- Хромопротеиды: биологическая роль. Синтез и распад гема. Метаболизм билирубина

Содержание
- 2. План лекции: Понятие о хромопротеинах, их классификация. Строение и функции гемопротеинов. Биосинтез гема. Распад гемопротеинов. Клинико-биохимические
- 3. 1. Понятие о хромопротеинах, их классификация. Белки бывают простые и сложные. У сложных белков имеется небелковая
- 4. Хромопротеины – это сложные белки, содержащие окрашенную простетическую группу. По этой причине молекулы хромопротеинов окрашены в
- 5. В природе известны следующие виды хромопротеинов: магнийпорфирины (хлорофилл) флавопротеины (рибофлавин) гемопротеины (гем)
- 6. Магнийпорфирины – это зелёные хлорофилл содержащие белки. Они образуются в растениях и сине-зелёных водорослях.
- 7. Структура хлорофилла
- 8. Их функция – это преобразование солнечной энергии в энергию макроэргических связей АТФ. Кроме того, именно хлорофилл
- 9. Флавопротеины Это жёлтые белки, содержащие рибофлавин. Все они являются ферментами класса оксидоредуктаз. Играют большую роль в
- 10. Структура ФМН
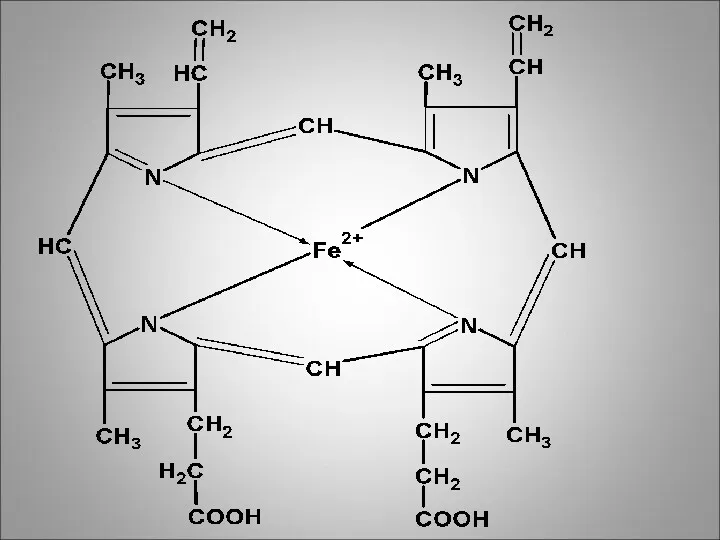
- 11. Гемопротеины В состав гемопротеинов входит гем, который придаёт им красное окрашивание. Гем – тетрапиррольный цикл, содержащий
- 12. Структура гема
- 13. 2. Строение и функции гемопротеинов. Гемопротеины подразделяются на: Дыхательные белки (гемоглобин, миоглобин) Ферменты
- 14. Гемоглобин – белок четвертичной структуры, тетрамер.
- 15. Два олигомера α-цепей состоят из 141 аминокислотных остатка, и два олигомера β-цепей – из 146 аминокислотных
- 16. Атом железа в гемоглобине может быть в состоянии Fe2+ или Fe3+, но только Fe2+ способен переносить
- 17. Гемоглобин способен транспортировать и углекислый газ – от тканей к лёгким (в виде карбогемоглобина). Но углекислый
- 18. Очень высокое сродство гемоглобина к окиси углерода (СО). При их взаимодействии образуется устойчивая молекула карбоксигемоглобина. Она
- 19. Миоглобин Миоглобин (молекулярная масса 16 000, состоит из 153 аминокислот) имеет третичную структуру, одну полипептидную цепь,
- 20. Миоглобин
- 21. Гем-содержащие ферменты Если в составе дыхательных гемопротеинов степень окисления железа не меняется, то в составе ферментов
- 22. Важнейшие гем-содержащие ферменты: каталаза цитохромы пероксидазы
- 23. 3. Биосинтез гема
- 24. Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени. Первая реакция
- 26. Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные этапы синтеза гема. Рассмотрим следующую
- 28. Затем происходит ряд последовательных реакций, в результате которых образуется тетрапиррольный цикл – протопорфирин. Фермент феррохелатаза, присоединяя
- 30. Источником железа для синтеза гема служит депонирующий железо белок ферритин. Синтезированный гем, соединяясь с α и
- 31. 4. Распад гемопротеинов
- 32. Рассмотрим распад хромопротеинов на примере гемоглобина, который количественно превосходит все другие белки этой группы. Продолжительность существования
- 33. В течение суток в организме животного разрушается 1–2 % эритроцитов крови. В одну секунду в организме
- 34. Распад гема начинается с разрыва одного метинового мостика между двумя пиррольными кольцами с сохранением на этой
- 36. Далее, от вердоглобина отщепляется ион железа и белок глобин. В результате образуется биливердин, который имеет линейную
- 37. Биливердин
- 38. Под влиянием биливердин-редуктазы биливердин при участии НАДФН2 восстанавливается в другой желчный пигмент – билирубин:
- 39. Билирубин
- 40. Образовавшийся в селезенке билирубин поступает в кровь. Билирубин плохо растворим в воде, поэтому он связывается с
- 41. Из сосудистого русла в гепатоциты билирубин попадает с помощью белков-переносчиков. Далее при участии белка лигандина он
- 42. После конъюгации билирубин секретируются в желчные протоки, в желчный пузырь и далее в кишечник. В начальных
- 43. В подвздошной и ободочной кишках коньюгаты билирубина атакуются бактериями и деконъюгируются. Бактериальные ферменты восстанавливают билирубин до
- 44. Уробилиноген в толстом кишечнике с помощью бактериальных ферментов может присоединять четыре атома водорода и переходить в
- 45. Большая часть метаболитов билирубина выводится из организма через кишечник. Во внешней среде они окисляются до стеркобилина
- 46. 5. Клинико-биохимические аспекты метаболизма гемопротеинов.
- 47. Количество гемоглобина в крови животных разных видов (г/л)
- 48. Основные причины снижения концентрации гемоглобина: Дефицит железа Дефицит витаминов (В6, В12, фолиевой кислоты) Кровопотери Алиментарное истощение
- 49. Основные причины повышения концентрации гемоглобина: Обезвоживание Гипоксия
- 50. Концентрация общего билирубина в крови животных разных видов (мкмоль/л)
- 51. Причины увеличения концентрации билирубина: Гемолиз эритроцитов (повышается концентрация непрямого билирубина) Цитолиз гепатоцитов (повышается в первую очередь
- 52. Если билирубин обнаруживается в моче, то это может быть только прямой билирубин (непрямой в мочу не
- 54. Скачать презентацию

План лекции:
Понятие о хромопротеинах, их классификация.
Строение и функции гемопротеинов.
Биосинтез гема.
Распад гемопротеинов.
Клинико-биохимические
План лекции:
Понятие о хромопротеинах, их классификация.
Строение и функции гемопротеинов.
Биосинтез гема.
Распад гемопротеинов.
Клинико-биохимические

1. Понятие о хромопротеинах, их классификация.
Белки бывают простые и сложные. У
1. Понятие о хромопротеинах, их классификация.
Белки бывают простые и сложные. У

Хромопротеины – это сложные белки, содержащие окрашенную простетическую группу. По этой
Хромопротеины – это сложные белки, содержащие окрашенную простетическую группу. По этой

В природе известны следующие виды хромопротеинов:
магнийпорфирины (хлорофилл)
флавопротеины (рибофлавин)
гемопротеины (гем)
В природе известны следующие виды хромопротеинов:
магнийпорфирины (хлорофилл)
флавопротеины (рибофлавин)
гемопротеины (гем)

Магнийпорфирины – это зелёные хлорофилл содержащие белки. Они образуются в растениях
Магнийпорфирины – это зелёные хлорофилл содержащие белки. Они образуются в растениях
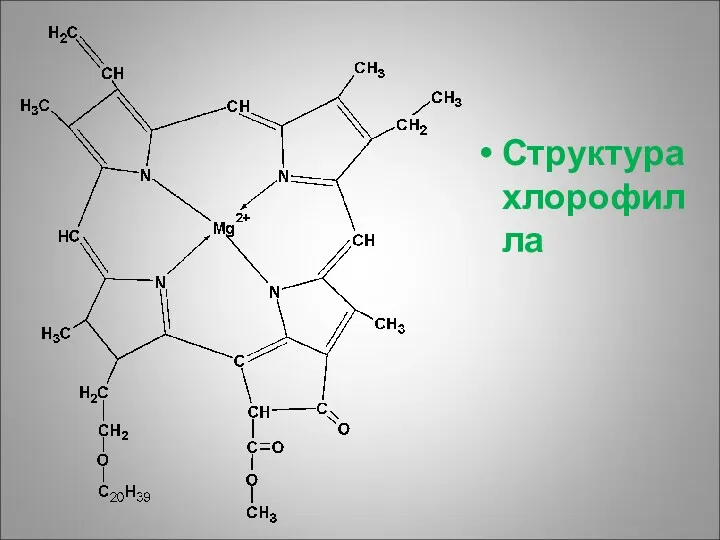
Структура хлорофилла

Их функция – это преобразование солнечной энергии в энергию макроэргических связей
Их функция – это преобразование солнечной энергии в энергию макроэргических связей

Флавопротеины
Это жёлтые белки, содержащие рибофлавин. Все они являются ферментами класса оксидоредуктаз.
Флавопротеины
Это жёлтые белки, содержащие рибофлавин. Все они являются ферментами класса оксидоредуктаз.
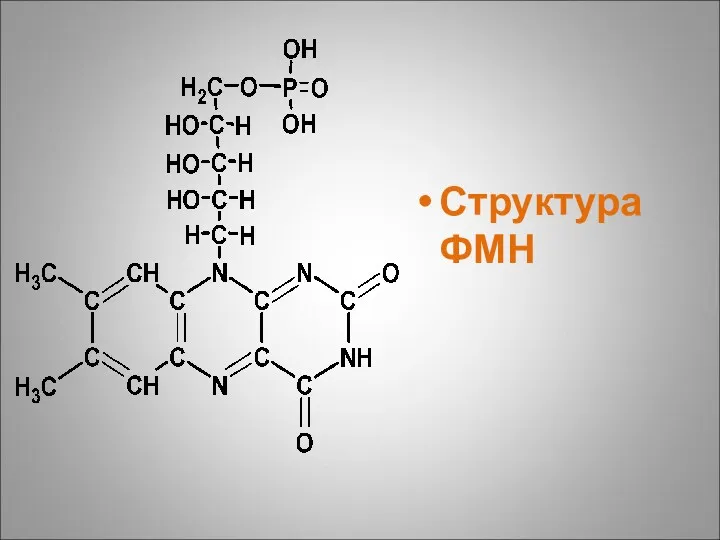
Структура ФМН
Структура ФМН

Гемопротеины
В состав гемопротеинов входит гем, который придаёт им красное окрашивание.
Гем –
Гемопротеины
В состав гемопротеинов входит гем, который придаёт им красное окрашивание.
Гем –
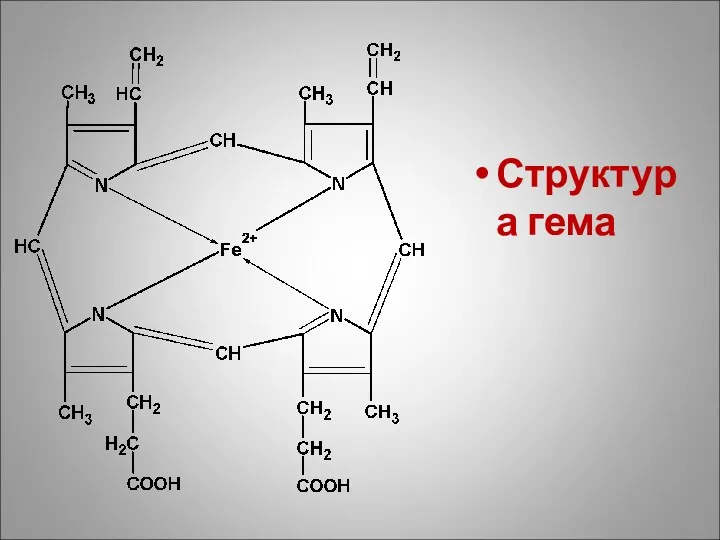
Структура гема
Структура гема

2. Строение и функции гемопротеинов.
Гемопротеины подразделяются на:
Дыхательные белки (гемоглобин, миоглобин)
Ферменты
2. Строение и функции гемопротеинов.
Гемопротеины подразделяются на:
Дыхательные белки (гемоглобин, миоглобин)
Ферменты
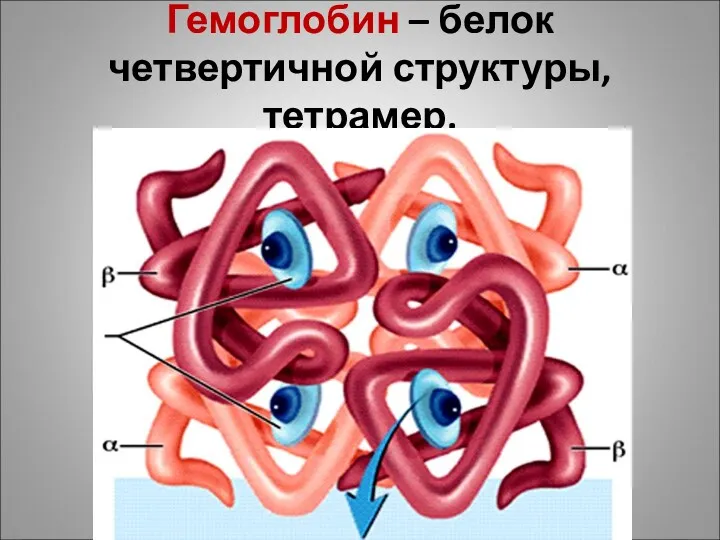
Гемоглобин – белок четвертичной структуры, тетрамер.
Гемоглобин – белок четвертичной структуры, тетрамер.

Два олигомера α-цепей состоят из 141 аминокислотных остатка, и два олигомера
Два олигомера α-цепей состоят из 141 аминокислотных остатка, и два олигомера

Атом железа в гемоглобине может быть в состоянии Fe2+ или Fe3+,
Атом железа в гемоглобине может быть в состоянии Fe2+ или Fe3+,

Гемоглобин способен транспортировать и углекислый газ – от тканей к лёгким
Гемоглобин способен транспортировать и углекислый газ – от тканей к лёгким

Очень высокое сродство гемоглобина к окиси углерода (СО). При их взаимодействии
Очень высокое сродство гемоглобина к окиси углерода (СО). При их взаимодействии

Миоглобин
Миоглобин (молекулярная масса 16 000, состоит из 153 аминокислот) имеет третичную структуру,
Миоглобин
Миоглобин (молекулярная масса 16 000, состоит из 153 аминокислот) имеет третичную структуру,
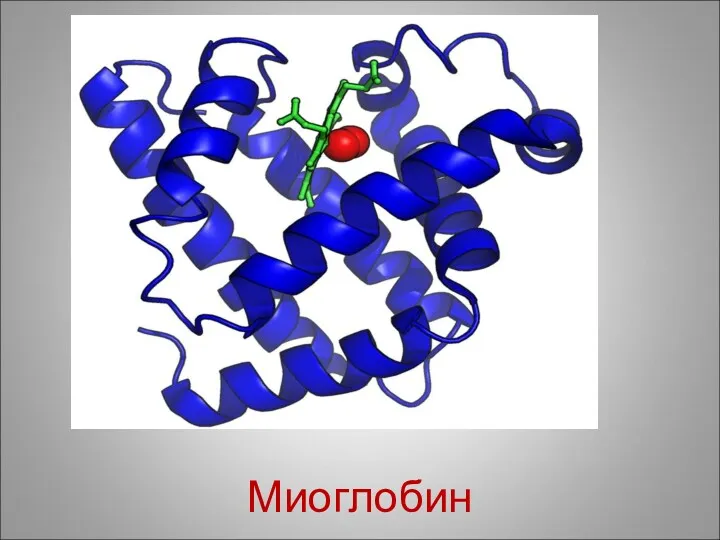
Миоглобин
Миоглобин

Гем-содержащие ферменты
Если в составе дыхательных гемопротеинов степень окисления железа не меняется,
Гем-содержащие ферменты
Если в составе дыхательных гемопротеинов степень окисления железа не меняется,

Важнейшие гем-содержащие ферменты:
каталаза
цитохромы
пероксидазы
Важнейшие гем-содержащие ферменты:
каталаза
цитохромы
пероксидазы

3. Биосинтез гема
3. Биосинтез гема

Гем синтезируется во всех тканях, но с наибольшей скоростью в костном
Гем синтезируется во всех тканях, но с наибольшей скоростью в костном
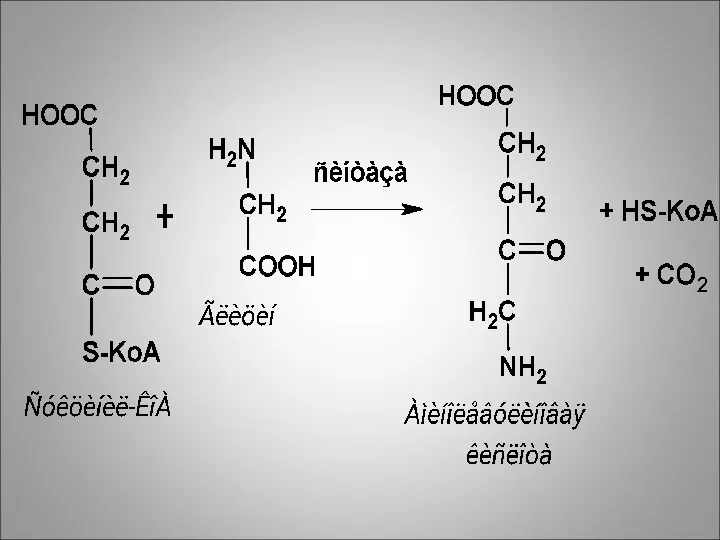

Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные
Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные
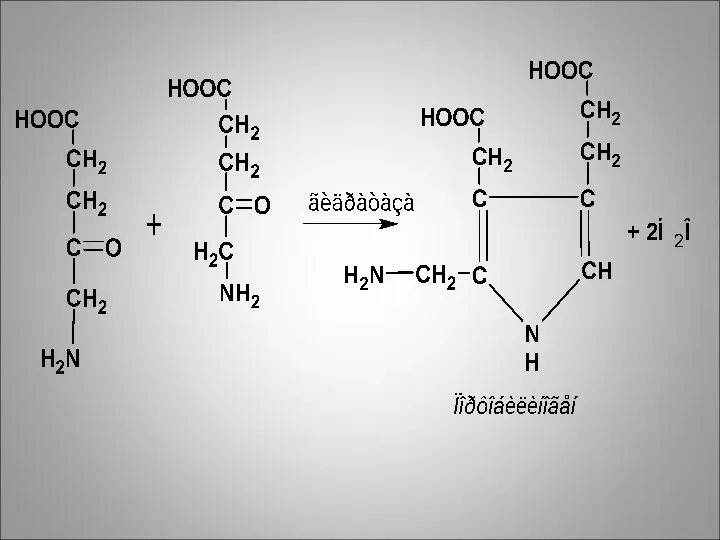

Затем происходит ряд последовательных реакций, в результате которых образуется тетрапиррольный цикл
Затем происходит ряд последовательных реакций, в результате которых образуется тетрапиррольный цикл

Источником железа для синтеза гема служит депонирующий железо белок ферритин. Синтезированный
Источником железа для синтеза гема служит депонирующий железо белок ферритин. Синтезированный

4. Распад гемопротеинов
4. Распад гемопротеинов

Рассмотрим распад хромопротеинов на примере гемоглобина, который количественно превосходит все другие
Рассмотрим распад хромопротеинов на примере гемоглобина, который количественно превосходит все другие

В течение суток в организме животного разрушается 1–2 % эритроцитов крови.
В течение суток в организме животного разрушается 1–2 % эритроцитов крови.

Распад гема начинается с разрыва одного метинового мостика между двумя пиррольными
Распад гема начинается с разрыва одного метинового мостика между двумя пиррольными
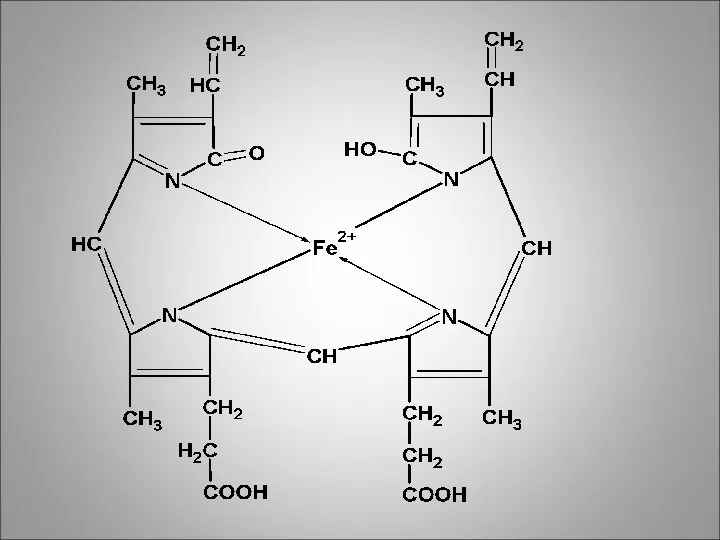

Далее, от вердоглобина отщепляется ион железа и белок глобин. В результате
Далее, от вердоглобина отщепляется ион железа и белок глобин. В результате
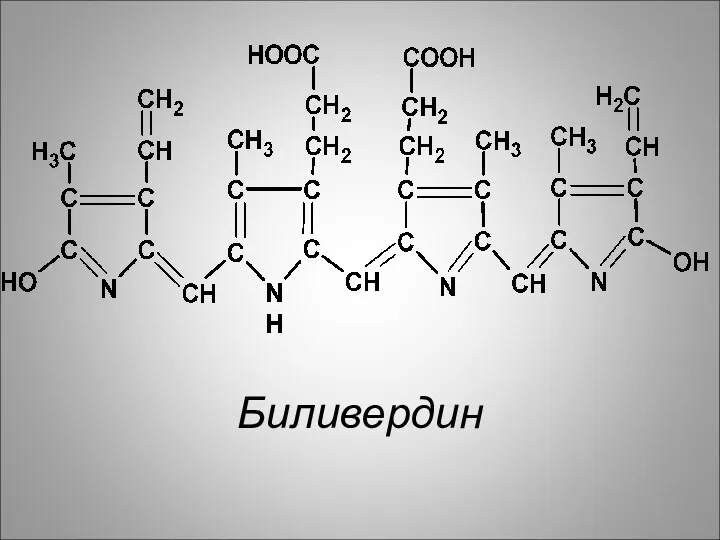
Биливердин
Биливердин

Под влиянием биливердин-редуктазы биливердин при участии НАДФН2 восстанавливается в другой
Под влиянием биливердин-редуктазы биливердин при участии НАДФН2 восстанавливается в другой
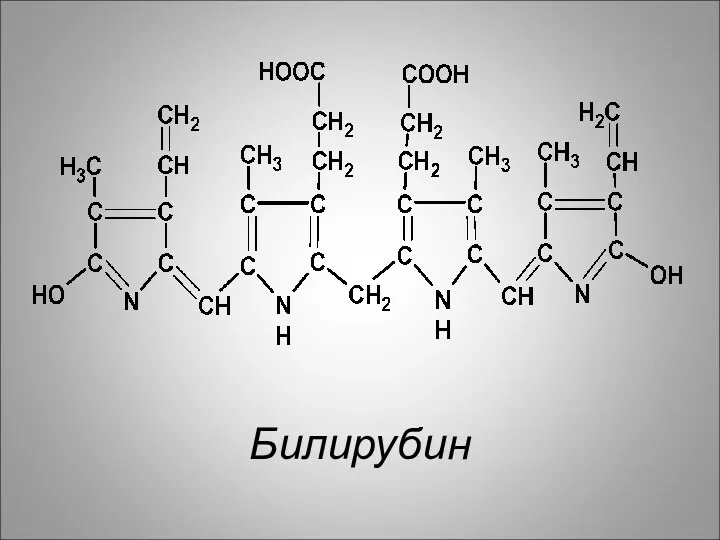
Билирубин
Билирубин

Образовавшийся в селезенке билирубин поступает в кровь. Билирубин плохо растворим в
Образовавшийся в селезенке билирубин поступает в кровь. Билирубин плохо растворим в

Из сосудистого русла в гепатоциты билирубин попадает с помощью белков-переносчиков.
Далее при
Из сосудистого русла в гепатоциты билирубин попадает с помощью белков-переносчиков.
Далее при

После конъюгации билирубин секретируются в желчные протоки, в желчный пузырь и
После конъюгации билирубин секретируются в желчные протоки, в желчный пузырь и

В подвздошной и ободочной кишках коньюгаты билирубина атакуются бактериями и деконъюгируются.
В подвздошной и ободочной кишках коньюгаты билирубина атакуются бактериями и деконъюгируются.

Уробилиноген в толстом кишечнике с помощью бактериальных ферментов может присоединять четыре
Уробилиноген в толстом кишечнике с помощью бактериальных ферментов может присоединять четыре

Большая часть метаболитов билирубина выводится из организма через кишечник. Во внешней
Большая часть метаболитов билирубина выводится из организма через кишечник. Во внешней

5. Клинико-биохимические аспекты метаболизма гемопротеинов.
5. Клинико-биохимические аспекты метаболизма гемопротеинов.

Количество гемоглобина в крови животных разных видов (г/л)
Количество гемоглобина в крови животных разных видов (г/л)

Основные причины снижения концентрации гемоглобина:
Дефицит железа
Дефицит витаминов (В6, В12, фолиевой кислоты)
Кровопотери
Алиментарное
Основные причины снижения концентрации гемоглобина:
Дефицит железа
Дефицит витаминов (В6, В12, фолиевой кислоты)
Кровопотери
Алиментарное

Основные причины повышения концентрации гемоглобина:
Обезвоживание
Гипоксия
Основные причины повышения концентрации гемоглобина:
Обезвоживание
Гипоксия

Концентрация общего билирубина в крови животных разных видов (мкмоль/л)
Концентрация общего билирубина в крови животных разных видов (мкмоль/л)

Причины увеличения концентрации билирубина:
Гемолиз эритроцитов (повышается концентрация непрямого билирубина)
Цитолиз гепатоцитов (повышается
Причины увеличения концентрации билирубина:
Гемолиз эритроцитов (повышается концентрация непрямого билирубина)
Цитолиз гепатоцитов (повышается

Если билирубин обнаруживается в моче, то это может быть только прямой
Если билирубин обнаруживается в моче, то это может быть только прямой
 Удивительные свойства воды
Удивительные свойства воды Electronică aplicată. (Curs 2)
Electronică aplicată. (Curs 2) Пространственное строение органических соединений. (Лекция 2)
Пространственное строение органических соединений. (Лекция 2) 20230416_urok_na_20.02.2023
20230416_urok_na_20.02.2023 Неізогіпсичні перетворення функціональних груп. Реакції окиснення
Неізогіпсичні перетворення функціональних груп. Реакції окиснення Виды присадок к моторным топливам. Присадки к дизельному топливу
Виды присадок к моторным топливам. Присадки к дизельному топливу Электронная конфигурация К. Классификация химических элементов. 8 класс
Электронная конфигурация К. Классификация химических элементов. 8 класс Спирты. Физические и химические свойства
Спирты. Физические и химические свойства Сучасні методи органічного синтезу. Ізогіпсичні перетворення функціональних груп. Реакції приєднання. Гідроборування
Сучасні методи органічного синтезу. Ізогіпсичні перетворення функціональних груп. Реакції приєднання. Гідроборування 20230321_obobshchenie_po_elektroliticheskoy_dissotsiatsii
20230321_obobshchenie_po_elektroliticheskoy_dissotsiatsii Классификация органических соединений, углеводородов
Классификация органических соединений, углеводородов Типовые процессы технологии неорганических веществ. (Тема 3)
Типовые процессы технологии неорганических веществ. (Тема 3) ЕГЭ Химия. Задание №5
ЕГЭ Химия. Задание №5 Изучение строения и свойств глюкозы
Изучение строения и свойств глюкозы Железо и его соединения
Железо и его соединения Химические превращения веществ
Химические превращения веществ Белки. Функции белков
Белки. Функции белков Високомолекулярні сполуки. Полімери
Високомолекулярні сполуки. Полімери Простые спектры 1Н ЯМР
Простые спектры 1Н ЯМР Значення хімічних процесів у природі
Значення хімічних процесів у природі Реакції йонного обміну між електролітами у водних розчинах. Практична робота №1. 9 клас
Реакції йонного обміну між електролітами у водних розчинах. Практична робота №1. 9 клас Реакции ионного обмена
Реакции ионного обмена Жиры. Мыла. 10 класс
Жиры. Мыла. 10 класс Группа редкоземельных элементов
Группа редкоземельных элементов Аммиак. № 12 зертханалық тәжірибе Аммиак молекуласының моделі
Аммиак. № 12 зертханалық тәжірибе Аммиак молекуласының моделі Железо
Железо Crystallography. Part 4: Crystal Forms Twinning
Crystallography. Part 4: Crystal Forms Twinning Функциональные производные с простой связью C-“Э. Галогенпроизводные. (Лекция 5)
Функциональные производные с простой связью C-“Э. Галогенпроизводные. (Лекция 5)