- Липиды и обмен липидов

Содержание
- 4. ФУНКЦИИ ЛИПИДОВ 1. Энергетическая (1 г липидов при окислении дает 9,3 ккал; 1 моль пальмитиновой кислоты
- 5. Функции липидов Структурная: холестерол и фосфолипиды – структурные компоненты мембран. Метаболическая: холестерол образует большое число биоактивных
- 6. Функции липидов Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат (вторичные мессенджеры гормонов); производные
- 7. Патология липидного обмена Первичные нарушения: Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз) Ожирение Желчнокаменная болезнь Метаболический ацидоз (кетонемия) Вторичные
- 8. Жирные кислоты > 70 жирных кислот идентифицированы в организме. Свободные жирные кислоты находятся в основном в
- 9. ЖИРНЫЕ КИСЛОТЫ Галогеновое число определяет число двойных связей в жирной кислоте. У человека возможен синтез только
- 10. Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ Семейства простагландинов, тромбоксанов, лейкотриенов др. - биологически активные вещества, действуют
- 11. ПРОСТАГЛАНДИНЫ Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4). Ингибируется аспирином по механизму ковалентной модификации фермента
- 12. Внешний обмен липидов Потребность в жирах – 80 -100 г/сут (возрастает при необходимости перехода энергетического обмена
- 13. ВНЕШНИЙ ОБМЕН ЛИПИДОВ Основные эмульгаторы жиров в ЖКТ – желчные кислоты – производные холестерола, но и
- 17. ВНЕШНИЙ ОБМЕН ЛИПИДОВ Гидролиз нейтральных жиров в 12 –п кишечнике: моно- , ди- и триглицеридлипазы: продукты
- 18. ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ Мелкие эмульгированные капли жира (до 0,5 мкм) проникают через кишечную стенку
- 19. Ресинтез жиров в энтероцитах На основе продуктов гидролиза экзо- генных жиров в энтероцитах синтезиру- ются видоспецифичные
- 20. Фосфолипиды образуются на основе фосфатидной кислоты и активных форм холина, серина, этаноламина или инозитола. O II
- 21. Транспортные формы экзогенных липидов * Глицерол и СЖК (С * СЖК (С>10) покидают энтероциты через кишечную
- 22. ХМ – транспортная форма экзогенных липидов ХМ образуются в энтероцитах: * Незрелые ХМ (насцентные) – 85%
- 23. Схема строения частицы ХМ
- 24. * В кровяном русле незрелые ХМ получа-ют от ЛПВП апо-Е, апо-С-II (кофактор ЛПЛ) и апо-А-IV. Этим
- 25. «Iceberg-sea» - модель строения частицы сывороточного липопротеида Assmann G. & Brever J. (1974)
- 26. Функции ХМ * ХМ доставляют экзогенные липиды в печень, жировую ткань, миокард и скелетные мышцы. ТАГ
- 27. ХМ, отдав часть ТАГ в результате их гидро-лиза ЛПЛ, превращаются в ремнантные ХМ (р-ХМ), которые поглощаются
- 28. Промежуточный обмен липидов Внутриклеточный липолиз Адипоциты или клетки жировой ткани (подкожный жир, малый и большой сальники
- 29. Механизм активации гормончувствительной ТАГ-липазы адипоцитов В постабсорбтивном периоде липолиз в ади-поцитах активируется глюкагоном; * При физической
- 30. * В результате активации аденилатцик-лазы повышается концентрация ц-АМФ, которая активирует протеинкиназу А (ПКА). * ПКА фосфорилирует
- 31. Суммарный результат гидролиза ТАГ в адипоцитах Гормончувствительная ТАГ-липаза сначала отщепляет ЖК в положении 1 = 1
- 32. Метаболизм глицерола Глицерол гидрофилен и переносится по крови в свободном виде. Ещё один путь образования глицерола
- 33. * Глицерол является субстратом для: - липогенеза; - глюконеогенеза - или может окисляется через диокси-ацетонфосфат по
- 34. * СЖК транспортируются по крови в комплексе с альбумином – молекула альбумина имеет 7 специфических сайтов
- 35. Окисление жирных кислот Путь окисления СЖК, сопряженный с синте-зом АТФ, протекает в митохондриях [Ю.Кеннеди и А.Ленинджер,
- 36. Опыты Франца Кнопа К ω-атому углерода ЖК с неразветвленной цепью присоединена фенильная группа. Продукты свидетельствуют о
- 37. Активация и транспорт ЖК в митохондрии 1. Активация ЖК идет с участием ацил-КоА-синтетазы (тиокиназы ЖК), локализованной
- 38. 2. Проникновение активированной ЖК в матрикс митохондрий: 2.1. Короткоцепочечные ЖК (С 2.2. Длинноцепочечные ЖК (С>10) проникают
- 39. 2.3. Обратное превращение: ацилкарнитин ? ацил-КоА Происходит с участием фермента, локализованном на внутренней поверхности внутренней мембраны
- 40. Реакции β-окисления жирных кислот Путь β-окисления – повторяющаяся последова- тельность четырех реакций. На каждом этапе окисления
- 41. Ацил-КоА-дегидрогеназа. Дегидрирование по α- и β- С атомам (положения 2 и 3). Атомы водорода переносятся на
- 42. 3-гидроксиацил-СоА-дегидрогеназа. Специ- фический акцептор электронов – NAD+ Образовавшийся NADH передает восстано- вительные эквиваленты на NADH-дегидроге- назу
- 43. Особенности β-окисления ЖК с нечетным числом С-атомов и ненасыщенных ЖК 1. ЖК с нечетным числом С-атомов
- 44. 2. Ненасыщенные ЖК (содержат двойные связи): Требует участия дополнительных ферментов: 1. Если ЖК имеет 1 двойную
- 45. Из цис-Δ3-еноил-КоА получается транс-Δ2-еноил-КоА, который яв- ляется нормальным субстратом для еноил-КоА-гидратазы.
- 46. Эпимераза превращает D-стереоизомер в L-стереоизомер, который является субстратом для 3-гидроксиацил-СоА-дегидрогеназы.
- 47. 2. Если окисляется ЖК с двумя двойными связями – линолевая к-та (С18:2, цис- Δ9,Δ12) I фермент:
- 48. Энергетика окисления жирных кислот Каждый этап β –окисления сопровождается образованием ФАДН2 и НАДН (реокисление их в
- 49. Ацетил-КоА, как продукт окисления ЖК, далее окисляется в ЦТК («Жиры сгорают в пламени углеводов»). В норме,
- 50. Выход АТФ при β-окислении пальмитиновой кислоты Каждый этап β–окисления сопровождается образованием ФАДН2 и НАДН. Их реокисление
- 51. КЕТОГЕНЕЗ Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит к активации кетогенеза в
- 52. При голодании и диабете (окисление ЖК усиливается, а глюкозы – подавляется): Ацетил-КоА образуется в избытке; Концентрация
- 53. 3-кетотиолаза Гидроксиметил- глутарил-КоА- синтетаза Гидроксиметил- глутарил-КоА- лиаза D-3-гидрокси- бутират-ДГ Спонтанное декарбокси- лирование (необратимо) («главное» КТ) Покидает
- 54. Длительность голодания, сутки Концентрация в сыворотке крови, mmol / l
- 55. Ацетоацетат и гидроксибутират свободно диффундируют (по градиенту концентрации) из гепатоцитов в кровь и доставляются к перифе-
- 56. Окисление кетоновых тел Печень не способна утилизировать кетоновые тела. В периферических тканях гидроксибутират окисляется до ацетоацетата.
- 57. Печень не может потреблять кетоновые тела, которые она синтезирует Во многих тканях (кроме печени), ацетоацетат может
- 58. Кетоацидоз При длительном голодании и при диабете концентрация кетоновых тел в крови может достигать чрезвычайно высоких
- 59. Липогенез Абсорбтивный период. Инсулин. Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира в энтероцитах (этерификация
- 60. ЛИПОГЕНЕЗ Субстраты: глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах; диоксиацетонфосфат ДГ (фосфоглицерол ДГ) в миоцитах и
- 61. Липогенез Избыток глюкозы обеспечивает: 1. запас гликогена (ограничен) 2. ДАФ ?глицерол-3-фосфат 3. ПВК ? ацетил-КоА ?
- 62. Биосинтез жирных кислот Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления! Другая ферментативная система и локализация
- 64. Транспорт ацетил-КоА в цитоплазму В ситуации накопления АТФ и НАДН ингибируется изоцитрат ДГ и накапливающийся цитрат
- 65. Транспорт ацетил-КоА в цитоплазму Возвращение ОА в МХ: 1.ОА –> малат (НАД зависимая цитоплазматическая МДГ); Малат
- 67. Образование малонил-КоА ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот, не входит в состав мультиферментного
- 68. Синтетаза жирных кислот мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно). 6 ферментов и АПБ (2
- 70. АПБ – структура и функция Активный центр этого белка – фосфопантотеновая кислота и тиоэтиламин (аналогичные структуре
- 73. Пальмитоил- синтетаза Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7 СО2 +SH-АПБ + 14
- 74. Синтез триацилглицеролов Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию образования ациладенилатов. Перенос ацилтрансферазой
- 77. Синтез фосфолипидов Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного синтеза (фосфатидная кислота). Синтез
- 82. Синтез сфинголипидов
- 84. Синтез холестерола 1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА 2. восстановление НАДФН-редуктазой? мевалоновая кислота 3. фосфорилирование и декарбоксилирование?
- 85. Метаболизм холестерола 0,5 г эндогенного и 0,5 г экзогенного ХЛ 10% ХЛ находится в виде эфиров
- 91. Транспортные формы ХЛ Экзогенный ХЛ транспортируется в составе хиломикронов, ремнантные формы которых поглощаются печенью. Эндогенный ХЛ,
- 93. Скачать презентацию

ФУНКЦИИ ЛИПИДОВ
1. Энергетическая (1 г липидов при окислении дает 9,3 ккал;
ФУНКЦИИ ЛИПИДОВ
1. Энергетическая (1 г липидов при окислении дает 9,3 ккал;

Функции липидов
Структурная: холестерол и фосфолипиды – структурные компоненты мембран.
Метаболическая: холестерол
Функции липидов
Структурная: холестерол и фосфолипиды – структурные компоненты мембран.
Метаболическая: холестерол

Функции липидов
Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат
Функции липидов
Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат

Патология липидного обмена
Первичные нарушения:
Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз)
Ожирение
Желчнокаменная болезнь
Метаболический ацидоз (кетонемия)
Вторичные гиперлипопротеинемии
Патология липидного обмена
Первичные нарушения:
Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз)
Ожирение
Желчнокаменная болезнь
Метаболический ацидоз (кетонемия)
Вторичные гиперлипопротеинемии

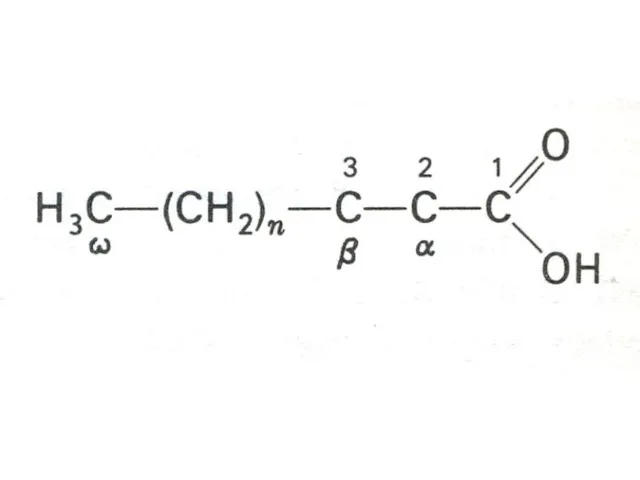
Жирные кислоты
> 70 жирных кислот идентифицированы в организме.
Свободные жирные кислоты находятся
Жирные кислоты
> 70 жирных кислот идентифицированы в организме.
Свободные жирные кислоты находятся

ЖИРНЫЕ КИСЛОТЫ
Галогеновое число определяет число двойных связей в жирной кислоте.
У
ЖИРНЫЕ КИСЛОТЫ
Галогеновое число определяет число двойных связей в жирной кислоте.
У

Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ
Семейства простагландинов, тромбоксанов, лейкотриенов др. -
Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ
Семейства простагландинов, тромбоксанов, лейкотриенов др. -

ПРОСТАГЛАНДИНЫ
Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4).
Ингибируется аспирином по механизму
ПРОСТАГЛАНДИНЫ
Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4).
Ингибируется аспирином по механизму

Внешний обмен липидов
Потребность в жирах – 80 -100 г/сут (возрастает при
Внешний обмен липидов
Потребность в жирах – 80 -100 г/сут (возрастает при

ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Основные эмульгаторы жиров в ЖКТ – желчные кислоты –
ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Основные эмульгаторы жиров в ЖКТ – желчные кислоты –
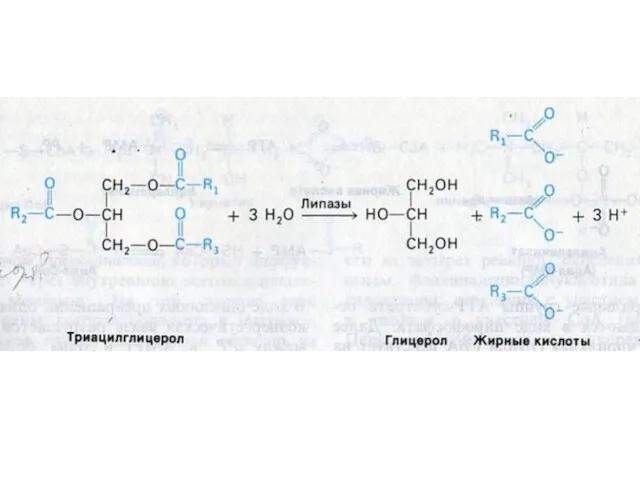
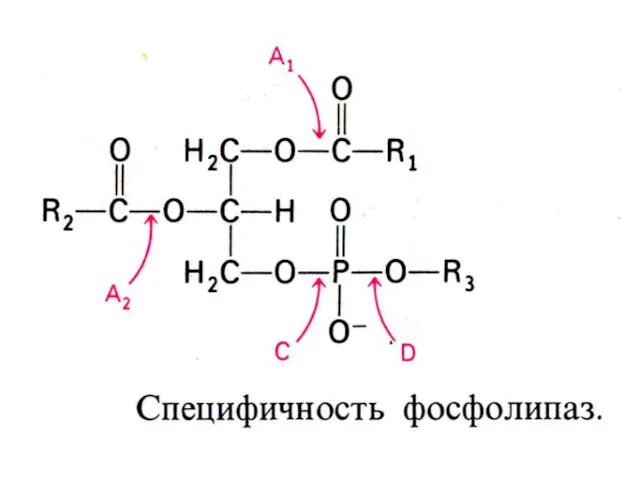

ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Гидролиз нейтральных жиров в 12 –п кишечнике: моно- ,
ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Гидролиз нейтральных жиров в 12 –п кишечнике: моно- ,

ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ
Мелкие эмульгированные капли жира (до 0,5 мкм)
ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ
Мелкие эмульгированные капли жира (до 0,5 мкм)
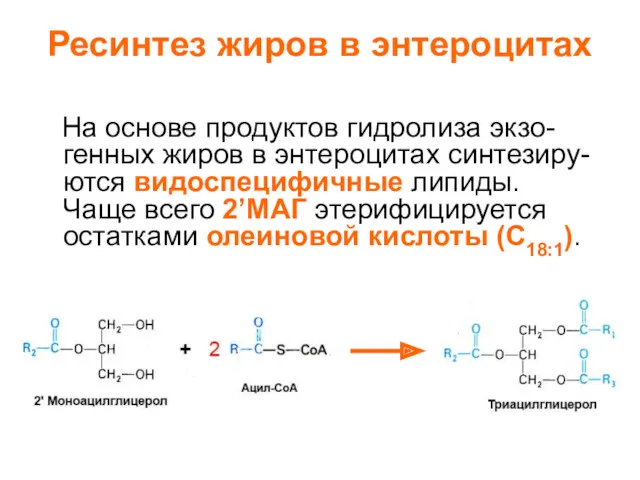
Ресинтез жиров в энтероцитах
На основе продуктов гидролиза экзо- генных жиров
Ресинтез жиров в энтероцитах
На основе продуктов гидролиза экзо- генных жиров
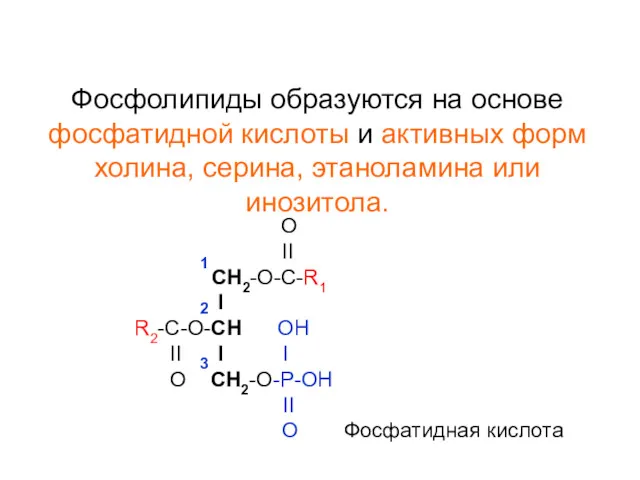
Фосфолипиды образуются на основе фосфатидной кислоты и активных форм холина, серина,
Фосфолипиды образуются на основе фосфатидной кислоты и активных форм холина, серина,

Транспортные формы экзогенных липидов
* Глицерол и СЖК (С<10) выходят из
Транспортные формы экзогенных липидов
* Глицерол и СЖК (С<10) выходят из

ХМ – транспортная форма экзогенных липидов
ХМ образуются в энтероцитах:
* Незрелые ХМ
ХМ – транспортная форма экзогенных липидов
ХМ образуются в энтероцитах:
* Незрелые ХМ
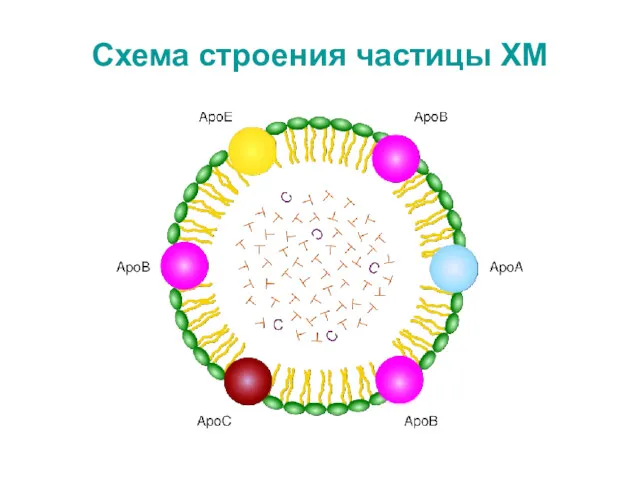
Схема строения частицы ХМ
Схема строения частицы ХМ

* В кровяном русле незрелые ХМ получа-ют от ЛПВП апо-Е, апо-С-II
* В кровяном русле незрелые ХМ получа-ют от ЛПВП апо-Е, апо-С-II
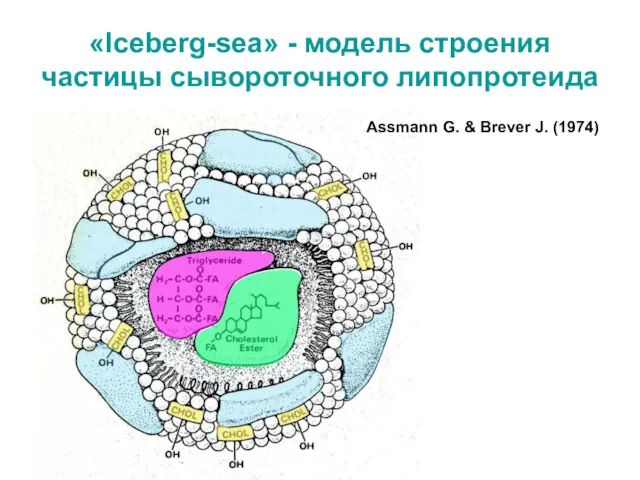
«Iceberg-sea» - модель строения частицы сывороточного липопротеида
Assmann G. & Brever J.
«Iceberg-sea» - модель строения частицы сывороточного липопротеида
Assmann G. & Brever J.

Функции ХМ
* ХМ доставляют экзогенные липиды в печень, жировую ткань, миокард
Функции ХМ
* ХМ доставляют экзогенные липиды в печень, жировую ткань, миокард

ХМ, отдав часть ТАГ в результате их гидро-лиза ЛПЛ, превращаются
ХМ, отдав часть ТАГ в результате их гидро-лиза ЛПЛ, превращаются

Промежуточный обмен липидов
Внутриклеточный липолиз
Адипоциты или клетки жировой ткани (подкожный жир,
Промежуточный обмен липидов
Внутриклеточный липолиз
Адипоциты или клетки жировой ткани (подкожный жир,

Механизм активации гормончувствительной ТАГ-липазы адипоцитов
В постабсорбтивном периоде липолиз в ади-поцитах активируется
Механизм активации гормончувствительной ТАГ-липазы адипоцитов
В постабсорбтивном периоде липолиз в ади-поцитах активируется

* В результате активации аденилатцик-лазы повышается концентрация ц-АМФ, которая активирует протеинкиназу
* В результате активации аденилатцик-лазы повышается концентрация ц-АМФ, которая активирует протеинкиназу
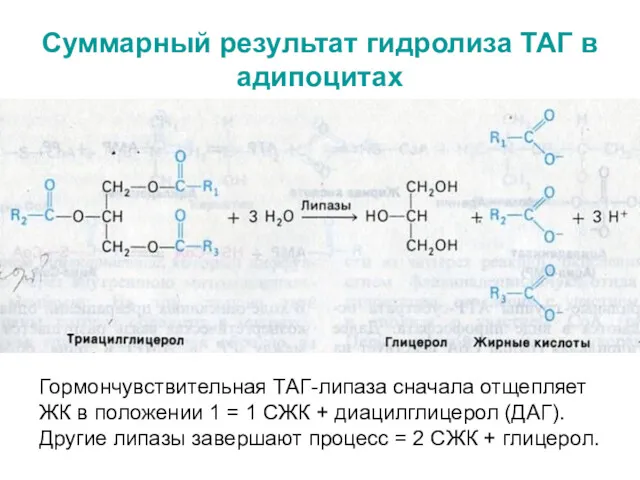
Суммарный результат гидролиза ТАГ в адипоцитах
Гормончувствительная ТАГ-липаза сначала отщепляет
ЖК в
Суммарный результат гидролиза ТАГ в адипоцитах
Гормончувствительная ТАГ-липаза сначала отщепляет
ЖК в
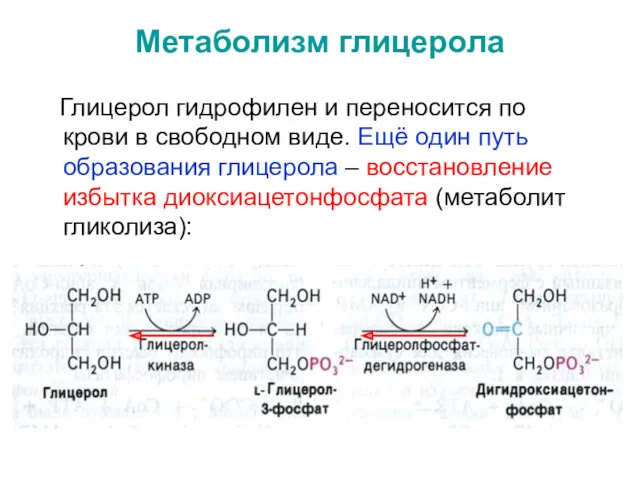
Метаболизм глицерола
Глицерол гидрофилен и переносится по крови в свободном виде.
Метаболизм глицерола
Глицерол гидрофилен и переносится по крови в свободном виде.

* Глицерол является субстратом для:
- липогенеза;
- глюконеогенеза
-
* Глицерол является субстратом для:
- липогенеза;
- глюконеогенеза
-

* СЖК транспортируются по крови в
комплексе с альбумином –
* СЖК транспортируются по крови в
комплексе с альбумином –

Окисление жирных кислот
Путь окисления СЖК, сопряженный с синте-зом АТФ, протекает в
Окисление жирных кислот
Путь окисления СЖК, сопряженный с синте-зом АТФ, протекает в
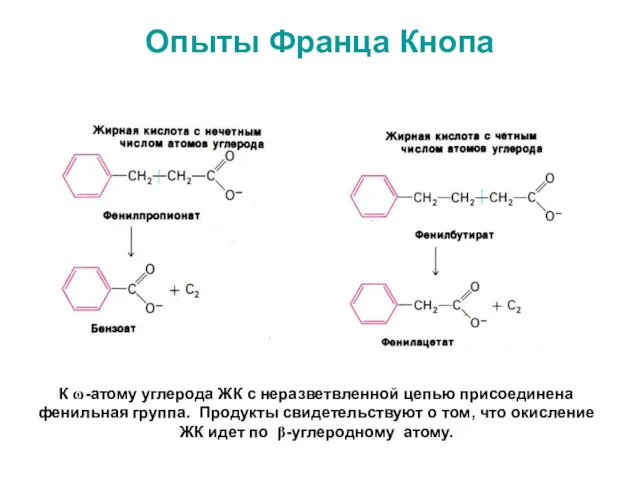
Опыты Франца Кнопа
К ω-атому углерода ЖК с неразветвленной цепью присоединена
фенильная
Опыты Франца Кнопа
К ω-атому углерода ЖК с неразветвленной цепью присоединена
фенильная
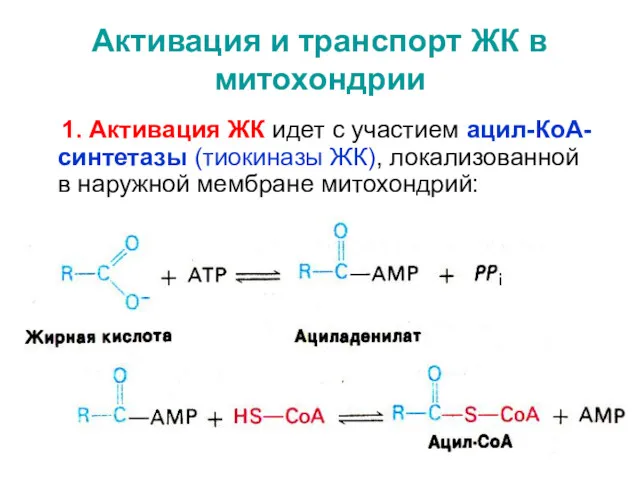
Активация и транспорт ЖК в митохондрии
1. Активация ЖК идет с
Активация и транспорт ЖК в митохондрии
1. Активация ЖК идет с
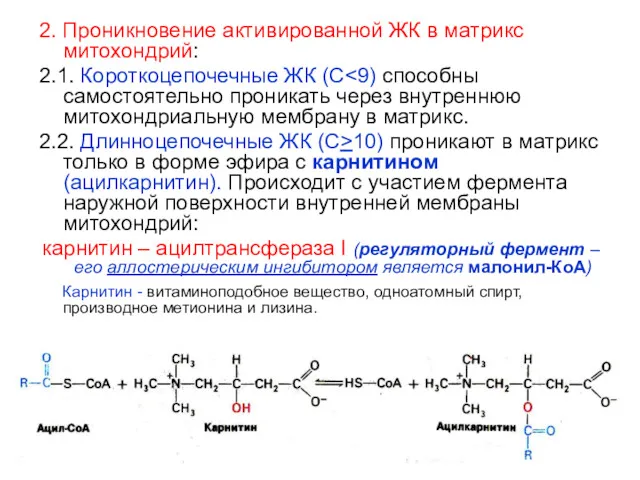
2. Проникновение активированной ЖК в матрикс митохондрий:
2.1. Короткоцепочечные ЖК (С<9) способны
2. Проникновение активированной ЖК в матрикс митохондрий:
2.1. Короткоцепочечные ЖК (С<9) способны

2.3. Обратное превращение:
ацилкарнитин ? ацил-КоА
Происходит с участием фермента,
2.3. Обратное превращение:
ацилкарнитин ? ацил-КоА
Происходит с участием фермента,

Реакции β-окисления жирных кислот
Путь β-окисления – повторяющаяся последова-
тельность четырех реакций.
На
Реакции β-окисления жирных кислот
Путь β-окисления – повторяющаяся последова-
тельность четырех реакций.
На
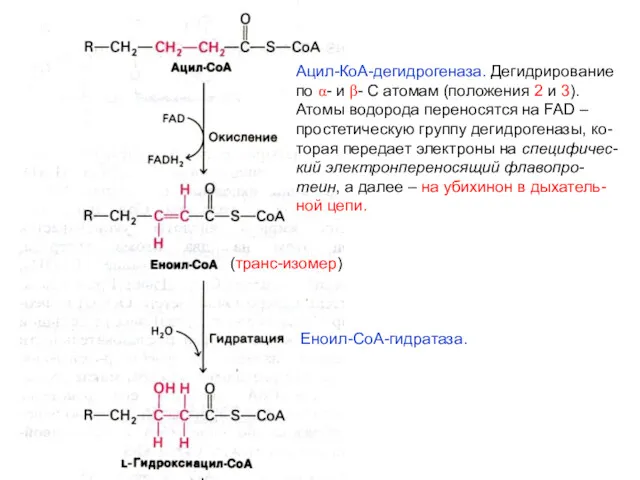
Ацил-КоА-дегидрогеназа. Дегидрирование
по α- и β- С атомам (положения 2 и 3).
Ацил-КоА-дегидрогеназа. Дегидрирование
по α- и β- С атомам (положения 2 и 3).
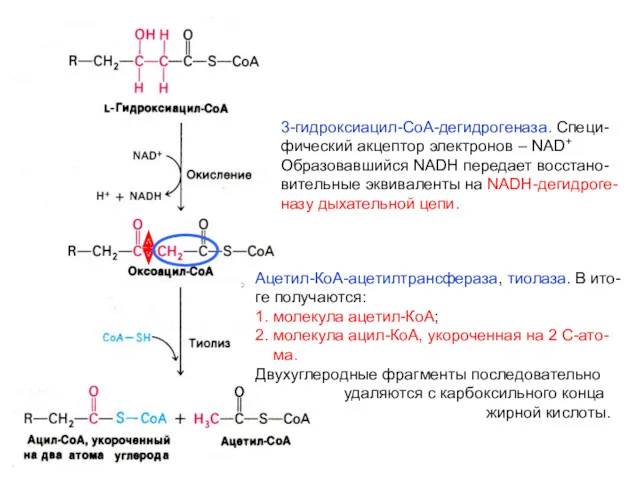
3-гидроксиацил-СоА-дегидрогеназа. Специ-
фический акцептор электронов – NAD+
Образовавшийся NADH передает восстано-
вительные эквиваленты
3-гидроксиацил-СоА-дегидрогеназа. Специ-
фический акцептор электронов – NAD+
Образовавшийся NADH передает восстано-
вительные эквиваленты

Особенности β-окисления ЖК с нечетным числом С-атомов и ненасыщенных ЖК
1.
Особенности β-окисления ЖК с нечетным числом С-атомов и ненасыщенных ЖК
1.

2. Ненасыщенные ЖК (содержат
двойные связи):
Требует участия дополнительных
2. Ненасыщенные ЖК (содержат
двойные связи):
Требует участия дополнительных

Из цис-Δ3-еноил-КоА получается
транс-Δ2-еноил-КоА, который яв-
ляется нормальным субстратом
для еноил-КоА-гидратазы.
Из цис-Δ3-еноил-КоА получается
транс-Δ2-еноил-КоА, который яв-
ляется нормальным субстратом
для еноил-КоА-гидратазы.
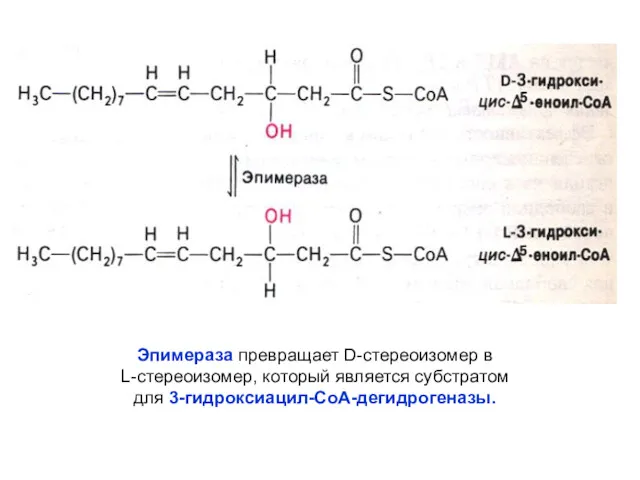
Эпимераза превращает D-стереоизомер в
L-стереоизомер, который является субстратом для 3-гидроксиацил-СоА-дегидрогеназы.
Эпимераза превращает D-стереоизомер в
L-стереоизомер, который является субстратом для 3-гидроксиацил-СоА-дегидрогеназы.

2. Если окисляется ЖК с двумя двойными
связями – линолевая к-та
2. Если окисляется ЖК с двумя двойными
связями – линолевая к-та

Энергетика окисления жирных кислот
Каждый этап β –окисления сопровождается образованием ФАДН2 и
Энергетика окисления жирных кислот
Каждый этап β –окисления сопровождается образованием ФАДН2 и

Ацетил-КоА, как продукт окисления ЖК, далее окисляется в ЦТК («Жиры сгорают
Ацетил-КоА, как продукт окисления ЖК, далее окисляется в ЦТК («Жиры сгорают

Выход АТФ при β-окислении пальмитиновой кислоты
Каждый этап β–окисления сопровождается образованием ФАДН2
Выход АТФ при β-окислении пальмитиновой кислоты
Каждый этап β–окисления сопровождается образованием ФАДН2

КЕТОГЕНЕЗ
Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит
КЕТОГЕНЕЗ
Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит

При голодании и диабете (окисление ЖК усиливается, а глюкозы –
При голодании и диабете (окисление ЖК усиливается, а глюкозы –
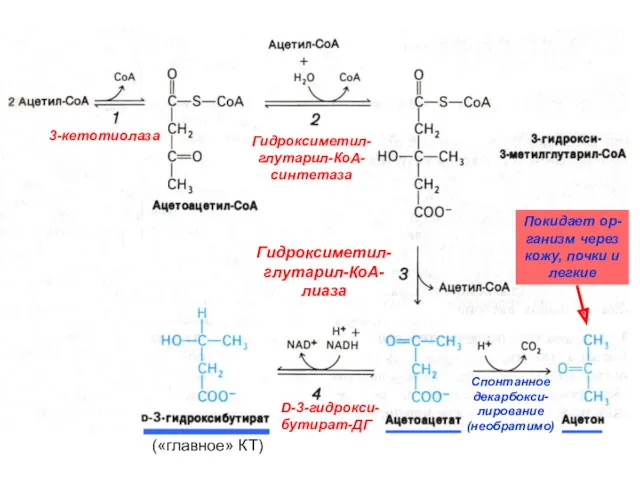
3-кетотиолаза
Гидроксиметил-
глутарил-КоА-
синтетаза
Гидроксиметил-
глутарил-КоА-
лиаза
D-3-гидрокси-
бутират-ДГ
Спонтанное
декарбокси-
лирование
(необратимо)
(«главное» КТ)
Покидает ор-
ганизм через
кожу, почки и
легкие
3-кетотиолаза
Гидроксиметил-
глутарил-КоА-
синтетаза
Гидроксиметил-
глутарил-КоА-
лиаза
D-3-гидрокси-
бутират-ДГ
Спонтанное
декарбокси-
лирование
(необратимо)
(«главное» КТ)
Покидает ор-
ганизм через
кожу, почки и
легкие
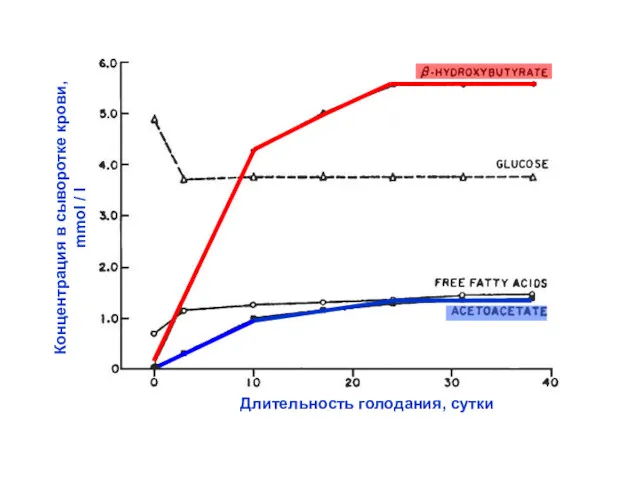
Длительность голодания, сутки
Концентрация в сыворотке крови, mmol / l
Длительность голодания, сутки
Концентрация в сыворотке крови, mmol / l

Ацетоацетат и гидроксибутират свободно диффундируют (по градиенту концентрации) из
гепатоцитов в кровь
Ацетоацетат и гидроксибутират свободно диффундируют (по градиенту концентрации) из
гепатоцитов в кровь

Окисление кетоновых тел
Печень не способна утилизировать кетоновые тела.
В периферических тканях
Окисление кетоновых тел
Печень не способна утилизировать кетоновые тела.
В периферических тканях
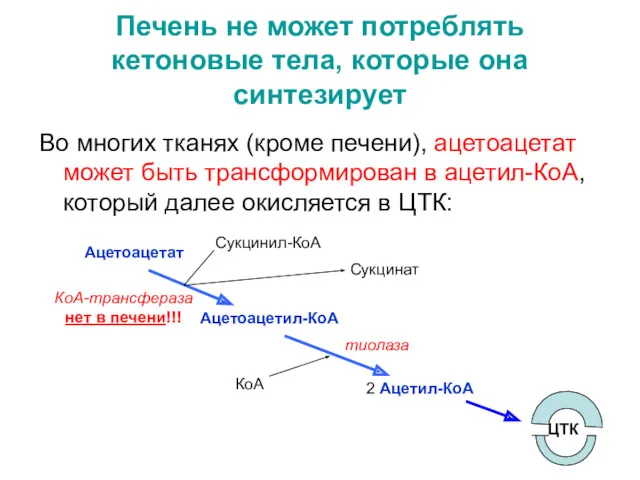
Печень не может потреблять кетоновые тела, которые она синтезирует
Во многих тканях
Печень не может потреблять кетоновые тела, которые она синтезирует
Во многих тканях

Кетоацидоз
При длительном голодании и при диабете концентрация кетоновых тел в крови
Кетоацидоз
При длительном голодании и при диабете концентрация кетоновых тел в крови

Липогенез
Абсорбтивный период. Инсулин.
Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира
Липогенез
Абсорбтивный период. Инсулин.
Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира

ЛИПОГЕНЕЗ
Субстраты:
глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах; диоксиацетонфосфат ДГ (фосфоглицерол ДГ)
ЛИПОГЕНЕЗ
Субстраты:
глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах; диоксиацетонфосфат ДГ (фосфоглицерол ДГ)

Липогенез
Избыток глюкозы обеспечивает:
1. запас гликогена (ограничен)
2. ДАФ ?глицерол-3-фосфат
3. ПВК ? ацетил-КоА
Липогенез
Избыток глюкозы обеспечивает:
1. запас гликогена (ограничен)
2. ДАФ ?глицерол-3-фосфат
3. ПВК ? ацетил-КоА

Биосинтез жирных кислот
Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления! Другая
Биосинтез жирных кислот
Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления! Другая
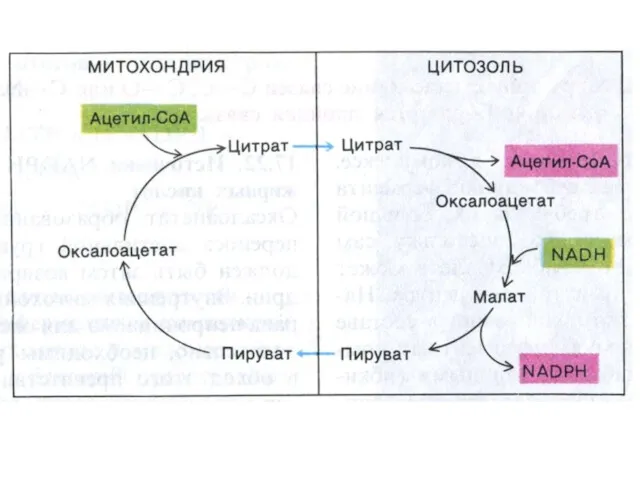

Транспорт ацетил-КоА в цитоплазму
В ситуации накопления АТФ и НАДН ингибируется изоцитрат
Транспорт ацетил-КоА в цитоплазму
В ситуации накопления АТФ и НАДН ингибируется изоцитрат

Транспорт ацетил-КоА в цитоплазму
Возвращение ОА в МХ:
1.ОА –> малат (НАД
Транспорт ацетил-КоА в цитоплазму
Возвращение ОА в МХ:
1.ОА –> малат (НАД
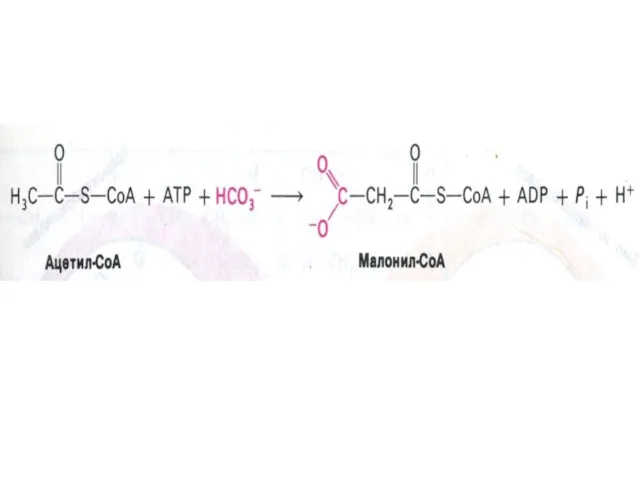

Образование малонил-КоА
ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот, не
Образование малонил-КоА
ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот, не

Синтетаза жирных кислот
мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно).
6 ферментов
Синтетаза жирных кислот
мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно).
6 ферментов
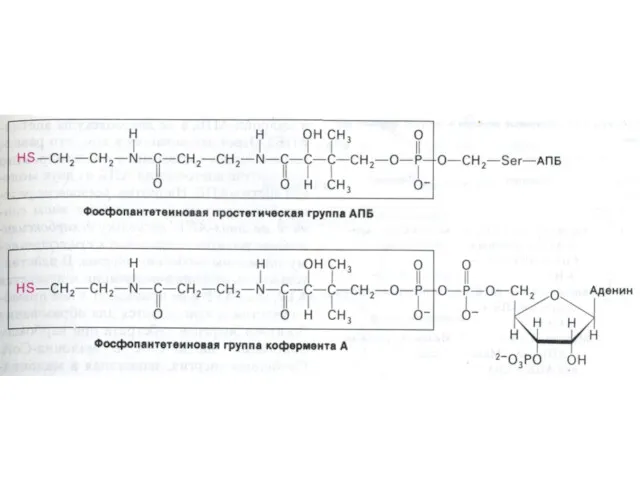

АПБ – структура и функция
Активный центр этого белка – фосфопантотеновая кислота
АПБ – структура и функция
Активный центр этого белка – фосфопантотеновая кислота

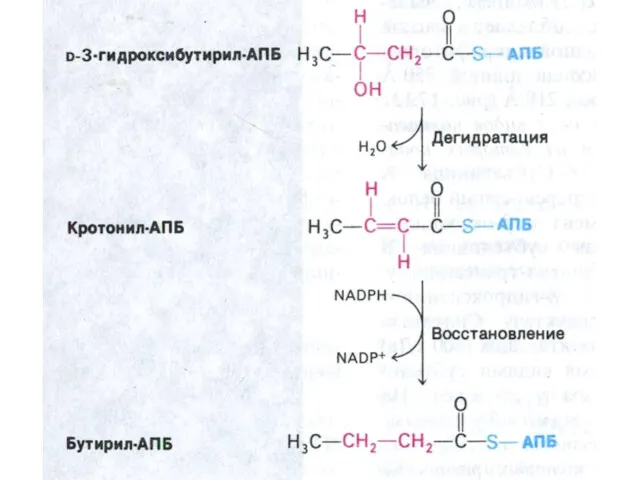

Пальмитоил- синтетаза
Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7
Пальмитоил- синтетаза
Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7

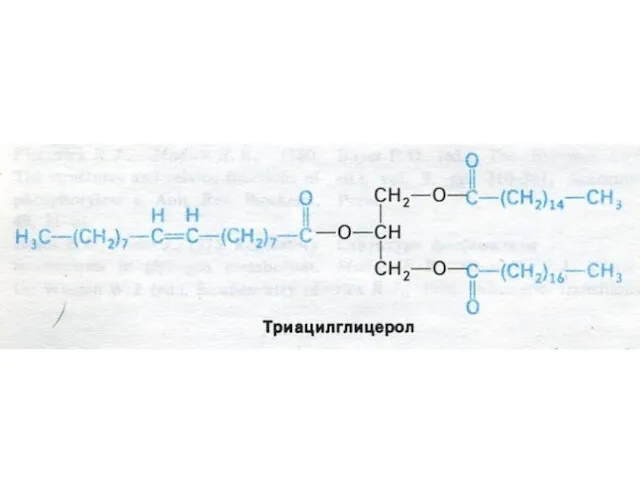
Синтез триацилглицеролов
Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию
Синтез триацилглицеролов
Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию
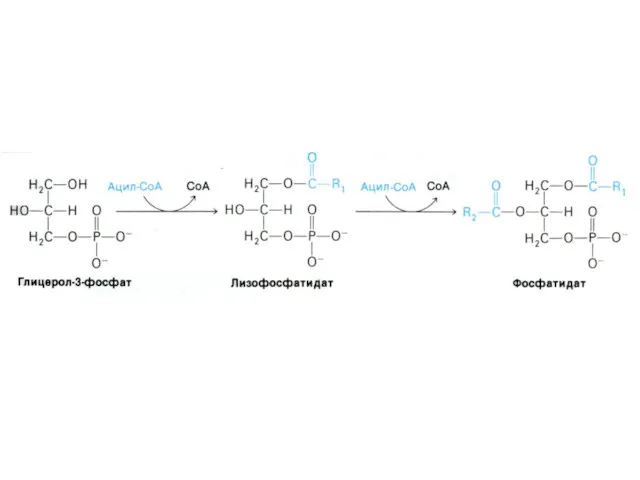
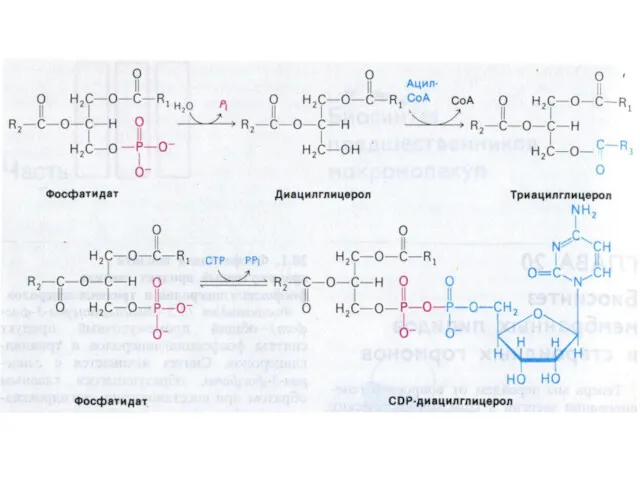

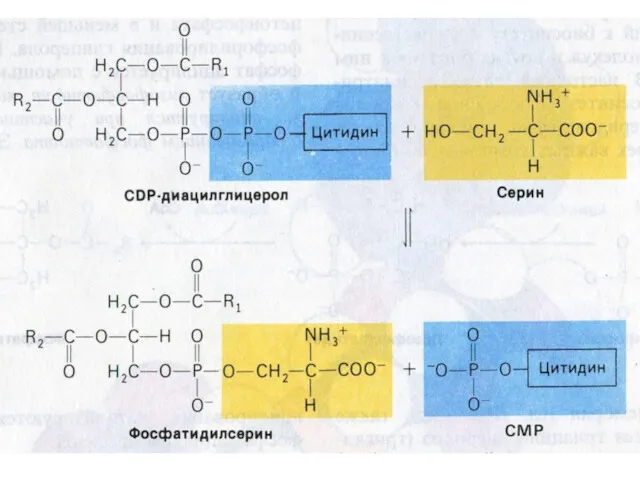
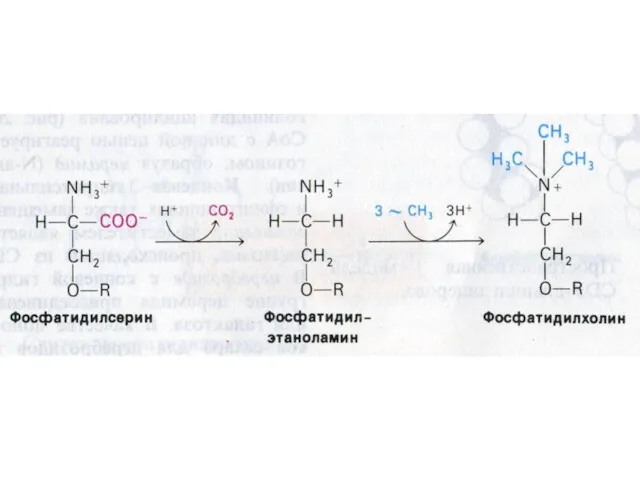
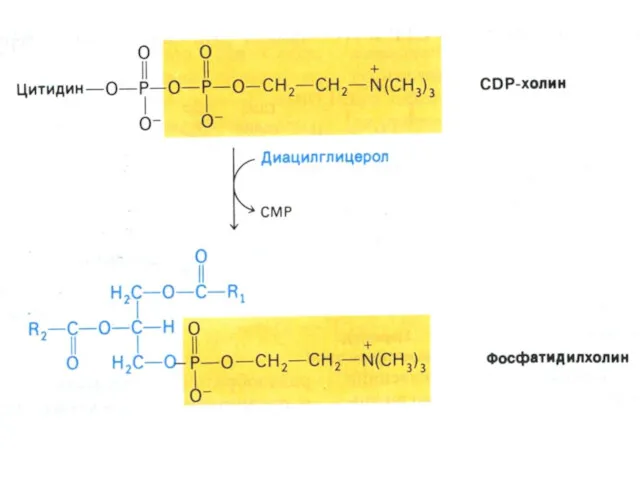
Синтез фосфолипидов
Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного
Синтез фосфолипидов
Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного
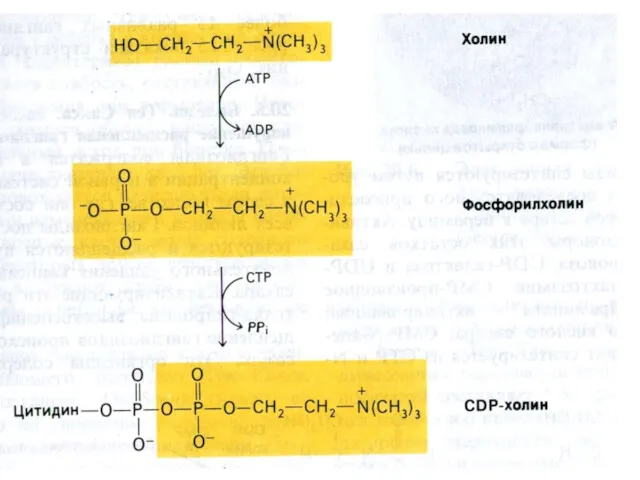

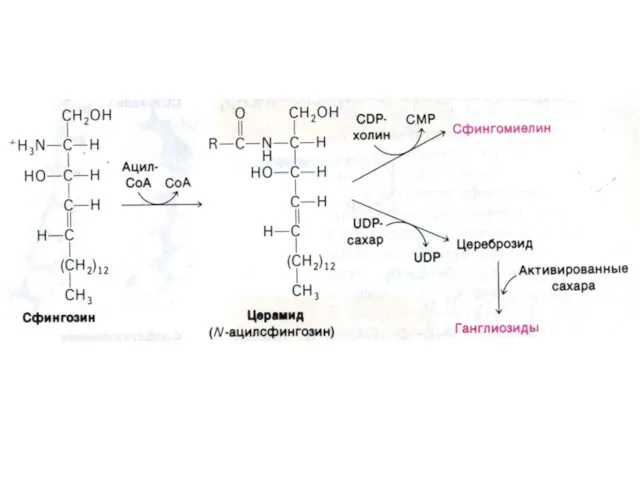
Синтез сфинголипидов
Синтез сфинголипидов

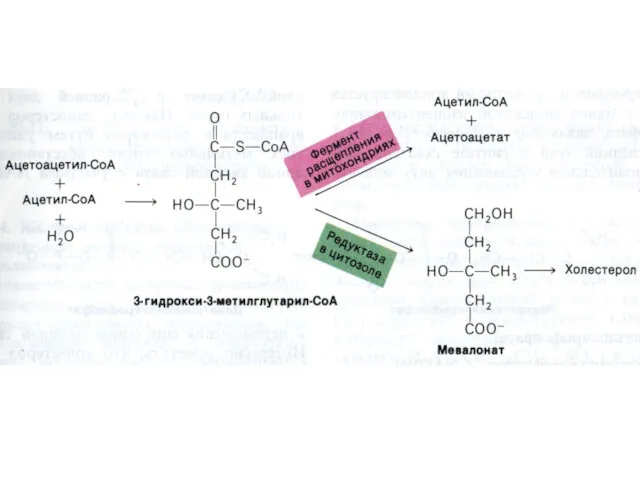
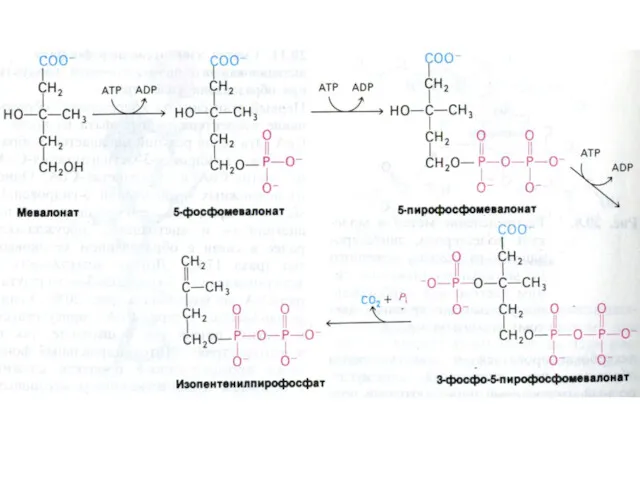
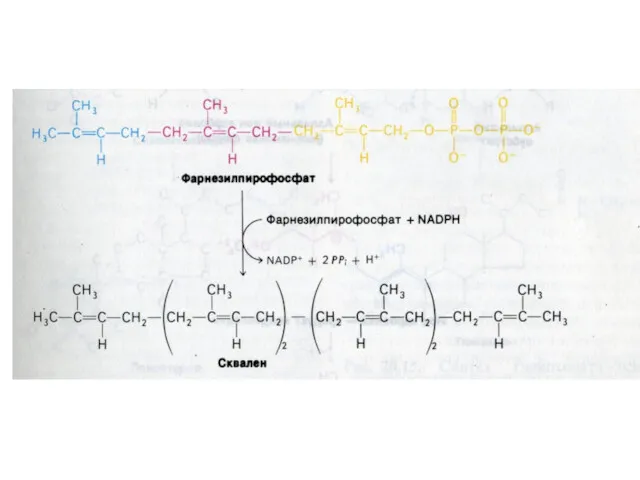
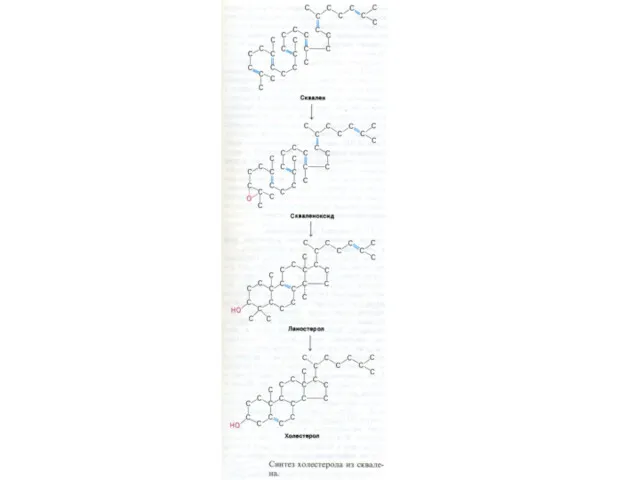
Синтез холестерола
1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА
2. восстановление НАДФН-редуктазой? мевалоновая кислота
3.
Синтез холестерола
1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА
2. восстановление НАДФН-редуктазой? мевалоновая кислота
3.

Метаболизм холестерола
0,5 г эндогенного и 0,5 г экзогенного ХЛ
10% ХЛ находится
Метаболизм холестерола
0,5 г эндогенного и 0,5 г экзогенного ХЛ
10% ХЛ находится


Транспортные формы ХЛ
Экзогенный ХЛ транспортируется в составе хиломикронов, ремнантные формы которых
Транспортные формы ХЛ
Экзогенный ХЛ транспортируется в составе хиломикронов, ремнантные формы которых
 Люминесцентный анализ
Люминесцентный анализ Термодинамика химических процессов
Термодинамика химических процессов Вcтуп до курсу Класифікації отрут і отруєнь. Токсикометрія
Вcтуп до курсу Класифікації отрут і отруєнь. Токсикометрія Полимеры (10 класс)
Полимеры (10 класс) Периодический закон и периодическая система химических элементов
Периодический закон и периодическая система химических элементов Серная кислота и её свойства
Серная кислота и её свойства Индикаторы кислотно-основного титрования
Индикаторы кислотно-основного титрования Высокомолекулярные соединения. Общий курс
Высокомолекулярные соединения. Общий курс Химическая промышленность
Химическая промышленность Альдегіди. Карбонові кислоти. Одержання. Фізичні та хімічні властивості
Альдегіди. Карбонові кислоти. Одержання. Фізичні та хімічні властивості Азотовмісні сполуки
Азотовмісні сполуки Карбон қышқылдары, жіктелуі, сипаттамалары, таралуы
Карбон қышқылдары, жіктелуі, сипаттамалары, таралуы Хімічні властивості кислот
Хімічні властивості кислот Основные этапы первичного процесса переработки нефти
Основные этапы первичного процесса переработки нефти Методика преподавания химии
Методика преподавания химии Отчет по исследовательской работе Образование АСПО
Отчет по исследовательской работе Образование АСПО Закон сохранения массы веществ. Урок вместе с умной колонкой
Закон сохранения массы веществ. Урок вместе с умной колонкой Получение и применение алканов
Получение и применение алканов Розв'язування задач за рівнянням хімічних реакцій
Розв'язування задач за рівнянням хімічних реакцій Кислород/водород. Повторение
Кислород/водород. Повторение История открытия химических веществ
История открытия химических веществ Некоторые структурные особенности макромолекул
Некоторые структурные особенности макромолекул 20231009_zhyostkost_vody
20231009_zhyostkost_vody Сложные эфиры. 10 класс
Сложные эфиры. 10 класс Элементтер-органогендер. Көміртек – органикалық қосылыстардың негізін құраушы. Тірі және өлі табиғат арасындағы шекара
Элементтер-органогендер. Көміртек – органикалық қосылыстардың негізін құраушы. Тірі және өлі табиғат арасындағы шекара Железо и его соединения
Железо и его соединения Особенности сжигания жидкого топлива и топливосжигающие устройства
Особенности сжигания жидкого топлива и топливосжигающие устройства Алкандар. Метан молекуласының құрылысы Метан және оның гомологтарының қасиеттері
Алкандар. Метан молекуласының құрылысы Метан және оның гомологтарының қасиеттері