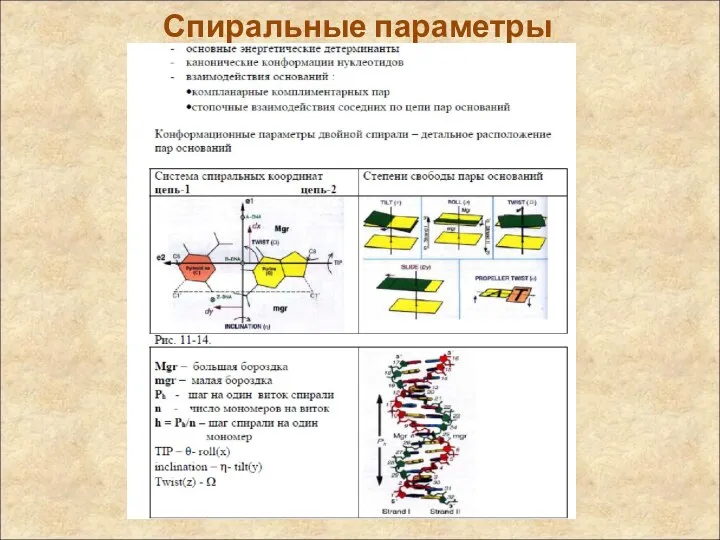
Спиральные параметры
Спиральные параметры
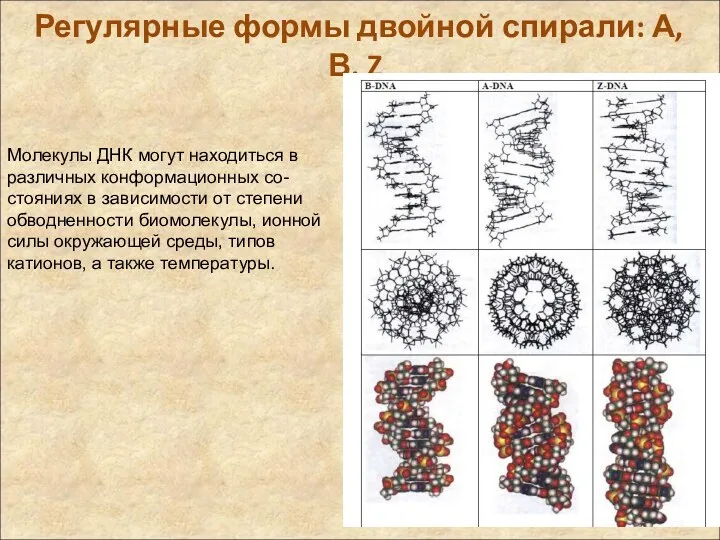
Регулярные формы двойной спирали: А, В, Z
Молекулы ДНК могут находиться в
Регулярные формы двойной спирали: А, В, Z
Молекулы ДНК могут находиться в
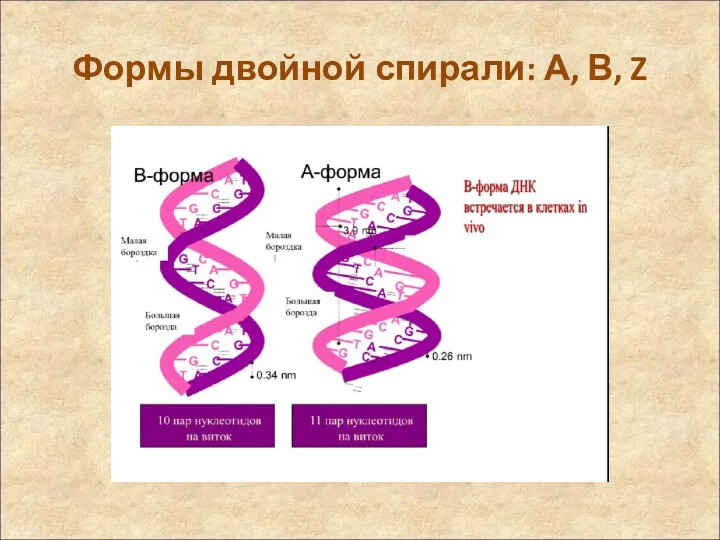
Формы двойной спирали: А, В, Z
Формы двойной спирали: А, В, Z

А-форма двойной спирали
Правозакрученные спирали образуют два семейства: А-семейство и В-семейство. А-семейство
А-форма двойной спирали
Правозакрученные спирали образуют два семейства: А-семейство и В-семейство. А-семейство

В-форма двойной спирали
Для В-форм ДНК характерно структурное разнообразие. Формы ДНК со
В-форма двойной спирали
Для В-форм ДНК характерно структурное разнообразие. Формы ДНК со

Формы двойной спирали: А, В, Z
Формы двойной спирали: А, В, Z

Z-форма ДНК
Z-форма ДНК имеет левозакрученную спираль и обнаружена у полинуклеотида с
Z-форма ДНК
Z-форма ДНК имеет левозакрученную спираль и обнаружена у полинуклеотида с

Z-форма ДНК
Область перехода перемещается вдоль спирали в виде небольшой петли. Переход
Z-форма ДНК
Область перехода перемещается вдоль спирали в виде небольшой петли. Переход


Регулярные формы двойной спирали: А, В, Z
Значения спиральных параметров и структурные
Регулярные формы двойной спирали: А, В, Z
Значения спиральных параметров и структурные
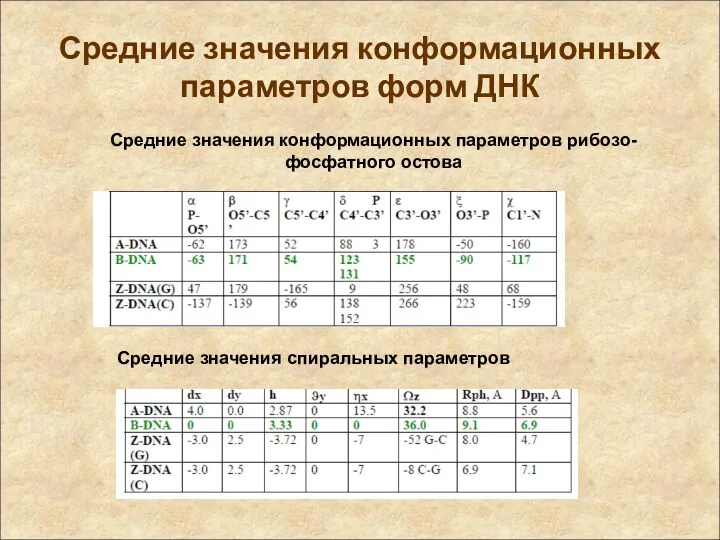
Средние значения конформационных параметров форм ДНК
Средние значения конформационных параметров рибозо-фосфатного остова
Средние
Средние значения конформационных параметров форм ДНК
Средние значения конформационных параметров рибозо-фосфатного остова
Средние
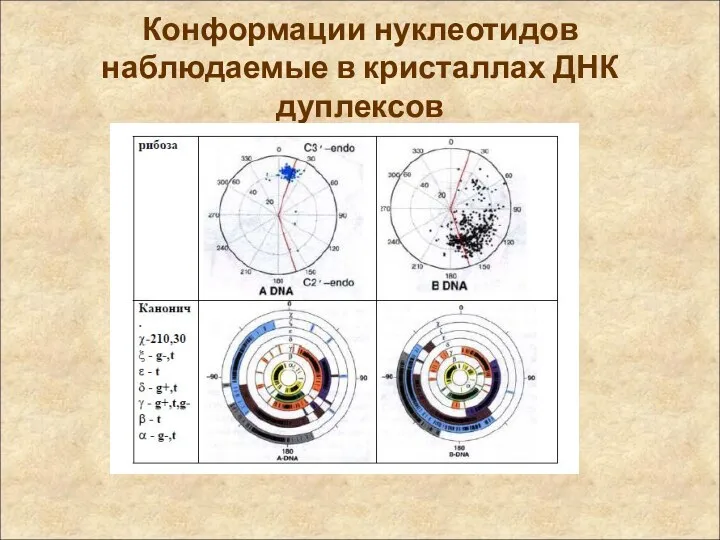
Конформации нуклеотидов
наблюдаемые в кристаллах ДНК дуплексов
Конформации нуклеотидов
наблюдаемые в кристаллах ДНК дуплексов
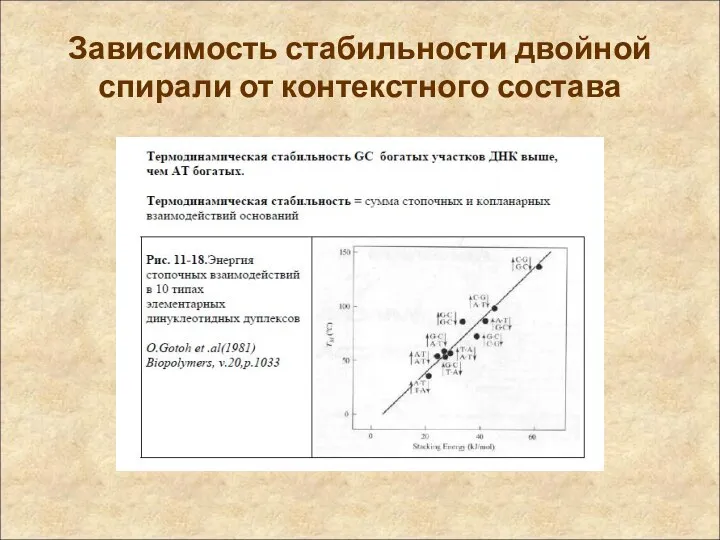
Зависимость стабильности двойной спирали от контекстного состава
Зависимость стабильности двойной спирали от контекстного состава

Сверхспирализация ДНК. (Суперскрученность)
Если взяться за концы верёвки и начать скручивать их
Сверхспирализация ДНК. (Суперскрученность)
Если взяться за концы верёвки и начать скручивать их

Физико-химические свойства ДНК
ДНК – довольно сильная многоосновная кислота, полностью ионизированная при
Физико-химические свойства ДНК
ДНК – довольно сильная многоосновная кислота, полностью ионизированная при

Физико-химические свойства ДНК
У большинства вирусов ДНК представляет собой двойную спираль, линейную
Физико-химические свойства ДНК
У большинства вирусов ДНК представляет собой двойную спираль, линейную

Спасибо за внимание!
Спасибо за внимание!






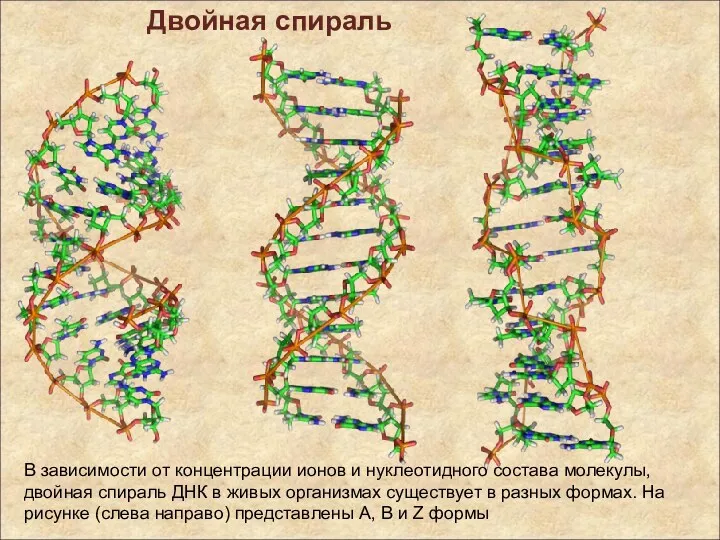
В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль
В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль
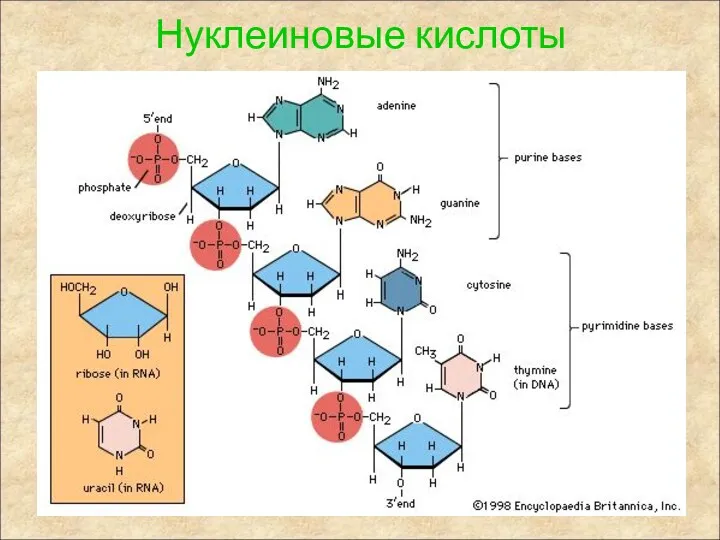
Нуклеиновые кислоты
Нуклеиновые кислоты
 Розв’язування задач за рівняннями реакцій з використанням розчинів із певною масовою часткою розчиненої речовини. Урок 13-14
Розв’язування задач за рівняннями реакцій з використанням розчинів із певною масовою часткою розчиненої речовини. Урок 13-14 Магия кристаллов
Магия кристаллов Пластмаси. Види пластмас. (11 клас)
Пластмаси. Види пластмас. (11 клас) Процессы и технологическая схема производства сегодня. АО Газпромнефть-ОНПЗ
Процессы и технологическая схема производства сегодня. АО Газпромнефть-ОНПЗ Производство полимеров
Производство полимеров Массасы 4,2 г көміртек (IV) оксиді сумен әрекеттескенде қанша грамм көмір қышқылы (Н2СО3) түзілетінін есепте
Массасы 4,2 г көміртек (IV) оксиді сумен әрекеттескенде қанша грамм көмір қышқылы (Н2СО3) түзілетінін есепте Геохимия гидротермального процесса
Геохимия гидротермального процесса Защита металлоконструкций от коррозии
Защита металлоконструкций от коррозии Химия металлов: влияние металлов на организм человека
Химия металлов: влияние металлов на организм человека Типы химических реакций
Типы химических реакций Органическая химия
Органическая химия Химия титанорганических соединений
Химия титанорганических соединений Соли и их свойства
Соли и их свойства Соединения серы
Соединения серы Химическая связь
Химическая связь Нанотехнологии в школьном образовании. Семинар учителей химии
Нанотехнологии в школьном образовании. Семинар учителей химии Генетическая связь между классами веществ
Генетическая связь между классами веществ Беломорская соль, проектная работа
Беломорская соль, проектная работа Химические свойства спиртов
Химические свойства спиртов Моделирование, как метод научного исследования. Ограниченный метод Хартри-Фока
Моделирование, как метод научного исследования. Ограниченный метод Хартри-Фока Арилалкиламины, гидроксифенилалкиламины и их производные
Арилалкиламины, гидроксифенилалкиламины и их производные Значение органической химии в жизни человека
Значение органической химии в жизни человека Классификация химических реакций
Классификация химических реакций Полезные геологические химические соединения. Получение меди из малахита
Полезные геологические химические соединения. Получение меди из малахита Механическая смесь и растворы
Механическая смесь и растворы Общая характеристика реакций органических соединений. Кислотные и основные свойства органических соединений
Общая характеристика реакций органических соединений. Кислотные и основные свойства органических соединений Применение солей
Применение солей Распределение электронов в атомах. 8 класс
Распределение электронов в атомах. 8 класс