- Нуклеопротеины

Содержание
- 2. Устойчивость нуклеопротеиновых комплексов обеспечивается нековалентным взаимодействием. У различных нуклеопротеинов в обеспечение стабильности комплекса вносят вклад различные
- 3. Примером специфичного взаимодействия могут служить нуклеопротеидные комплексы рРНК — субъединицы рибосом; неспецифичное электростатическое взаимодействие характерно для
- 4. Наличие отрицательно заряженного фосфата в каждом нуклеотиде делает НК полианионами. Поэтому с белками они образуют солеподобные
- 5. В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК).
- 6. Каждый нуклеотид в свою очередь состоит из трех компонентов: азотистого основания, являющегося производным пурина или пиримидина,
- 7. Свойства азотистых оснований: —плохо растворимы в воде (гидрофобны); — плоскостные (копланарные); — поглощают ультрафиолет при 260
- 9. В нуклеиновые кислоты входят два вида пентоз: β-D-рибоза в РНК и β-D-2-дезоксирибоза в молекулу ДНК АО
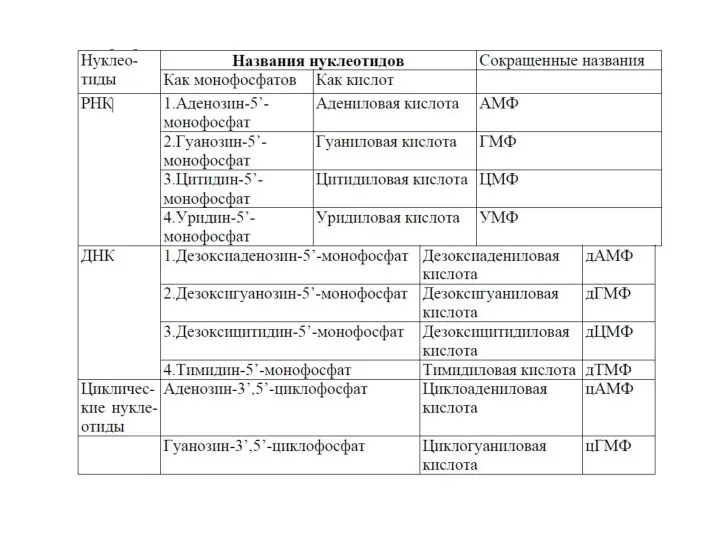
- 10. Свойства: —несут отрицательный заряд — проявляют кислотные свойства Номенклатура нуклеотидов: нуклеозид-5´-монофосфат, нуклеозид-5´-дифосфат, нуклеозид-5´-трифосфат. Строение АТФ Строение
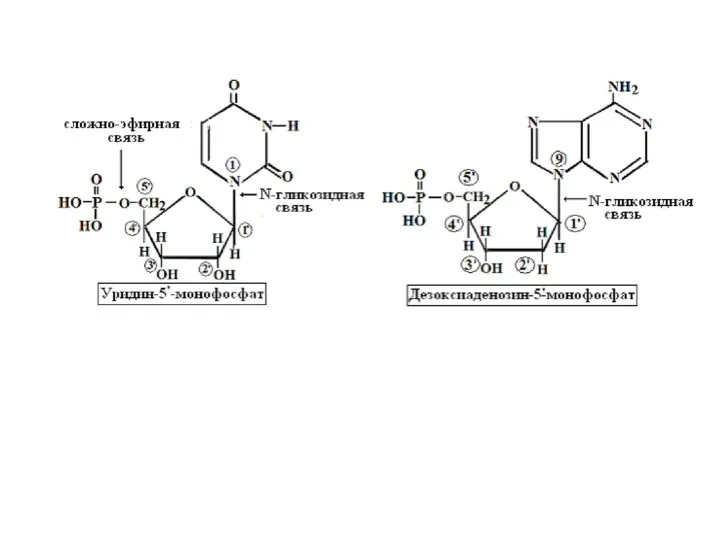
- 11. Образование названий нуклеозидов и нуклеотидов аденозин-5`-монофосфат или адениловая кислота или АМФ аденин аденозин гуанин цитозин урацил
- 14. Известны также циклические нуклеотиды, в которых фосфорная кислота образует сложноэфирные связи одновременно с 5’ и 3’-атомами
- 15. Гуанозин-5’-дифосфат-3’-дифосфат —внутриклеточный регулятор метаболизма бактерий
- 17. Соединение двух нуклеотидов в динуклеотид 3’,5’- фосфодиэфирной связью
- 18. Первичная структура нуклеиновых кислот - это порядок чередования нуклеотидов, связанных друг с другом в линейной последовательности
- 19. Первичная структура нуклеиновых кислот Х = Н для ДНК, Х = ОН для РНК Связи в
- 20. Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева
- 21. Пространственная структура ДНК
- 22. Вторичная структура ДНК представлена двойной спиралью, в которой две полинуклеотидные цепи расположены антипараллельно и удерживаются относительно
- 23. Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются
- 24. наибольшее перекрывание наименьшее перекрывание Комплементарные основания обращены внутрь молекулы, лежат в одной плоскости, которая практически перпендикулярна
- 25. Существует несколько форм правозакрученной двойной спирали ДНК. В клетке ДНК чаще всего находится в В-форме, в
- 26. Третичная структура ДНК формируется при ее взаимодействии с белками. Каждая молекула ДНК упакована в отдельную хромосому,
- 27. Гистоны - это небольшие белки с высоким содержанием положительно заряженных аминокислот лизина и аргинина. Они взаимодействуют
- 28. Взаимодействие гистонов и ДНК Участок суперспирали ДНК
- 29. Пространственная структура РНК
- 30. Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК. В спирализованных участках или шпильках
- 31. Минорные основания входят в состав 10% от всех нуклеотидов. Обнаружено до 50 разновидностей. Встречаются в т-РНК,
- 32. Типы клеточной РНК в зависимости от функций.
- 33. Транспортные РНК (тРНК) являются молекулами-адапторами, у которых к 3'-концу присоединяется аминокислота, а участок антикодона - к
- 36. Рибосомные РНК (рРНК) составляют около 80% всей РНК клетки и входят в состав рибосом. В цитоплазматические
- 37. Отличия между РНК и ДНК: — количество цепей: в РНК одна цепь, в ДНК две цепи,
- 38. Таким образом, нуклеотиды, мононуклеотиды и олигонуклеотиды присутствуют в цитоплазме клеток и ее органеллах, выполняя определенные функции.
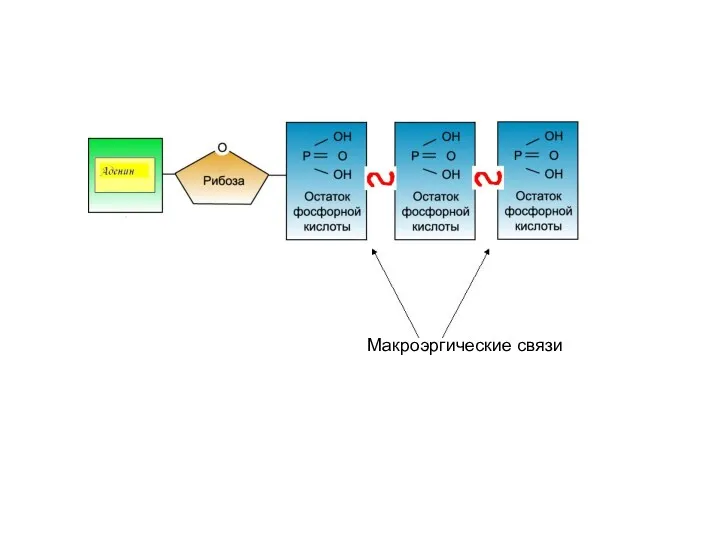
- 42. 2. Энергетическая. Макроэргические молекулы (макроэрги) — биологические молекулы, которые способны накапливать и передавать энергию в ходе
- 44. При отщеплении от молекулы АТФ (ГТФ и т.д.) одного или двух остатков фосфорной кислоты образуется соответственно
- 45. Наличие макроэргических связей в нуклеотидах позволяет им являться активаторами и переносчиками мономеров в клетке: — УТФ
- 47. Скачать презентацию

Устойчивость нуклеопротеиновых комплексов обеспечивается нековалентным взаимодействием. У различных нуклеопротеинов в обеспечение
Устойчивость нуклеопротеиновых комплексов обеспечивается нековалентным взаимодействием. У различных нуклеопротеинов в обеспечение
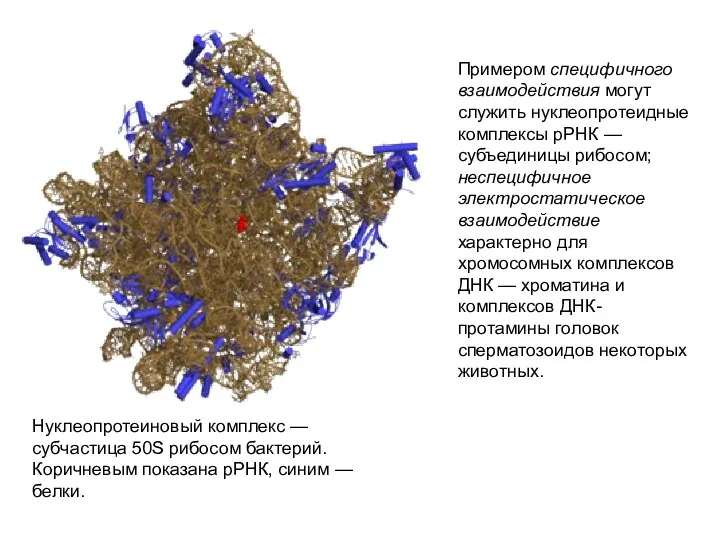
Примером специфичного взаимодействия могут служить нуклеопротеидные комплексы рРНК — субъединицы рибосом;
Примером специфичного взаимодействия могут служить нуклеопротеидные комплексы рРНК — субъединицы рибосом;
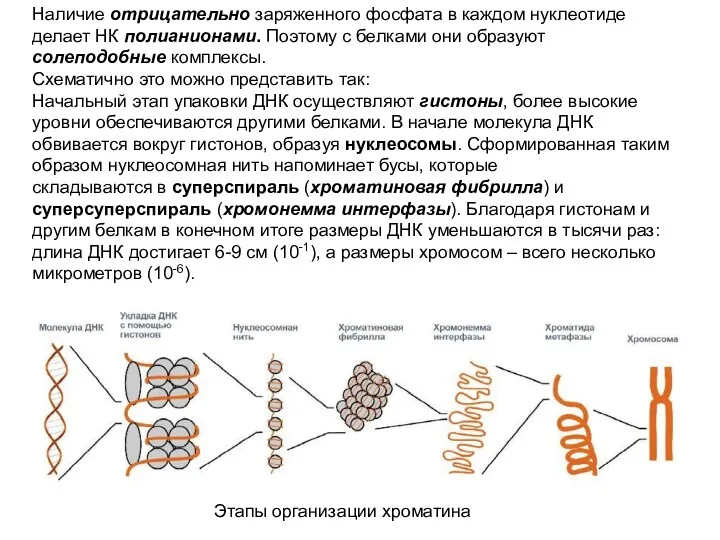
Наличие отрицательно заряженного фосфата в каждом нуклеотиде делает НК полианионами. Поэтому
Наличие отрицательно заряженного фосфата в каждом нуклеотиде делает НК полианионами. Поэтому

В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота
В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота
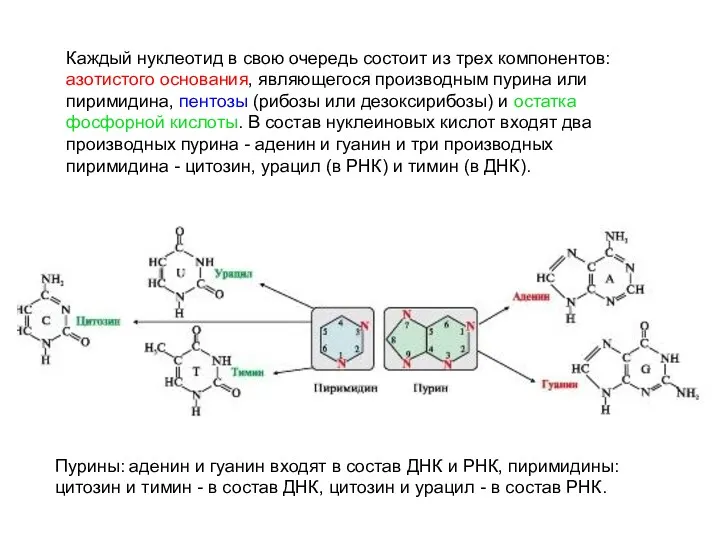
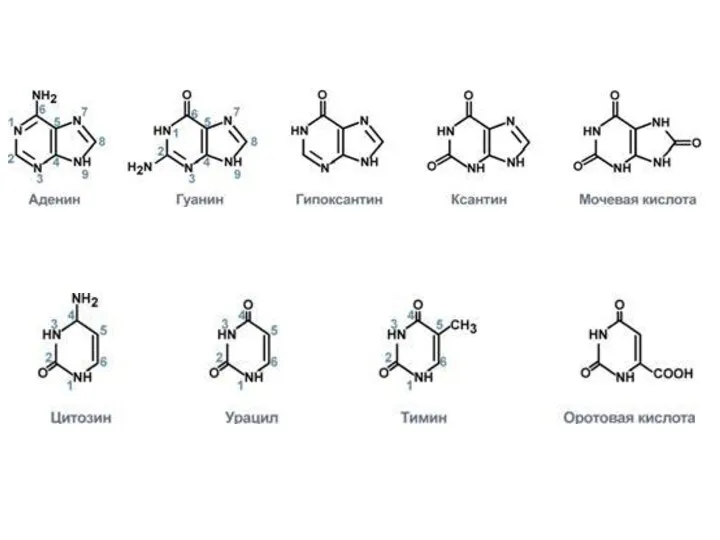
Каждый нуклеотид в свою очередь состоит из трех компонентов: азотистого основания,
Каждый нуклеотид в свою очередь состоит из трех компонентов: азотистого основания,

Свойства азотистых оснований:
—плохо растворимы в воде (гидрофобны);
— плоскостные (копланарные);
— поглощают ультрафиолет
Свойства азотистых оснований:
—плохо растворимы в воде (гидрофобны);
— плоскостные (копланарные);
— поглощают ультрафиолет
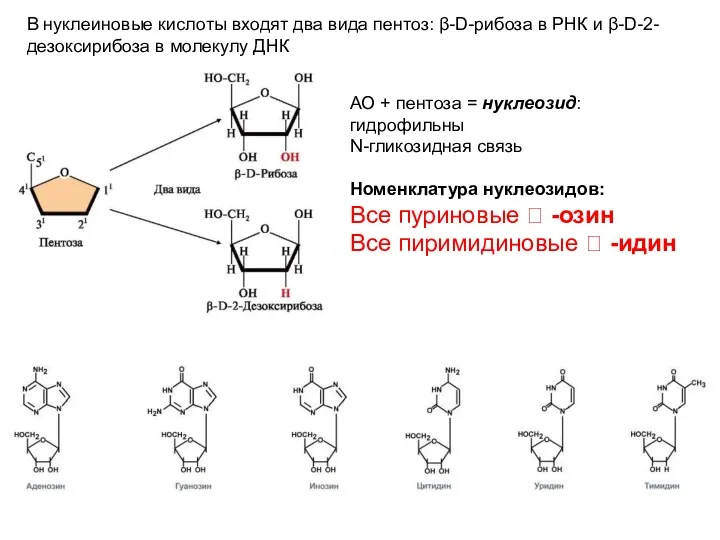
В нуклеиновые кислоты входят два вида пентоз: β-D-рибоза в РНК и
В нуклеиновые кислоты входят два вида пентоз: β-D-рибоза в РНК и
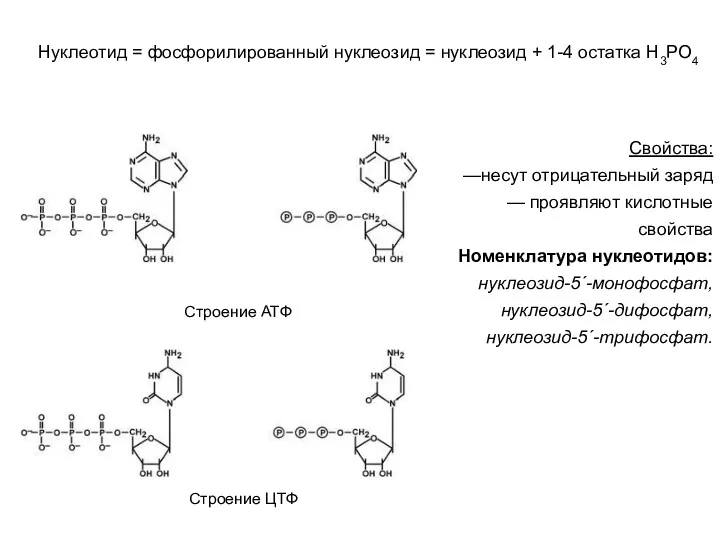
Свойства:
—несут отрицательный заряд
— проявляют кислотные свойства
Номенклатура нуклеотидов:
нуклеозид-5´-монофосфат, нуклеозид-5´-дифосфат, нуклеозид-5´-трифосфат.
Строение АТФ
Строение ЦТФ
Нуклеотид
Свойства:
—несут отрицательный заряд
— проявляют кислотные свойства
Номенклатура нуклеотидов:
нуклеозид-5´-монофосфат, нуклеозид-5´-дифосфат, нуклеозид-5´-трифосфат.
Строение АТФ
Строение ЦТФ
Нуклеотид
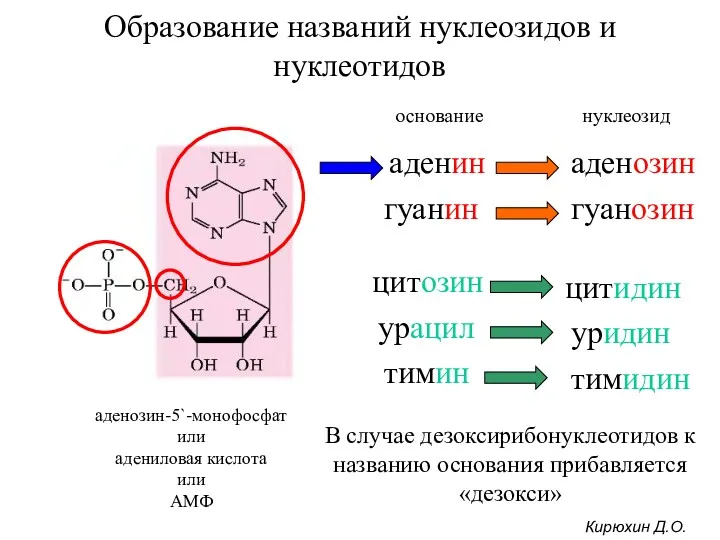
Образование названий нуклеозидов и нуклеотидов
аденозин-5`-монофосфат
или
адениловая кислота
или
АМФ
аденин
аденозин
гуанин
цитозин
урацил
тимин
гуанозин
цитидин
уридин
тимидин
В случае дезоксирибонуклеотидов к названию
Образование названий нуклеозидов и нуклеотидов
аденозин-5`-монофосфат
или
адениловая кислота
или
АМФ
аденин
аденозин
гуанин
цитозин
урацил
тимин
гуанозин
цитидин
уридин
тимидин
В случае дезоксирибонуклеотидов к названию
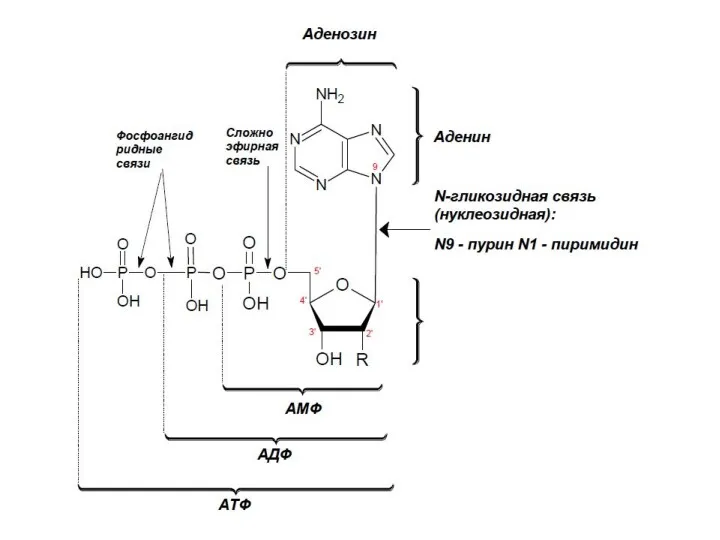
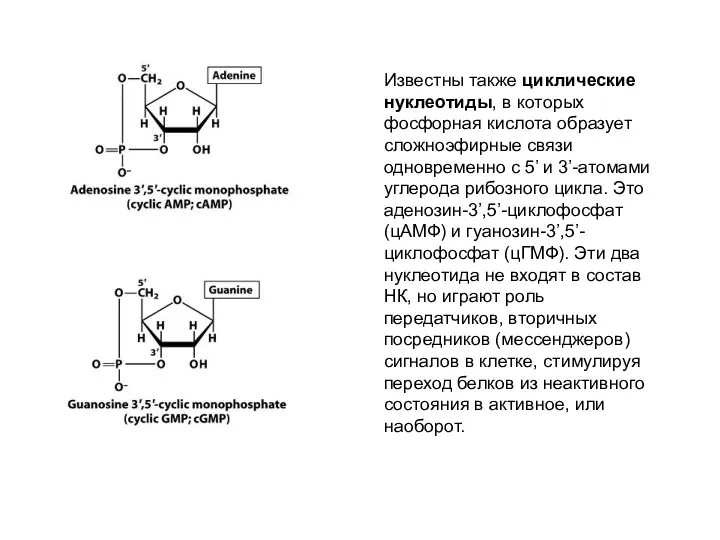
Известны также циклические нуклеотиды, в которых фосфорная кислота образует сложноэфирные связи
Известны также циклические нуклеотиды, в которых фосфорная кислота образует сложноэфирные связи
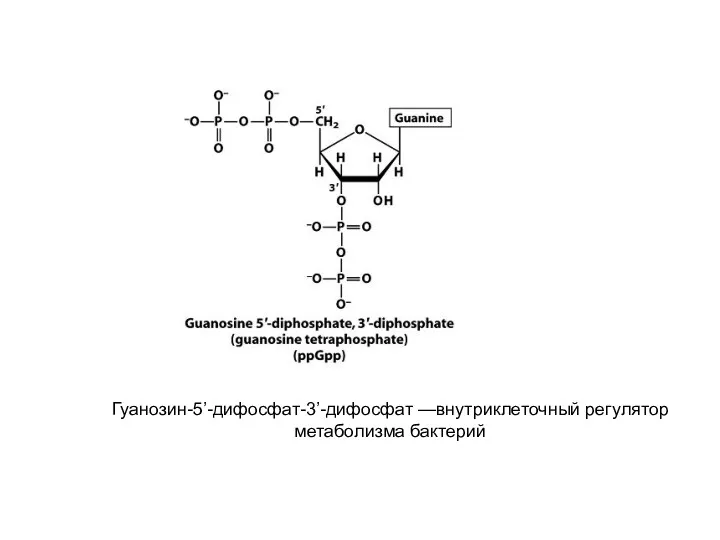
Гуанозин-5’-дифосфат-3’-дифосфат —внутриклеточный регулятор метаболизма бактерий
Гуанозин-5’-дифосфат-3’-дифосфат —внутриклеточный регулятор метаболизма бактерий
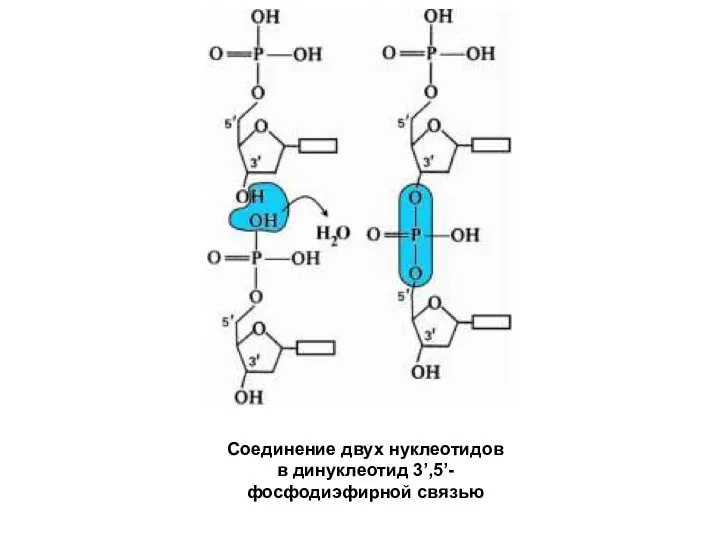
Соединение двух нуклеотидов
в динуклеотид 3’,5’- фосфодиэфирной связью
Соединение двух нуклеотидов
в динуклеотид 3’,5’- фосфодиэфирной связью
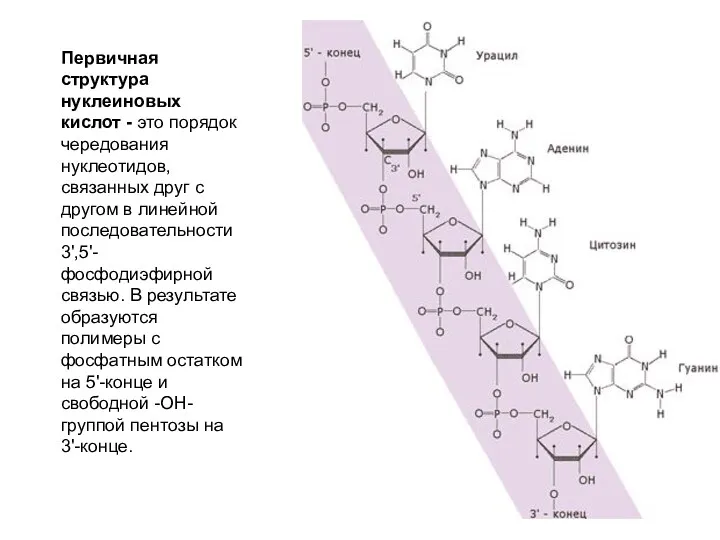
Первичная структура нуклеиновых кислот - это порядок чередования нуклеотидов, связанных друг
Первичная структура нуклеиновых кислот - это порядок чередования нуклеотидов, связанных друг
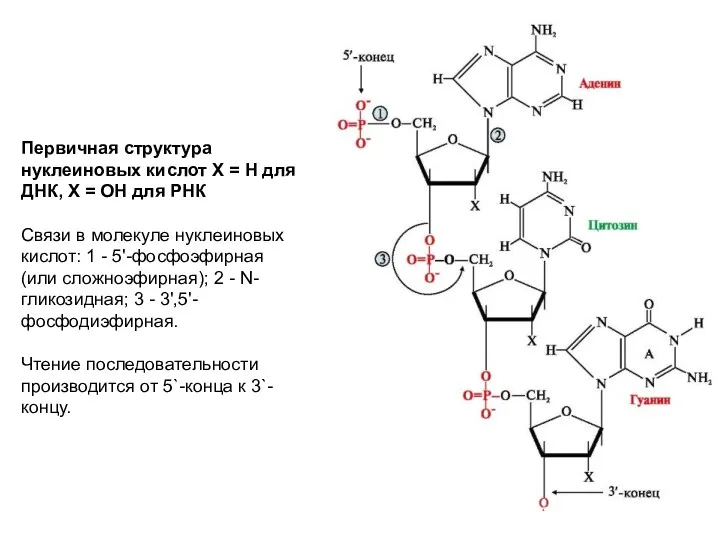
Первичная структура нуклеиновых кислот Х = Н для ДНК, Х =
Первичная структура нуклеиновых кислот Х = Н для ДНК, Х =

Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом.
Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом.

Пространственная структура ДНК
Пространственная структура ДНК
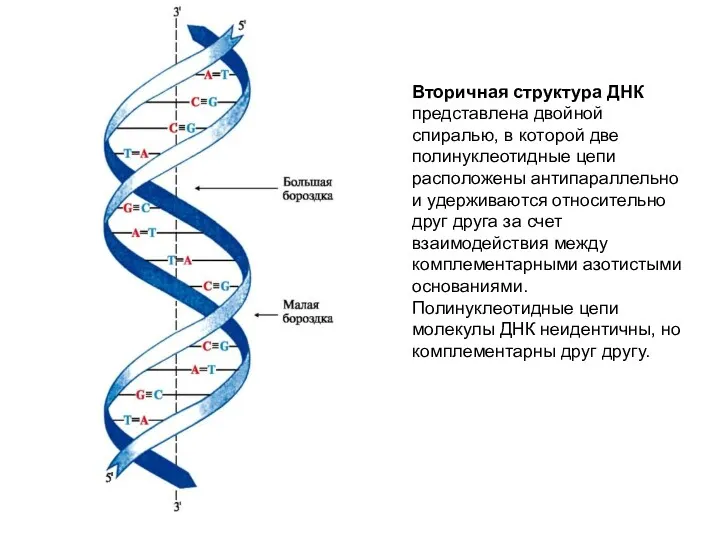
Вторичная структура ДНК представлена двойной спиралью, в которой две полинуклеотидные цепи
Вторичная структура ДНК представлена двойной спиралью, в которой две полинуклеотидные цепи
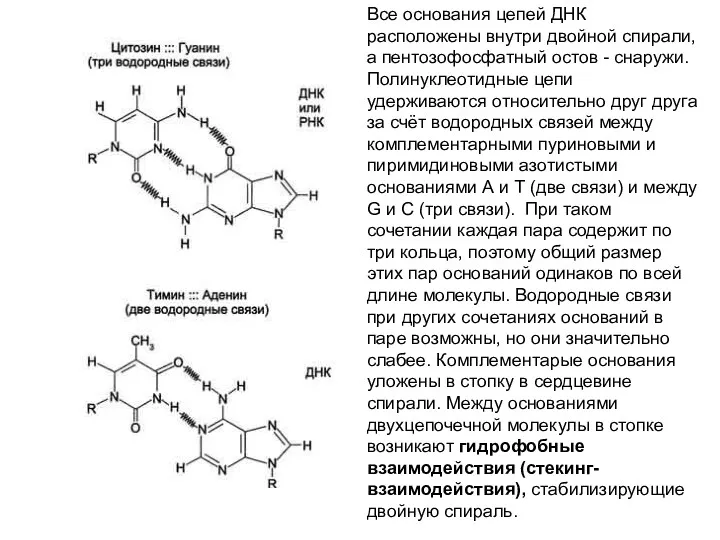
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов
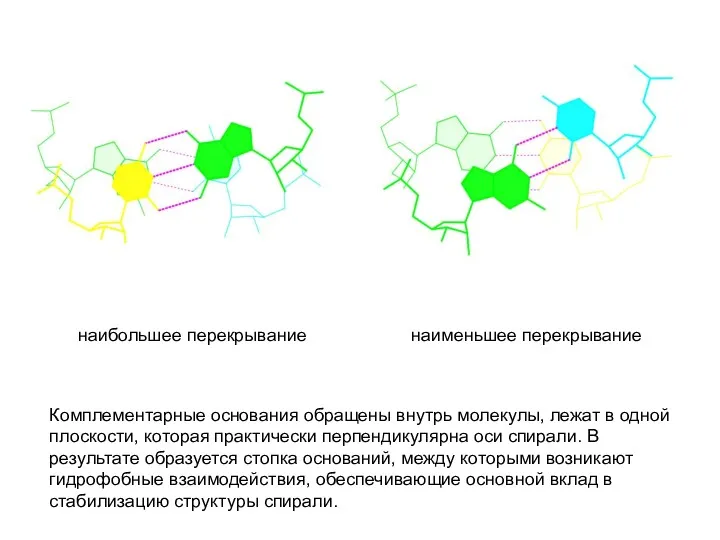
наибольшее перекрывание наименьшее перекрывание
Комплементарные основания обращены внутрь молекулы, лежат в одной
наибольшее перекрывание наименьшее перекрывание
Комплементарные основания обращены внутрь молекулы, лежат в одной
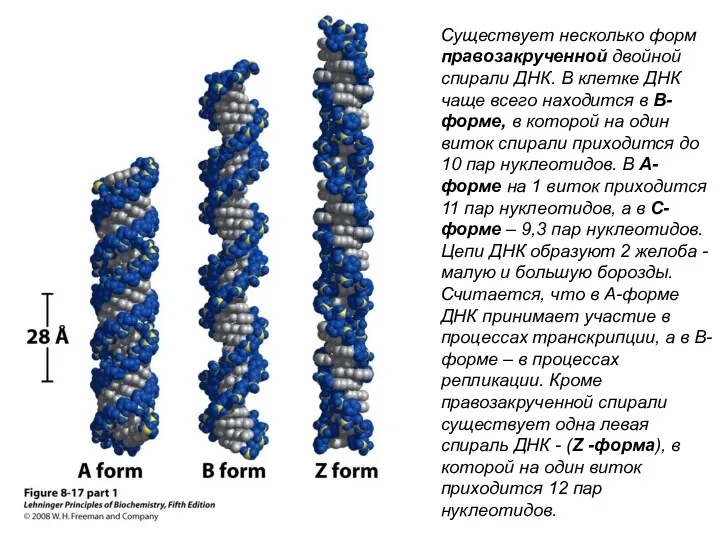
Существует несколько форм правозакрученной двойной спирали ДНК. В клетке ДНК чаще
Существует несколько форм правозакрученной двойной спирали ДНК. В клетке ДНК чаще

Третичная структура ДНК формируется при ее взаимодействии с белками. Каждая молекула
Третичная структура ДНК формируется при ее взаимодействии с белками. Каждая молекула
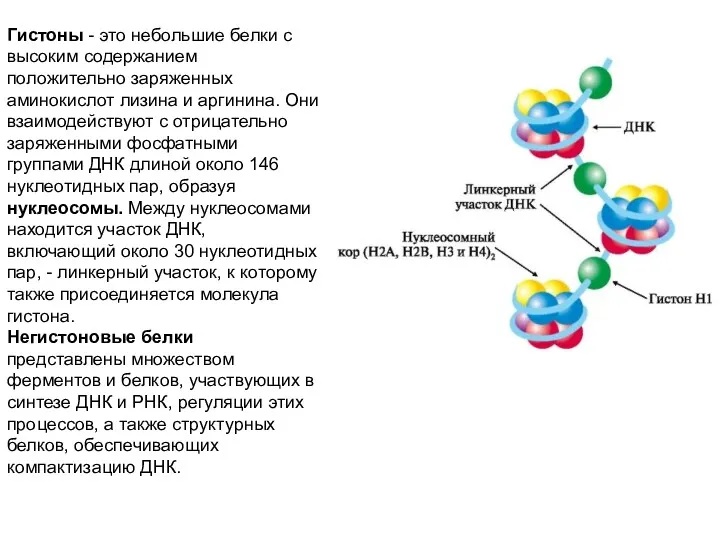
Гистоны - это небольшие белки с высоким содержанием положительно заряженных аминокислот
Гистоны - это небольшие белки с высоким содержанием положительно заряженных аминокислот
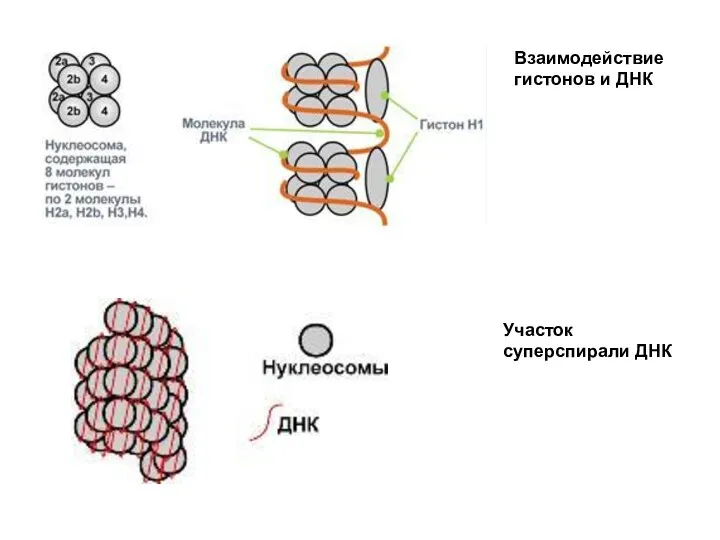
Взаимодействие гистонов и ДНК
Участок суперспирали ДНК
Взаимодействие гистонов и ДНК
Участок суперспирали ДНК

Пространственная структура РНК
Пространственная структура РНК

Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК.
Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК.
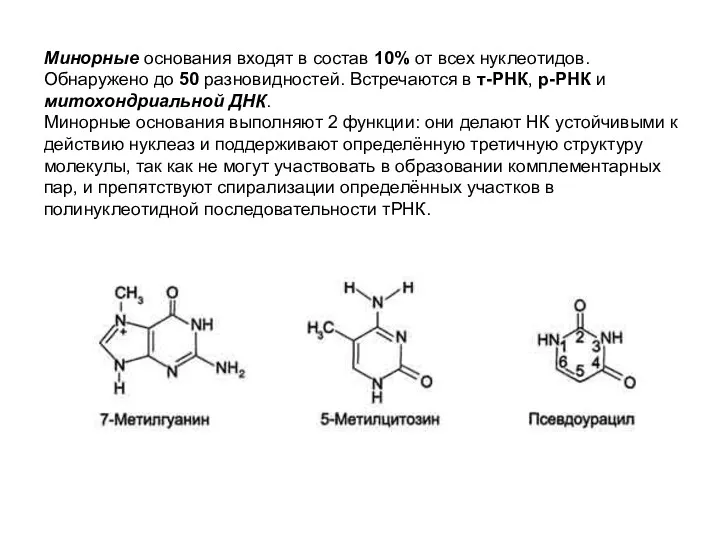
Минорные основания входят в состав 10% от всех нуклеотидов.
Обнаружено до 50
Минорные основания входят в состав 10% от всех нуклеотидов.
Обнаружено до 50
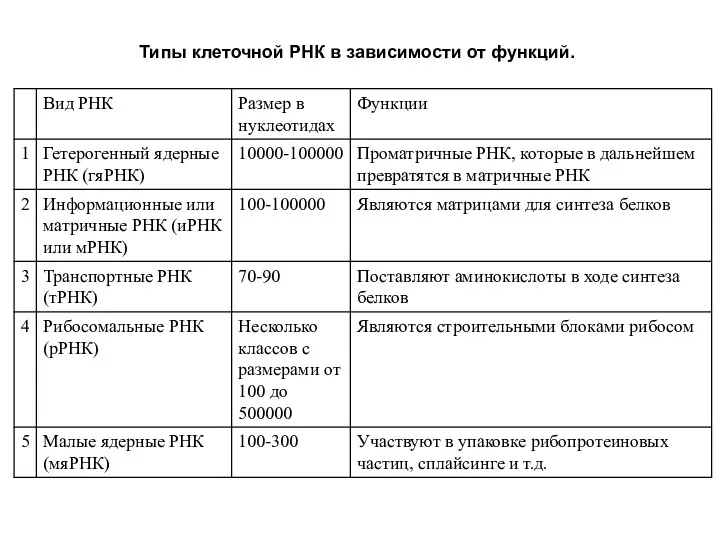
Типы клеточной РНК в зависимости от функций.
Типы клеточной РНК в зависимости от функций.

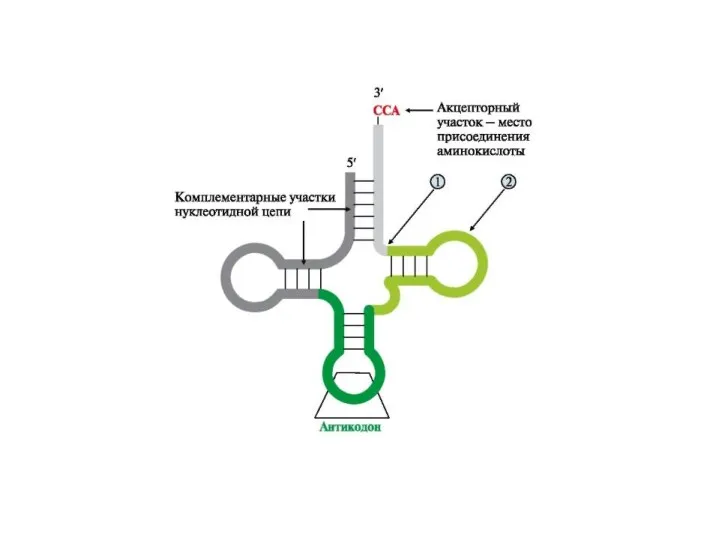
Транспортные РНК (тРНК) являются молекулами-адапторами, у которых к 3'-концу присоединяется аминокислота,
Транспортные РНК (тРНК) являются молекулами-адапторами, у которых к 3'-концу присоединяется аминокислота,
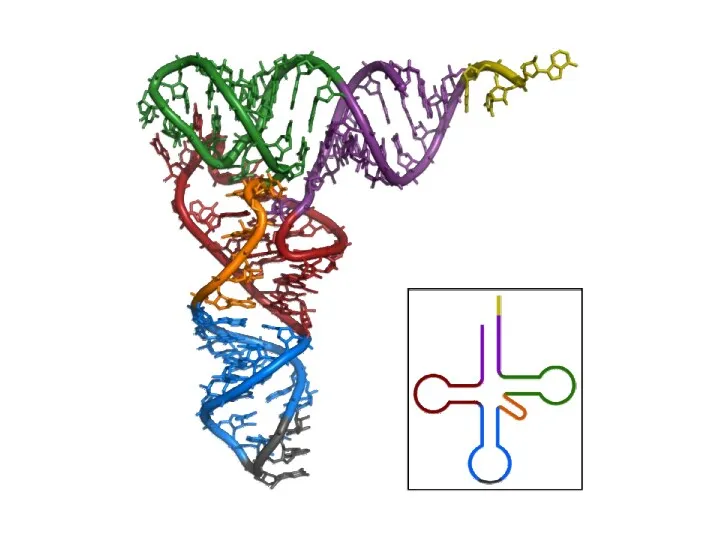

Рибосомные РНК (рРНК) составляют около 80% всей РНК клетки и входят
Рибосомные РНК (рРНК) составляют около 80% всей РНК клетки и входят

Отличия между РНК и ДНК:
— количество цепей: в РНК одна цепь,
— количество цепей: в РНК одна цепь,

Таким образом, нуклеотиды, мононуклеотиды и олигонуклеотиды присутствуют в цитоплазме клеток и
Таким образом, нуклеотиды, мононуклеотиды и олигонуклеотиды присутствуют в цитоплазме клеток и
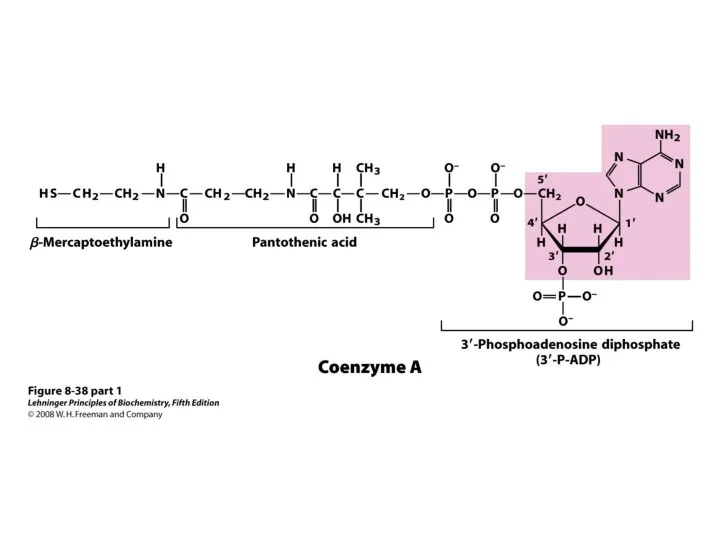
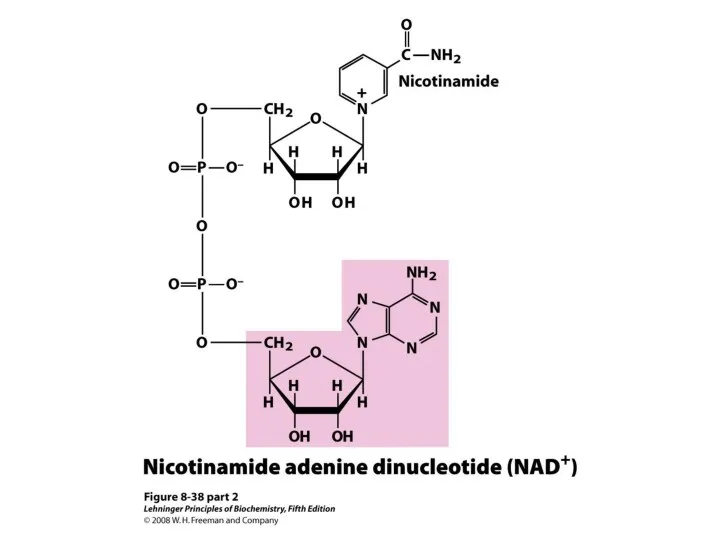
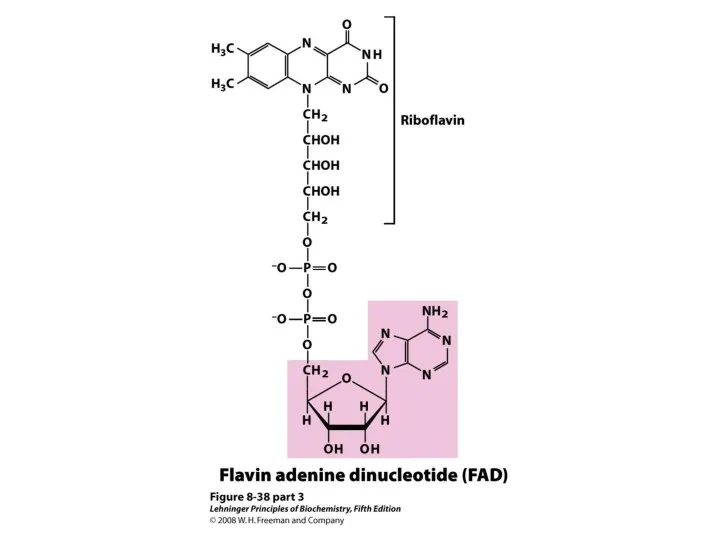

2. Энергетическая.
Макроэргические молекулы (макроэрги) — биологические молекулы, которые способны накапливать
2. Энергетическая.
Макроэргические молекулы (макроэрги) — биологические молекулы, которые способны накапливать
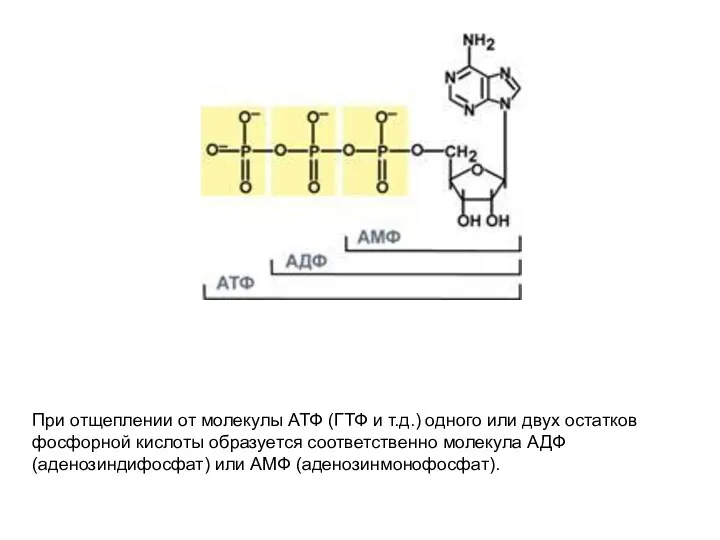
При отщеплении от молекулы АТФ (ГТФ и т.д.) одного или двух
При отщеплении от молекулы АТФ (ГТФ и т.д.) одного или двух

Наличие макроэргических связей в нуклеотидах позволяет им являться активаторами и переносчиками
Наличие макроэргических связей в нуклеотидах позволяет им являться активаторами и переносчиками
 Оксид серы (IV). Сернистая кислота
Оксид серы (IV). Сернистая кислота Подготовка к ГИА. В2. Первоначальные сведения об органических веществах. Биологически важные вещества: белки, жиры, углеводы
Подготовка к ГИА. В2. Первоначальные сведения об органических веществах. Биологически важные вещества: белки, жиры, углеводы Строение атома. 10 ен
Строение атома. 10 ен Химия и повседневная жизнь человека
Химия и повседневная жизнь человека Ароматические кислоты и их соли
Ароматические кислоты и их соли Амины и их свойства
Амины и их свойства Кислородсодержащие соединения серы
Кислородсодержащие соединения серы Арены. Бензол. Урок химии. 10 класс
Арены. Бензол. Урок химии. 10 класс Газы, применяемые при сварке
Газы, применяемые при сварке Методы разделения и очистки биомолекул. Ультрацентрифугирование (седиментация)
Методы разделения и очистки биомолекул. Ультрацентрифугирование (седиментация) Алкадиены (диены, диеновые углеводороды)
Алкадиены (диены, диеновые углеводороды) Алюминий. Сплавы алюминия
Алюминий. Сплавы алюминия Полисахариды. Крахмал и целлюлоза
Полисахариды. Крахмал и целлюлоза Коррозия металлов
Коррозия металлов История мыловарения
История мыловарения Твердые вещества
Твердые вещества Влияние циклогексанола селективную очистку масляных фракций нефти N-метилпирролидоном (N-МП)
Влияние циклогексанола селективную очистку масляных фракций нефти N-метилпирролидоном (N-МП) Мыловарение. Мыло своими руками
Мыловарение. Мыло своими руками Азотная кислота и нитраты
Азотная кислота и нитраты Химические свойства оксидов
Химические свойства оксидов Сулы ерітінділер. Тірі ағза сұйықтықтарындағы тепе-теңдік
Сулы ерітінділер. Тірі ағза сұйықтықтарындағы тепе-теңдік Жиры и масла
Жиры и масла Алкадиены. Строение алкадиенов
Алкадиены. Строение алкадиенов Експлуатаційні матеріали. Пальне для карбюраторних, дизельних двигунів. Змащувальні масла, пластичні мастила військової техніки
Експлуатаційні матеріали. Пальне для карбюраторних, дизельних двигунів. Змащувальні масла, пластичні мастила військової техніки Prezentatsia
Prezentatsia Геохимия стабильных изотопов, Радиоуглеродный метод
Геохимия стабильных изотопов, Радиоуглеродный метод Решение задач в химии (ОГЭ, ЕГЭ, Олимпиады)
Решение задач в химии (ОГЭ, ЕГЭ, Олимпиады) История открытия фосфора
История открытия фосфора