- Обмен липидов

Содержание
- 2. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ Источник липидов: жирная растительная и животная пища. Потребность взрослого человека в жире
- 3. Полость рта – нет ферментов для переваривания липидов. Роль язычной липазы в полости рта. Желудок –
- 4. СХЕМА расщепления жиров в процессе пищеварения
- 5. Этапы переваривания жиров в тонком кишечнике 1 этап – эмульгирование жира жёлчью Жёлчь имеет щелочную реакцию.
- 6. Соли жёлчных кислот и фосфолипиды жёлчи способствуют образованию устойчивых эмульсий (3, 7, 12). Жёлчные кислоты образуются
- 7. Жёлчные кислоты конъюгируют с аминокислотами: глицином или таурином. Из холевой образуются гликохолевая и таурохолевая кислоты (в
- 8. Механизм действия жёлчных кислот Резко снижают поверхностное натяжение на поверхности раздела фаз жир/вода, капли жира дробятся,
- 9. Функции солей жёлчных кислот : как поверхностно активные вещества, они эмульгируют частицы жира, 2) активируют панкреатическую
- 10. липаза Панкреатическая липаза, оптимум рН=8,0-9,0. В кишечник поступает в виде пролипазы и превращается в активную форму
- 11. 2 этап - гидролиз липазами поджелудочной железы, слизистой оболочки тонкого кишечника 1) Триглицерины: активированная липаза гидролизует
- 12. 2) Фосфолипиды: гидролиз полностью или частично под действием фосфолипаз ФЛ поступают в тонкий кишечник (~1-2 г/сутки
- 13. 3) Стероиды в кишечнике гидролизует холестераза поджелудочной железы. Эфиры стероидов распадаются на жирные кислоты и холестерин.
- 14. Состав мицелл: жёлчные кислоты и их соли (основной компонент), фосфолипиды, холестерол, жирорастворимые вита-мины образуют оболочки мицелл,
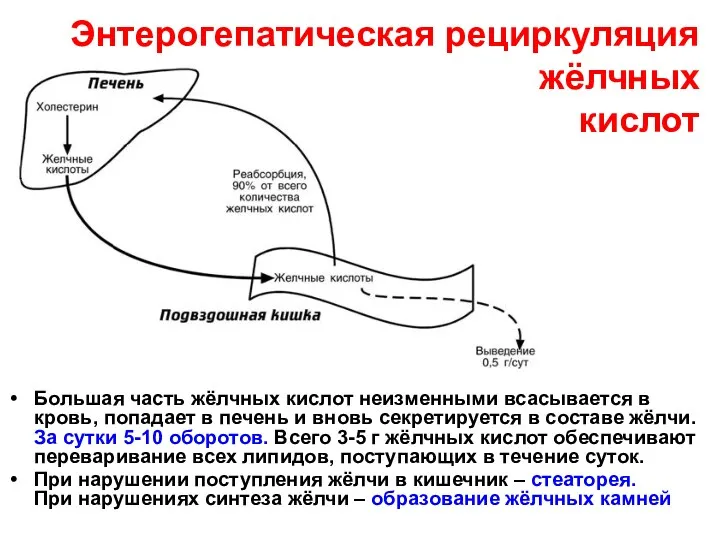
- 15. Энтерогепатическая рециркуляция жёлчных кислот Большая часть жёлчных кислот неизменными всасывается в кровь, попадает в печень и
- 16. 3 этап – всасывание продуктов гидролиза В клетках кишечной стенки – ресинтез новых триглицеринов (нейтральный жир)
- 17. 1) 2-моноацилглицеридный путь: участвуют экзогенные 2-МАГ и жирные кислоты (ацил-S-КоА). В гладком ЭПР мульти- ферментный комплекс
- 18. 2 путь глицеролфосфатный активируется в шероховатом ЭПР при отсутствии 2-МАГ и наличии большого количества жирных кислот.
- 19. Ресинтез других липидов в энтероцитах Реэтерификация холестерола Холестерол этерифицируется ферментом АХАТ с использованием ацил~SКоА. Реэтерификация напрямую
- 20. Жиры нерастворимы в воде и не могут в свободном состоянии находиться в водной среде крови, поскольку
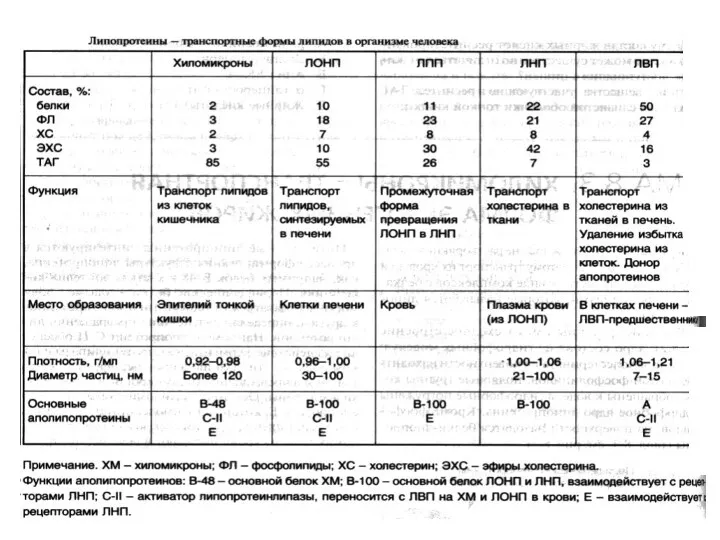
- 21. КЛАССИФИКАЦИЯ ЛИПОПРОТЕИНОВ КРОВИ Липопротеины называют по плотности или (по положению, которое они занимают на электрофореграмме): ХМ
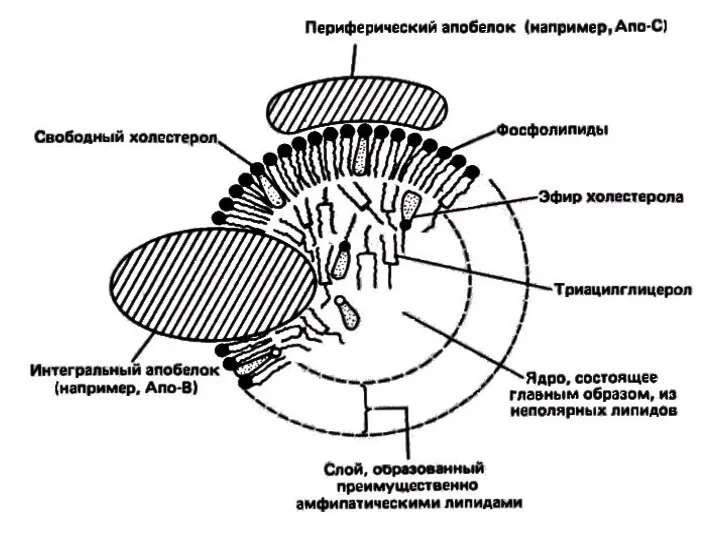
- 22. Состав липопротеинов: белки (апопротеины), фосфолипиды, нейтральные жиры, эфиры холестерола, свободный холестерол. Снаружи - вещества с полярными
- 23. Белки липопротеинов Белки липопротеинов называют апобелками, выделяют несколько их типов – А, В, С, D, Е.
- 27. ХИЛОМИКРОНЫ В энтероцитах ресинтезированные ТАГ, ФЛ, эфиры ХС и поступивший ХС соединяются с апобелками и образуют
- 28. Если сразу после еды взять кровь, то сыворотка будет мутная, на вид белёсая. Ее называют хилёзной
- 29. Липопротеинлипаза может убрать до 90% ТАГ, находящихся в ХМ (или ЛПОНП)
- 30. ЛП-ЛИПАЗА расщепляет ТАГ в ХМ с образованием тех же продуктов: - глицерол высшие жирные кислоты Обычно
- 31. ХМ после ухода ТАГ становятся хилоремнантами ("ремнанты" - остатки). Остаточные ХМ поглощаются клетками печени путём рецептор-опосредованного
- 32. ЛИПОПРОТЕИНЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ ЛПОНП образуются в печени Состав. 10% белок, 90% липиды: 55% - ТАГ
- 33. ЛПОНП разносятся из печени в другие ткани с током крови Катаболизм ЛПОНП идёт по такому же
- 35. ЛИПОПРОТЕИНЫ НИЗКОЙ ПЛОТНОСТИ ЛПНП образуются из остатков ЛПОНП: бόльшая часть в печени, часть непосредственно в плазме
- 36. Распад ЛПНП до ремнантов: - в печени около 50%, - во внепеченочных тканях ещё 50% Разрушение
- 37. ЛИПОПРОТЕИНЫ ВЫСОКОЙ ПЛОТНОСТИ ЛПВП – синтезируются в печени, но окончательно формируются в крови. Состав: 50%белок, 25%ФЛ,
- 38. На поверхности ЛПВП расположены фосфолипиды – лецитины(фосфатидилхолин) Фермент ЛХАТ отнимает жирную кислоту от лецитина, переносит на
- 40. НАРУШЕНИЯ ОБМЕНА ЛИПОПРОТЕИНОВ КРОВИ ЛПОНП и ЛПНП считаются "атерогенными", так как они переносят холестерол к тканям
- 41. ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ включает: 1) гидролиз ТАГ – липолиз 2) β-окисление ЖК, распад глицерина до конечных
- 43. 1) Липолиз Мобилизация ТАГ из депо: гидролиз липидов до СЖК и глицерина, которые используются как источник
- 44. 2а) β-окисление жирных кислот Впервые механизм изучен в 1904 г. Кнопом ЖК окисляется по β-атому С:
- 46. 1 этап: активация ЖК. Фермент ацил-КоА-синтетаза (АТФ) RCOOH + HS-КоА+АТФ → RCO~SКоА+АМФ+ФФ+H2O
- 47. 2 этап – сам процесс β-окисления R-CH2-CH2-CO~SКоА R-CH=CH-CO~SКоА R-CHОН-CH2-CO~SКоА R-CО-CH2-CO~SКоА R-CО~SКоА + CH3-CO~SКоА Если пальмитиновая кислота:
- 48. энергетическое значение β-окисления у пальмитиновой кислоты 16С 7 циклов окисления (ФАДН2 + НАДН = 5 АТФ
- 49. Окисление ненасыщенных жирных кислот При окислении ненасыщенных жирных кислот возникает потребность клетки в дополнительных ферментах изомеразах:
- 50. Окисление жирных кислот с нечетным числом атомов углерода Жирные кислоты с нечетным числом углеродов поступают в
- 51. 2б) РАСПАД ГЛИЦЕРОЛА до СО2 и Н2О Ферменты глицеролкиназа (-1АТФ) глицерофосфатдегидрогеназа (НАД+→НАДН) (+3АТФ) триозофосфатизомераза далее окисление
- 52. Энергетическое значение распада глицерола до Н2О и СО2 Дегидрогеназа (НАД+→НАДН) 3 АТФ 3ФГА до Ас~КоА (2
- 53. Субстраты - предшественники для синтеза липидов ДОАФ (промежуточный продукт гликолиза) → глицерол, если образование липидов идёт
- 54. Витамин В3 (пантотен) в составе ацилпереносящего белка и коэнзима А
- 55. 3) Синтез жирных кислот При синтезе наращиваются 2С-фрагменты и затем восстанавливаются с участием НАДФН. Работает мультиферментный
- 56. 1 этап – образование малонил~КоА СН3СО~КоА + СО2 → СООН-СН2СО~КоА Фермент карбоксилаза (кофермент – биотин) использует
- 58. ацетил-трансфераза присоединяет СН3-СО-КоА к SH-гр. цистеина (SH-ЦИС-) малонил-трансфераза присоединяет СООН-СН2-СО-КоА по АПБ-SH-группе (обе эти трансферазы относят
- 59. 4) Синтез триацилглицеролов 2 пути синтеза ТАГ: 1) на основе глицерина – в кишечнике и почках,
- 60. Обмен углеводов и обмен ТАГ связаны односторонне: 1) из углеводов жиры синтезируются (АсКоА из глюкозы в
- 61. СИНТЕЗ и ДЕПОНИРОВАНИЕ ЖИРОВ
- 62. Транспорт жиров из печени В печени синтезируется большое количество различных видов эндогенного жира. Жир выводится из
- 63. 5) Синтез фосфолипидов Первый путь синтеза (схема далее на слайде) Активация и перенос азотистых оснований. Холин/или
- 64. Первый путь синтеза ФЛ (лёгкие, кишечник)
- 65. Второй путь синтеза ФЛ Фосфатидная кислота – ключевой продукт обмена липидов. До фосфатидной кислоты синтез как
- 66. Все азотистые основания фосфолипидов могут переходить друг в друга с участием липотропных веществ Пути синтеза фосфолипидов
- 68. При наличии 1) глицерол-3-фосфата и 2) ацил~S-КоА синтезируется фосфатидная кислота.
- 70. Липотропные вещества Липотропные вещества - сдвиг синтеза жиров в сторону фосфолипидов,ограничение синтеза и накопления нейтрального жира
- 71. Судьба фосфолипидов Построение клеточных мембран, асимметрия локализации в наружном и внутреннем слое плазматической мембраны (ФС снаружи
- 72. Распад фосфолипидов Вспомнить действие фосфолипаз при пищеварении (внешний обмен липидов). В организме распад фосфолипидов осуществляют тканевые
- 73. Фосфолипиды гидролизуются фосфо-липазами, наиболее важны ФЛА2(В) и ФЛС Продукты полного гидролиза: - глицерин, - высшие жирные
- 74. Метаболизм арахидоновой кислоты Производные арахидоновой кислоты – эйкозаноиды Образуется несколько классов веществ: простагландины, простациклины, тромбоксаны, лейкотриены
- 75. Схема синтеза эйкозаноидов
- 77. Основные производные арахидоновой кислоты
- 78. Эйкозаноиды Строение. В составе молекул содержатся: циклы (ПГ, ТХ и ПЦ, а у ЛТ их нет),
- 79. Биологическая активность эйкозаноидов Эйкозаноиды – локальные биорегуляторы, действуют путем связывания с мембранными рецепторами в непосредственной близости
- 80. ПРОСТАГЛАНДИНЫ Функции (используемые для терапии) : расширение сосудов, снижение артериального давления ингибиторы тромбообразования PgF2 прерывает беременность
- 81. 6) Синтез кетоновых тел происходит только в печени, клетки остальных тканей (кроме эритроцитов) лишь потребляют их
- 82. Утилизация кетоновых тел идёт с обходом необратимого этапа их синтеза: в трансферазной реакции к ацетоацетату переносится
- 83. Общая схема синтеза и утилизации кетоновых тел
- 84. 7) СИНТЕЗ ХОЛЕСТЕРОЛА Холестерол – стероид, характерный только для животных организмов. Содержание ХС в организме: 50%
- 85. Синтез ХС идёт в цитозоле клеток – один из самых длинных метаболических путей. Расшифровал последовательность пути
- 86. Образование мевалоновой кислоты
- 87. II этап – мевалоновая кислота конденсируется в сквален Сначала мевалоновая кислота теряет СО2 и превращается в
- 88. II этап синтеза холестерола Через стадию Геранилдифосфата (С10)
- 89. ГМГ-редуктаза ключевой фермент синтеза холестерола, его активность регулируется: высокие концентрации ХС блокируют синтез фермента в печени
- 90. Пути холестерола Встраивание в мембраны клеток На синтезы: -- желчных кислот -- витамина D -- эфиров
- 91. II этап синтеза холестерола промежуточные продукты синтеза - геранил-геранил и - фарнезил, а также - долихол,
- 92. Использование липидных якорей в ЭПР. Начальные этапы N-гликозилирования белков (связь через АСН) Формирование сложного олигосахарида, связанного
- 93. Липопротеины с С-концевой липидной группировкой. Пренилирование белков S-пренилирование белков – присоединение изопреноидов к атому серы в
- 94. Заякоривание белков в мембране
- 96. Основные виды патологии липидного обмена Ожирение - избыточное отложение жира чаще алиментарного, реже эндокринного характера 1)
- 97. Осложнения ожирения заболевание эндокринной системы → дает ослож- нения сразу на несколько систем организма Нарушения желудочно-кишечного
- 98. Основные виды патологии липидного обмена Желчекаменная болезнь Холестерол является компонентом желчных кислот и может находиться в
- 99. Камни желчных путей
- 100. Основные виды патологии липидного обмена Дислипопротеинемии – могут быть как наследственными, так и приобретёнными заболеваниями. Сопровождаются
- 101. Нарушения обмена разных ЛП можно приобрести при ряде заболеваний (сахарный диабет, алкоголизм, снижение функции щитовидной железы,
- 102. Типы дислипопротеинемий I тип: повышена концентрация ХМ При наследственной недостаточности ЛП-липазы ХМ не исчезают даже через
- 103. II тип: повышено содержание ЛПНП или суммарно ЛПОНП + ЛПНП Причина в недостатке или отсутствии рецепторов
- 104. IV тип. В крови повышено содержание ЛПОНП, ХС, ТАГ. Это характерно для сахарного диабета II типа,
- 105. Основные виды патологии липидного обмена Атеросклероз Причиной атеросклероза может быть наруше-ние как синтеза, так и транспорта
- 106. Синтез холестерола, экзогенный холестерол пищи и пути транспорта холестерола
- 107. Формирование атеросклероза При повреждении эндотелия ЛП проникают в сосудистую стенку, поглощаются фагоцитами. Все их составные части
- 108. • холестериновый коэффициент атерогенности k = ХС общ – ХС ЛПВП ХС – концентрация общего холестерола,
- 110. Все основные виды патологии липидного обмена так или иначе взаимосвязаны
- 112. Скачать презентацию

ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ
Источник липидов:
жирная растительная и животная пища.
Потребность
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ
Источник липидов:
жирная растительная и животная пища.
Потребность

Полость рта – нет ферментов для переваривания липидов. Роль язычной липазы
Полость рта – нет ферментов для переваривания липидов. Роль язычной липазы
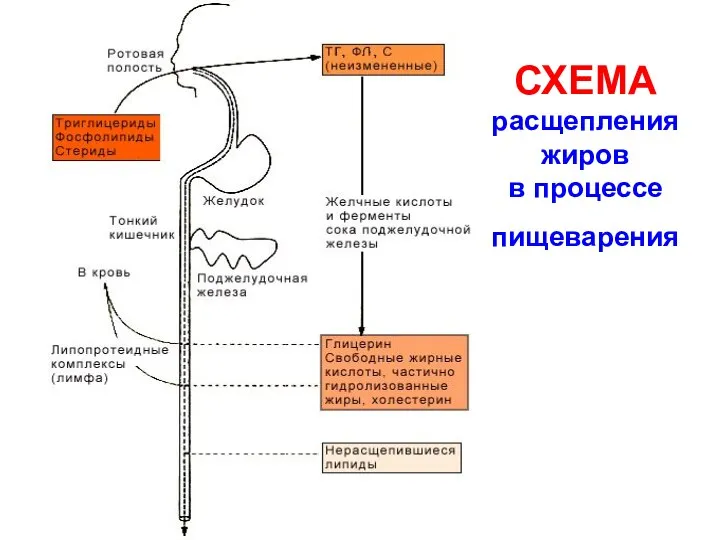
СХЕМА расщепления жиров
в процессе пищеварения
СХЕМА расщепления жиров
в процессе пищеварения

Этапы переваривания жиров
в тонком кишечнике
1 этап – эмульгирование жира жёлчью
Жёлчь
Этапы переваривания жиров
в тонком кишечнике
1 этап – эмульгирование жира жёлчью
Жёлчь
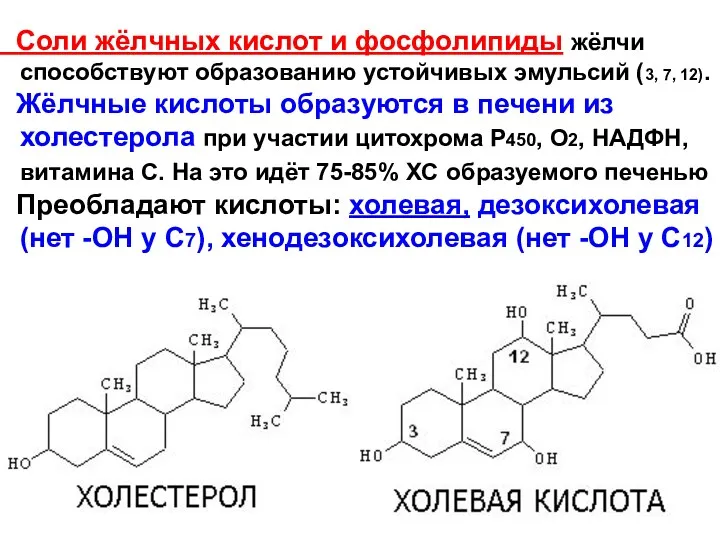
Соли жёлчных кислот и фосфолипиды жёлчи способствуют образованию устойчивых эмульсий
Соли жёлчных кислот и фосфолипиды жёлчи способствуют образованию устойчивых эмульсий
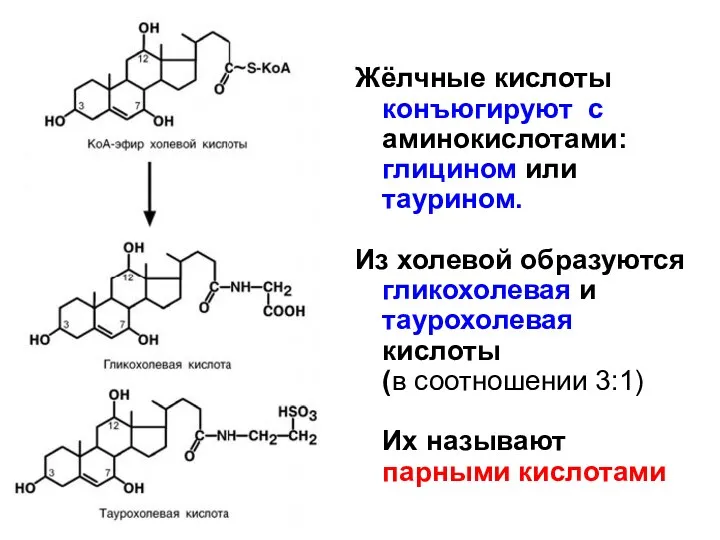
Жёлчные кислоты конъюгируют с аминокислотами: глицином или таурином.
Из холевой образуются
Жёлчные кислоты конъюгируют с аминокислотами: глицином или таурином.
Из холевой образуются

Механизм действия жёлчных кислот
Резко снижают поверхностное натяжение
на поверхности раздела фаз
Механизм действия жёлчных кислот
Резко снижают поверхностное натяжение
на поверхности раздела фаз

Функции солей жёлчных кислот :
как поверхностно активные вещества, они
Функции солей жёлчных кислот :
как поверхностно активные вещества, они

липаза
Панкреатическая липаза, оптимум рН=8,0-9,0. В кишечник поступает в виде пролипазы и
липаза
Панкреатическая липаза, оптимум рН=8,0-9,0. В кишечник поступает в виде пролипазы и

2 этап - гидролиз липазами поджелудочной железы, слизистой оболочки тонкого кишечника
2 этап - гидролиз липазами поджелудочной железы, слизистой оболочки тонкого кишечника
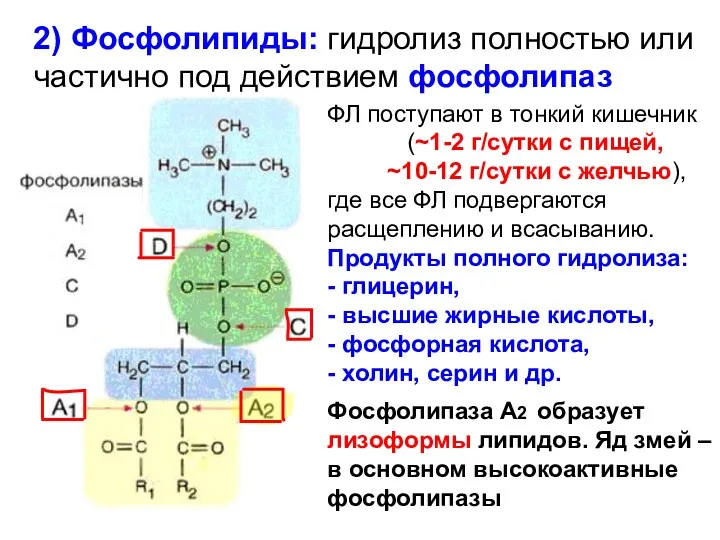
2) Фосфолипиды: гидролиз полностью или частично под действием фосфолипаз
ФЛ
2) Фосфолипиды: гидролиз полностью или частично под действием фосфолипаз
ФЛ

3) Стероиды в кишечнике гидролизует холестераза поджелудочной железы. Эфиры стероидов распадаются
3) Стероиды в кишечнике гидролизует холестераза поджелудочной железы. Эфиры стероидов распадаются

Состав мицелл:
жёлчные кислоты и их соли (основной компонент), фосфолипиды,
Состав мицелл: жёлчные кислоты и их соли (основной компонент), фосфолипиды,
Энтерогепатическая рециркуляция жёлчных
кислот
Большая часть жёлчных кислот неизменными всасывается в кровь,
Энтерогепатическая рециркуляция жёлчных
кислот
Большая часть жёлчных кислот неизменными всасывается в кровь,

3 этап – всасывание продуктов гидролиза
В клетках кишечной стенки – ресинтез
3 этап – всасывание продуктов гидролиза
В клетках кишечной стенки – ресинтез
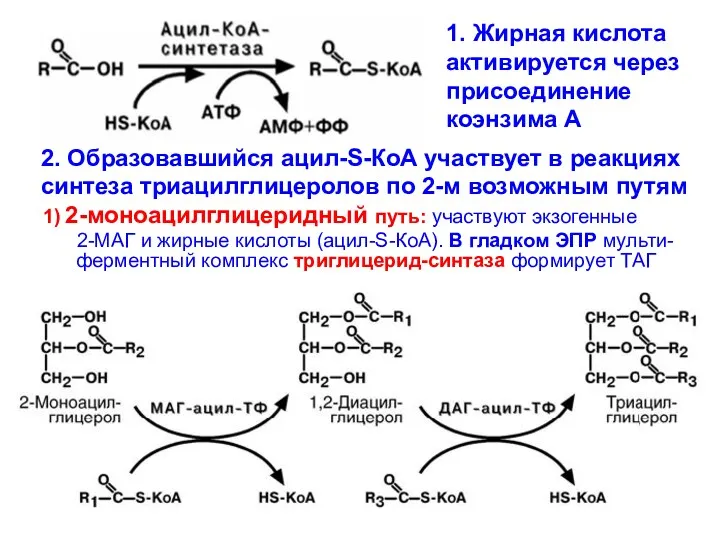
1) 2-моноацилглицеридный путь: участвуют экзогенные
2-МАГ и жирные кислоты
1) 2-моноацилглицеридный путь: участвуют экзогенные
2-МАГ и жирные кислоты
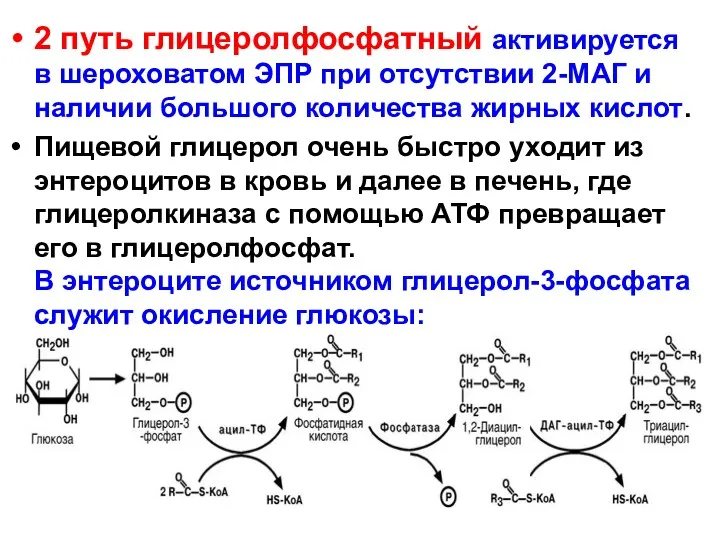
2 путь глицеролфосфатный активируется
в шероховатом ЭПР при отсутствии 2-МАГ и
2 путь глицеролфосфатный активируется в шероховатом ЭПР при отсутствии 2-МАГ и

Ресинтез других липидов
в энтероцитах
Реэтерификация холестерола Холестерол этерифицируется ферментом АХАТ с
Ресинтез других липидов
в энтероцитах
Реэтерификация холестерола Холестерол этерифицируется ферментом АХАТ с

Жиры нерастворимы в воде и не могут
в свободном состоянии находиться
Жиры нерастворимы в воде и не могут в свободном состоянии находиться

КЛАССИФИКАЦИЯ
ЛИПОПРОТЕИНОВ КРОВИ
Липопротеины называют по плотности или (по положению, которое они
КЛАССИФИКАЦИЯ
ЛИПОПРОТЕИНОВ КРОВИ
Липопротеины называют по плотности или (по положению, которое они

Состав липопротеинов:
белки (апопротеины), фосфолипиды,
нейтральные жиры, эфиры холестерола,
свободный
Состав липопротеинов: белки (апопротеины), фосфолипиды, нейтральные жиры, эфиры холестерола, свободный

Белки липопротеинов
Белки липопротеинов называют апобелками, выделяют несколько их типов – А,
Белки липопротеинов
Белки липопротеинов называют апобелками, выделяют несколько их типов – А,
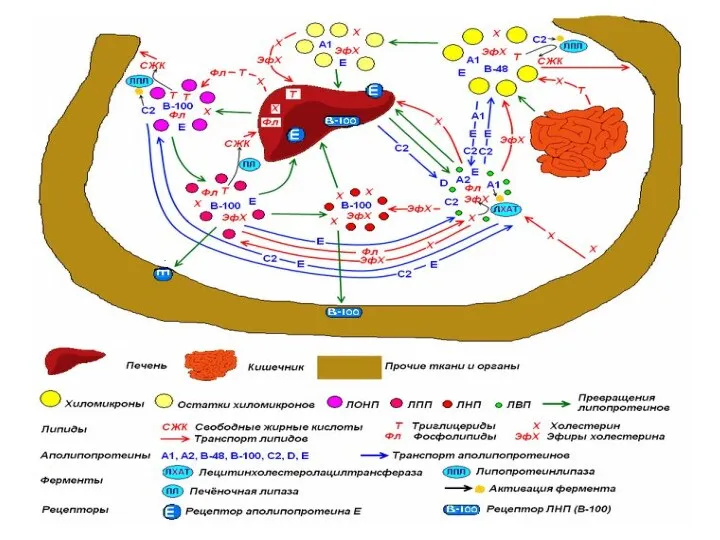

ХИЛОМИКРОНЫ
В энтероцитах ресинтезированные ТАГ, ФЛ, эфиры ХС и поступивший ХС
ХИЛОМИКРОНЫ
В энтероцитах ресинтезированные ТАГ, ФЛ, эфиры ХС и поступивший ХС

Если сразу после еды взять кровь,
то сыворотка будет мутная, на
Если сразу после еды взять кровь, то сыворотка будет мутная, на
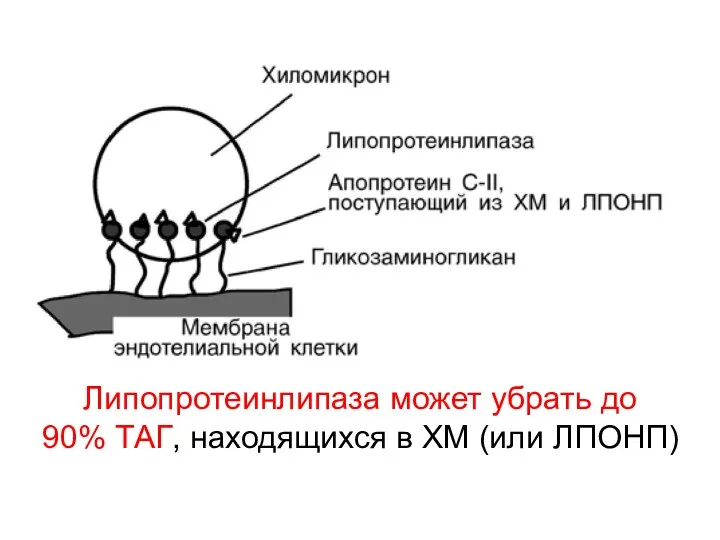
Липопротеинлипаза может убрать до
90% ТАГ, находящихся в ХМ (или ЛПОНП)
Липопротеинлипаза может убрать до
90% ТАГ, находящихся в ХМ (или ЛПОНП)

ЛП-ЛИПАЗА расщепляет ТАГ в ХМ
с образованием тех же продуктов:
-
ЛП-ЛИПАЗА расщепляет ТАГ в ХМ с образованием тех же продуктов: -

ХМ после ухода ТАГ становятся хилоремнантами ("ремнанты" - остатки).
Остаточные
ХМ после ухода ТАГ становятся хилоремнантами ("ремнанты" - остатки). Остаточные

ЛИПОПРОТЕИНЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ
ЛПОНП образуются в печени
Состав. 10% белок, 90% липиды:
ЛИПОПРОТЕИНЫ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ
ЛПОНП образуются в печени
Состав. 10% белок, 90% липиды:

ЛПОНП разносятся из печени
в другие ткани с током крови
Катаболизм
ЛПОНП разносятся из печени
в другие ткани с током крови
Катаболизм
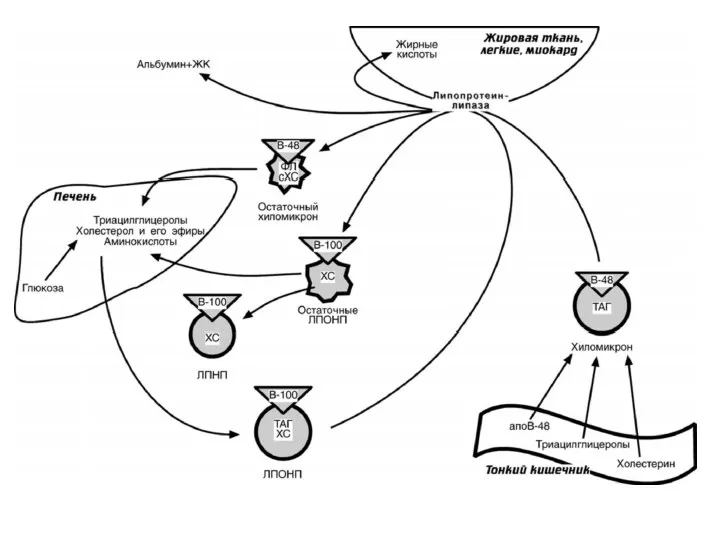

ЛИПОПРОТЕИНЫ НИЗКОЙ ПЛОТНОСТИ
ЛПНП образуются из остатков ЛПОНП:
бόльшая часть в печени,
ЛИПОПРОТЕИНЫ НИЗКОЙ ПЛОТНОСТИ
ЛПНП образуются из остатков ЛПОНП:
бόльшая часть в печени,

Распад ЛПНП до ремнантов:
- в печени около 50%,
- во
Распад ЛПНП до ремнантов: - в печени около 50%, - во

ЛИПОПРОТЕИНЫ ВЫСОКОЙ ПЛОТНОСТИ
ЛПВП – синтезируются в печени, но окончательно формируются в
ЛИПОПРОТЕИНЫ ВЫСОКОЙ ПЛОТНОСТИ
ЛПВП – синтезируются в печени, но окончательно формируются в

На поверхности ЛПВП расположены фосфолипиды – лецитины(фосфатидилхолин)
Фермент ЛХАТ отнимает
На поверхности ЛПВП расположены фосфолипиды – лецитины(фосфатидилхолин)
Фермент ЛХАТ отнимает
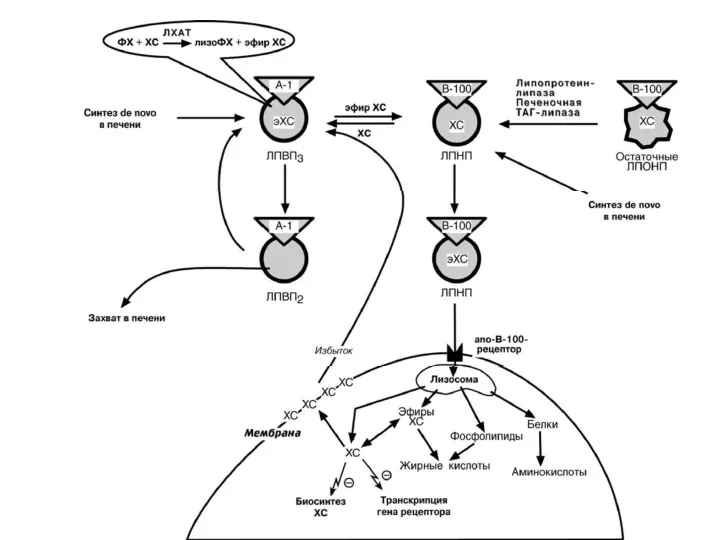

НАРУШЕНИЯ ОБМЕНА ЛИПОПРОТЕИНОВ КРОВИ
ЛПОНП и ЛПНП считаются "атерогенными", так как они
НАРУШЕНИЯ ОБМЕНА ЛИПОПРОТЕИНОВ КРОВИ
ЛПОНП и ЛПНП считаются "атерогенными", так как они

ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ включает:
1) гидролиз ТАГ – липолиз
2)
ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ включает:
1) гидролиз ТАГ – липолиз
2)

1) Липолиз
Мобилизация ТАГ из депо: гидролиз липидов до СЖК
и
1) Липолиз
Мобилизация ТАГ из депо: гидролиз липидов до СЖК и
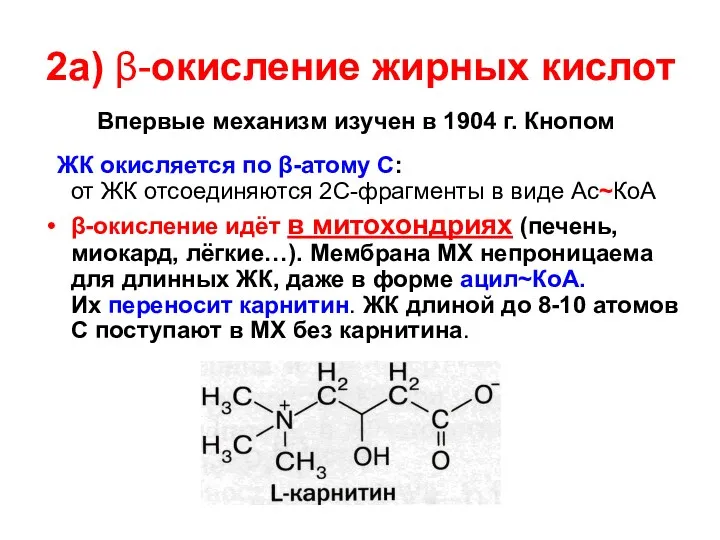
2а) β-окисление жирных кислот
Впервые механизм изучен в 1904 г.
2а) β-окисление жирных кислот
Впервые механизм изучен в 1904 г.
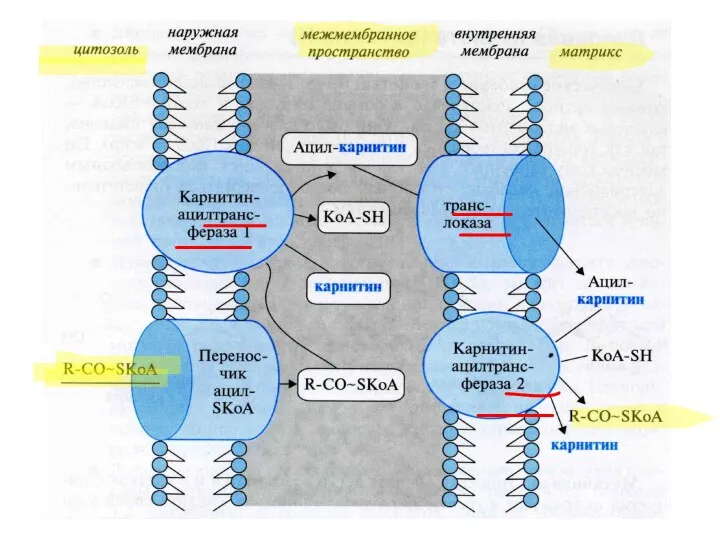
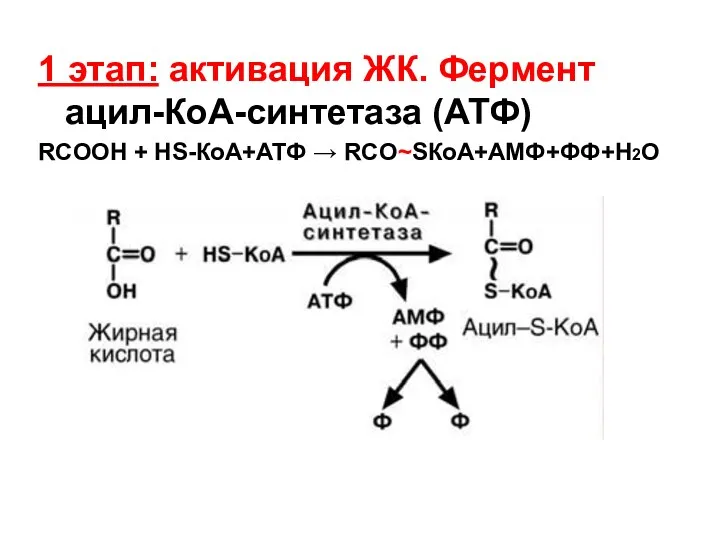
1 этап: активация ЖК. Фермент ацил-КоА-синтетаза (АТФ)
RCOOH + HS-КоА+АТФ → RCO~SКоА+АМФ+ФФ+H2O
1 этап: активация ЖК. Фермент ацил-КоА-синтетаза (АТФ)
RCOOH + HS-КоА+АТФ → RCO~SКоА+АМФ+ФФ+H2O

2 этап – сам процесс β-окисления
R-CH2-CH2-CO~SКоА
R-CH=CH-CO~SКоА
R-CHОН-CH2-CO~SКоА
R-CО-CH2-CO~SКоА
2 этап – сам процесс β-окисления
R-CH2-CH2-CO~SКоА
R-CH=CH-CO~SКоА
R-CHОН-CH2-CO~SКоА
R-CО-CH2-CO~SКоА

энергетическое значение
β-окисления
у пальмитиновой кислоты 16С 7 циклов окисления (ФАДН2 + НАДН
энергетическое значение
β-окисления
у пальмитиновой кислоты 16С 7 циклов окисления (ФАДН2 + НАДН

Окисление ненасыщенных жирных кислот
При окислении ненасыщенных жирных кислот возникает потребность
Окисление ненасыщенных жирных кислот
При окислении ненасыщенных жирных кислот возникает потребность

Окисление жирных кислот с нечетным числом атомов углерода
Жирные кислоты с
Окисление жирных кислот с нечетным числом атомов углерода
Жирные кислоты с
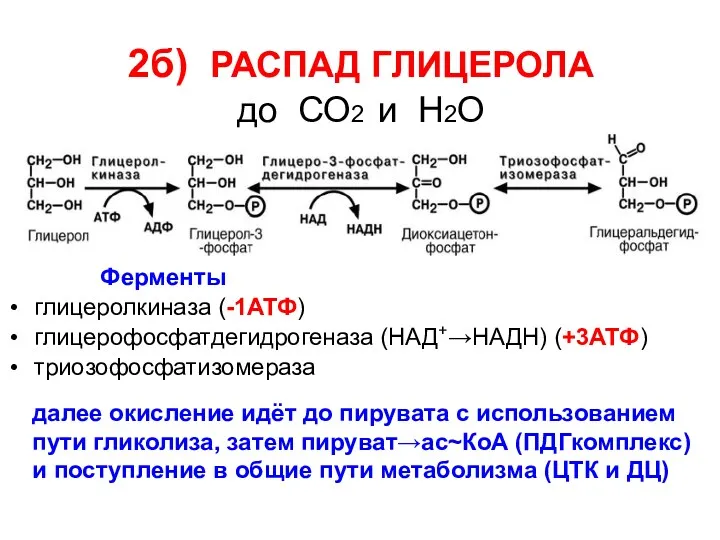
2б) РАСПАД ГЛИЦЕРОЛА
до СО2 и Н2О
Ферменты
глицеролкиназа (-1АТФ)
глицерофосфатдегидрогеназа (НАД+→НАДН) (+3АТФ)
триозофосфатизомераза
далее
2б) РАСПАД ГЛИЦЕРОЛА
до СО2 и Н2О
Ферменты
глицеролкиназа (-1АТФ)
глицерофосфатдегидрогеназа (НАД+→НАДН) (+3АТФ)
триозофосфатизомераза
далее

Энергетическое значение распада глицерола до Н2О и СО2
Дегидрогеназа (НАД+→НАДН) 3
Энергетическое значение распада глицерола до Н2О и СО2
Дегидрогеназа (НАД+→НАДН) 3

Субстраты - предшественники
для синтеза липидов
ДОАФ (промежуточный продукт гликолиза) → глицерол,
Субстраты - предшественники
для синтеза липидов
ДОАФ (промежуточный продукт гликолиза) → глицерол,
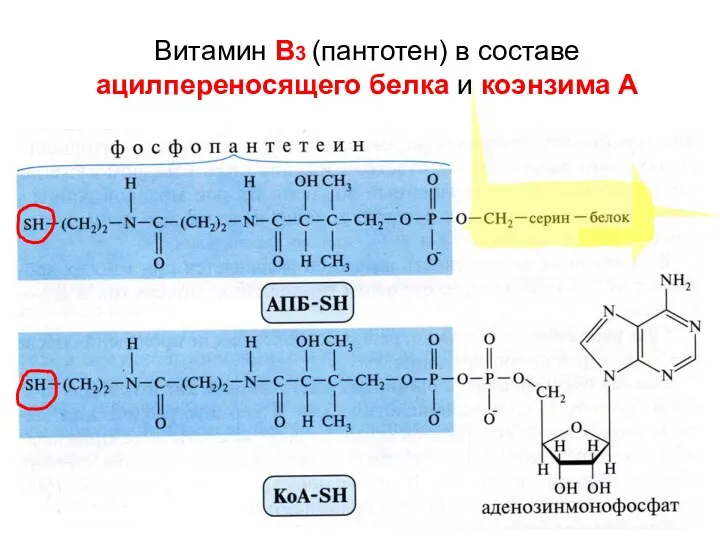
Витамин В3 (пантотен) в составе
ацилпереносящего белка и коэнзима А
Витамин В3 (пантотен) в составе
ацилпереносящего белка и коэнзима А

3) Синтез жирных кислот
При синтезе наращиваются 2С-фрагменты
и затем восстанавливаются
3) Синтез жирных кислот
При синтезе наращиваются 2С-фрагменты и затем восстанавливаются

1 этап – образование малонил~КоА
СН3СО~КоА + СО2 → СООН-СН2СО~КоА
Фермент карбоксилаза (кофермент
1 этап – образование малонил~КоА
СН3СО~КоА + СО2 → СООН-СН2СО~КоА
Фермент карбоксилаза (кофермент
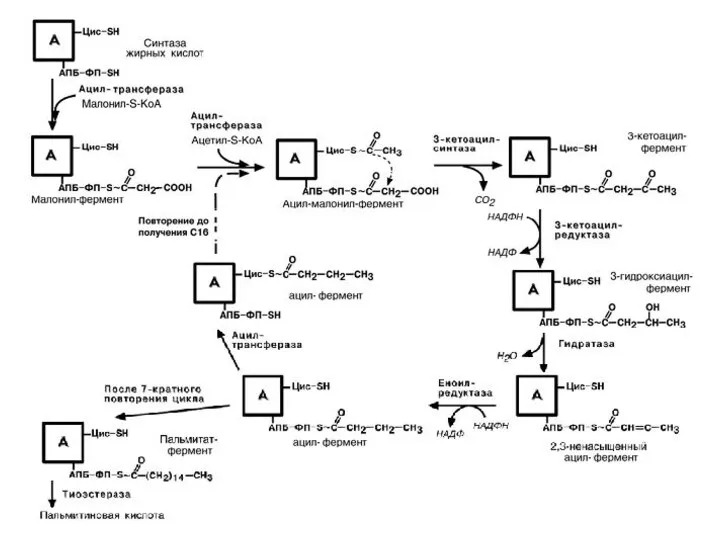

ацетил-трансфераза
присоединяет СН3-СО-КоА к SH-гр. цистеина (SH-ЦИС-)
малонил-трансфераза
присоединяет СООН-СН2-СО-КоА по АПБ-SH-группе
(обе
ацетил-трансфераза
присоединяет СН3-СО-КоА к SH-гр. цистеина (SH-ЦИС-)
малонил-трансфераза
присоединяет СООН-СН2-СО-КоА по АПБ-SH-группе
(обе

4) Синтез триацилглицеролов
2 пути синтеза ТАГ:
1) на основе глицерина
4) Синтез триацилглицеролов
2 пути синтеза ТАГ: 1) на основе глицерина
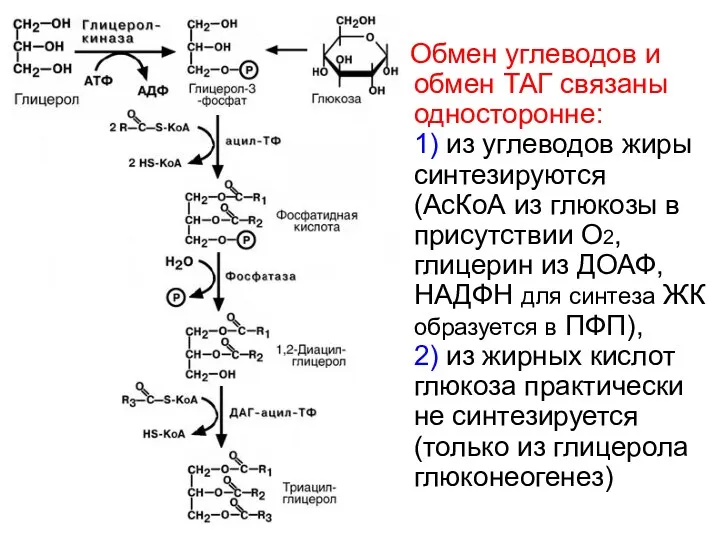
Обмен углеводов и обмен ТАГ связаны односторонне:
1) из углеводов
Обмен углеводов и обмен ТАГ связаны односторонне: 1) из углеводов
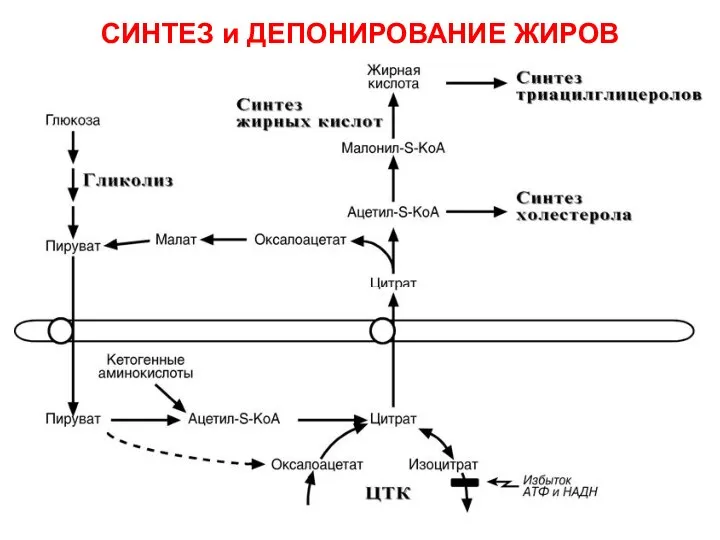
СИНТЕЗ и ДЕПОНИРОВАНИЕ ЖИРОВ
СИНТЕЗ и ДЕПОНИРОВАНИЕ ЖИРОВ

Транспорт жиров из печени
В печени синтезируется большое количество различных видов эндогенного
Транспорт жиров из печени
В печени синтезируется большое количество различных видов эндогенного

5) Синтез фосфолипидов
Первый путь синтеза (схема далее на слайде)
Активация и
5) Синтез фосфолипидов
Первый путь синтеза (схема далее на слайде)
Активация и
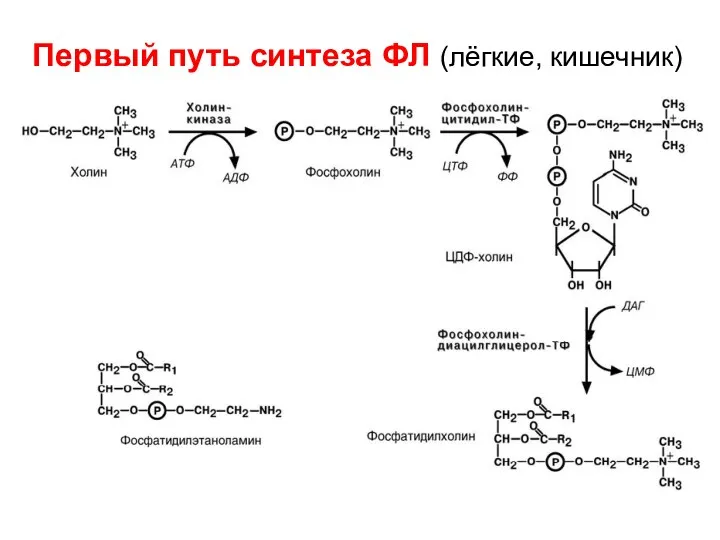
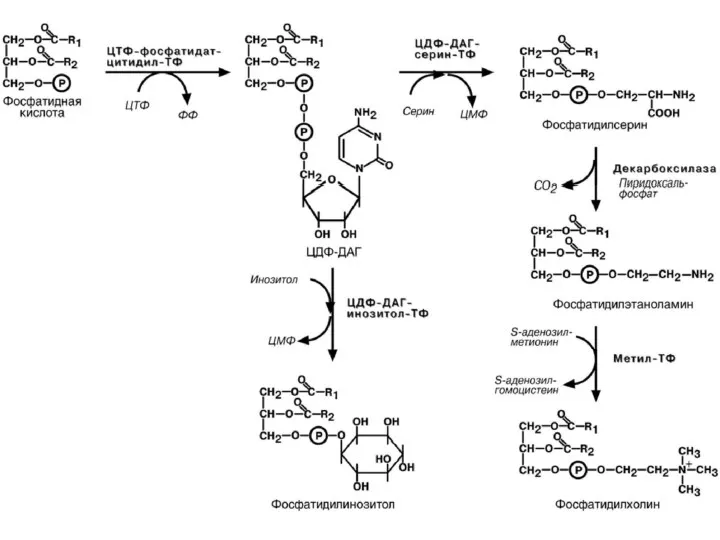
Первый путь синтеза ФЛ (лёгкие, кишечник)
Первый путь синтеза ФЛ (лёгкие, кишечник)

Второй путь синтеза ФЛ
Фосфатидная кислота – ключевой продукт обмена липидов. До
Второй путь синтеза ФЛ
Фосфатидная кислота – ключевой продукт обмена липидов. До
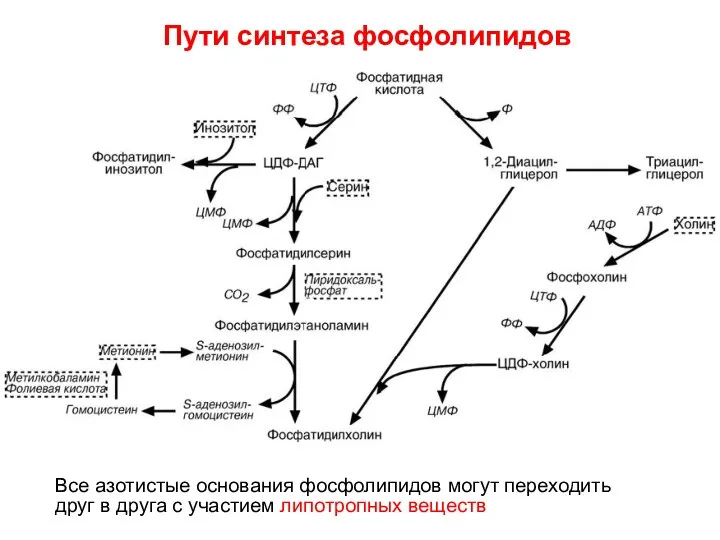
Все азотистые основания фосфолипидов могут переходить
друг в друга с участием
Все азотистые основания фосфолипидов могут переходить друг в друга с участием
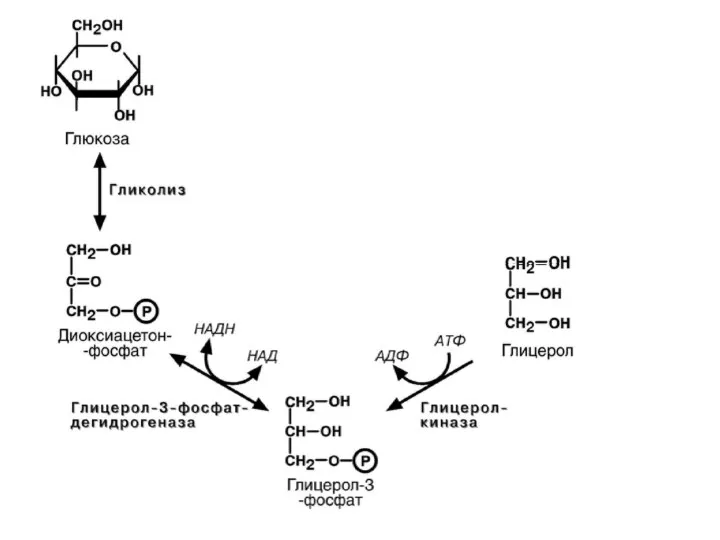
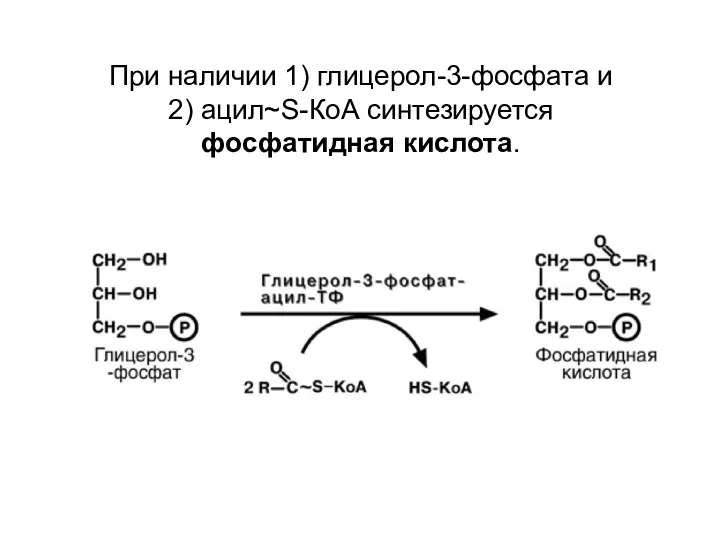
При наличии 1) глицерол-3-фосфата и
2) ацил~S-КоА синтезируется
фосфатидная кислота.
При наличии 1) глицерол-3-фосфата и
2) ацил~S-КоА синтезируется
фосфатидная кислота.

Липотропные вещества
Липотропные вещества - сдвиг синтеза жиров
в сторону фосфолипидов,ограничение синтеза
Липотропные вещества
Липотропные вещества - сдвиг синтеза жиров в сторону фосфолипидов,ограничение синтеза

Судьба фосфолипидов
Построение клеточных мембран, асимметрия локализации в наружном и внутреннем слое
Судьба фосфолипидов
Построение клеточных мембран, асимметрия локализации в наружном и внутреннем слое

Распад фосфолипидов
Вспомнить действие фосфолипаз при пищеварении (внешний обмен липидов).
В организме
Распад фосфолипидов
Вспомнить действие фосфолипаз при пищеварении (внешний обмен липидов).
В организме
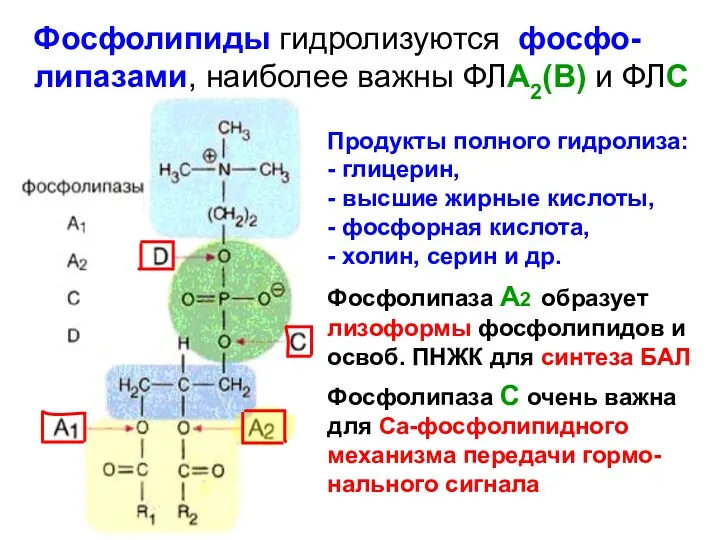
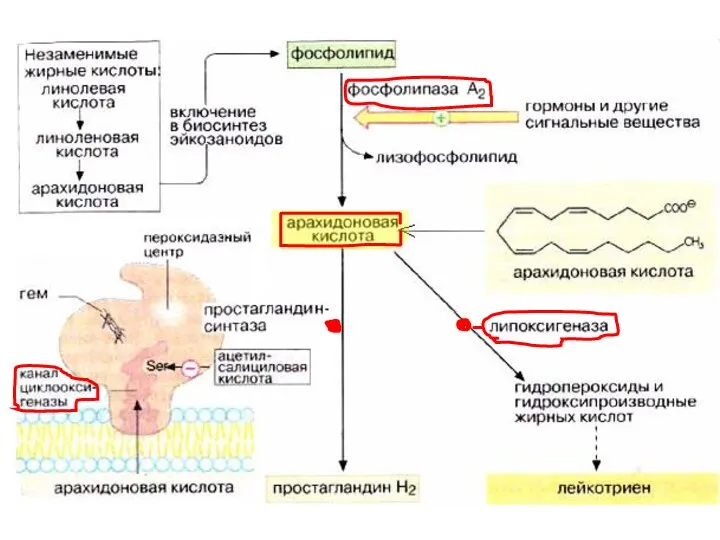
Фосфолипиды гидролизуются фосфо-липазами, наиболее важны ФЛА2(В) и ФЛС
Продукты полного
Фосфолипиды гидролизуются фосфо-липазами, наиболее важны ФЛА2(В) и ФЛС
Продукты полного

Метаболизм арахидоновой кислоты
Производные арахидоновой кислоты – эйкозаноиды
Образуется несколько классов
Метаболизм арахидоновой кислоты
Производные арахидоновой кислоты – эйкозаноиды
Образуется несколько классов
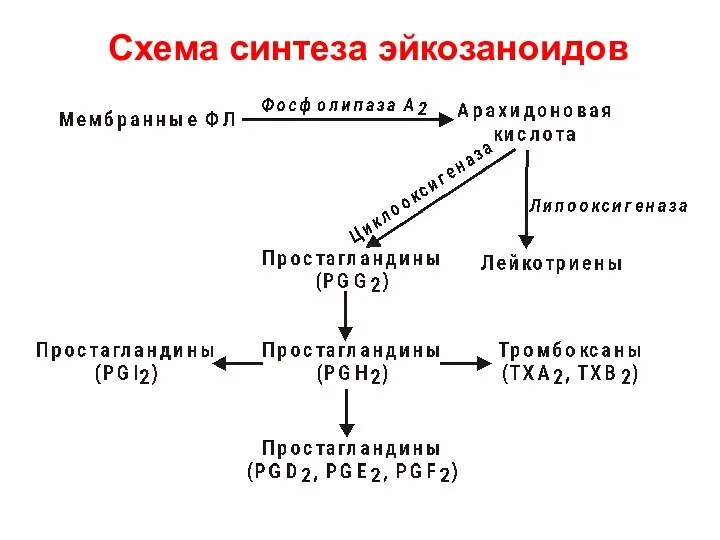
Схема синтеза эйкозаноидов
Схема синтеза эйкозаноидов
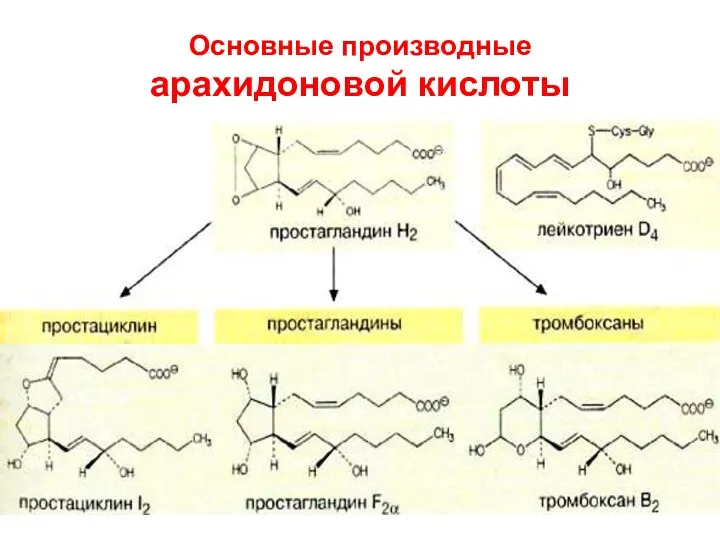
Основные производные
арахидоновой кислоты
Основные производные
арахидоновой кислоты

Эйкозаноиды
Строение. В составе молекул содержатся: циклы (ПГ, ТХ
Эйкозаноиды
Строение. В составе молекул содержатся: циклы (ПГ, ТХ

Биологическая активность эйкозаноидов
Эйкозаноиды – локальные биорегуляторы, действуют
путем связывания
Биологическая активность эйкозаноидов
Эйкозаноиды – локальные биорегуляторы, действуют путем связывания
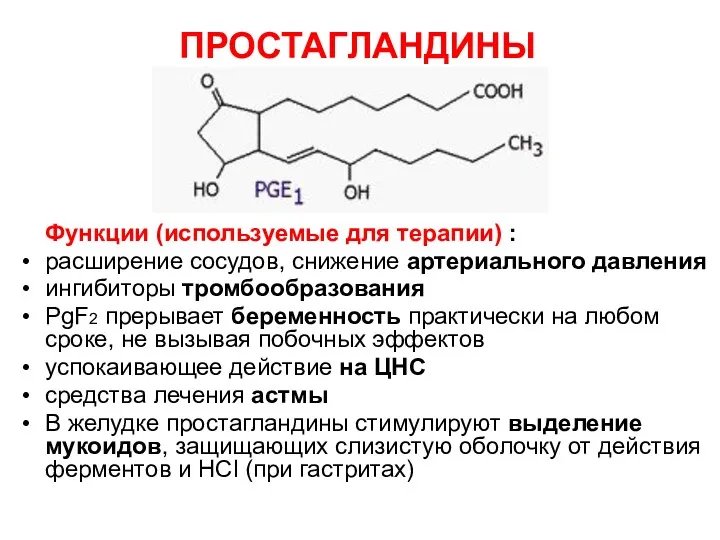
ПРОСТАГЛАНДИНЫ
Функции (используемые для терапии) :
расширение сосудов, снижение артериального давления
ингибиторы тромбообразования
PgF2
ПРОСТАГЛАНДИНЫ
Функции (используемые для терапии) :
расширение сосудов, снижение артериального давления
ингибиторы тромбообразования
PgF2
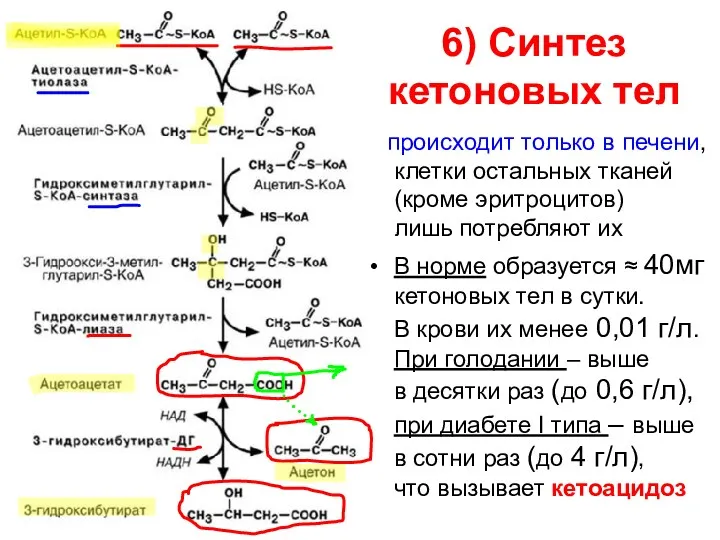
6) Синтез кетоновых тел
происходит только в печени, клетки остальных тканей
6) Синтез кетоновых тел
происходит только в печени, клетки остальных тканей
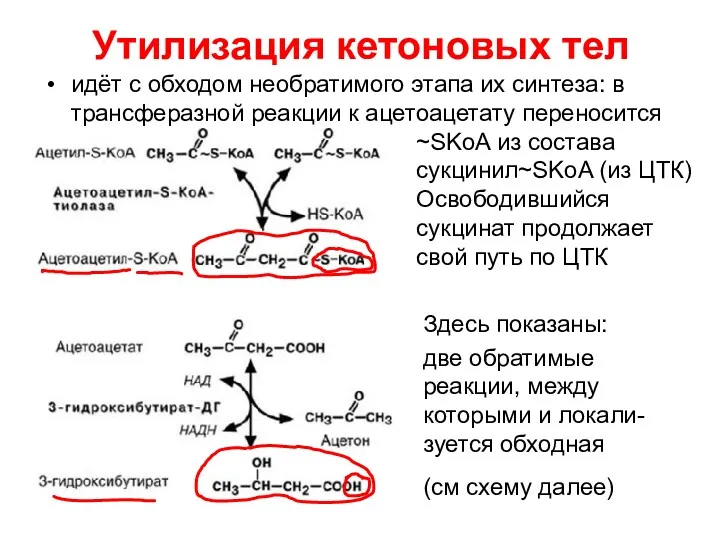
Утилизация кетоновых тел
идёт с обходом необратимого этапа их синтеза: в трансферазной
Утилизация кетоновых тел
идёт с обходом необратимого этапа их синтеза: в трансферазной
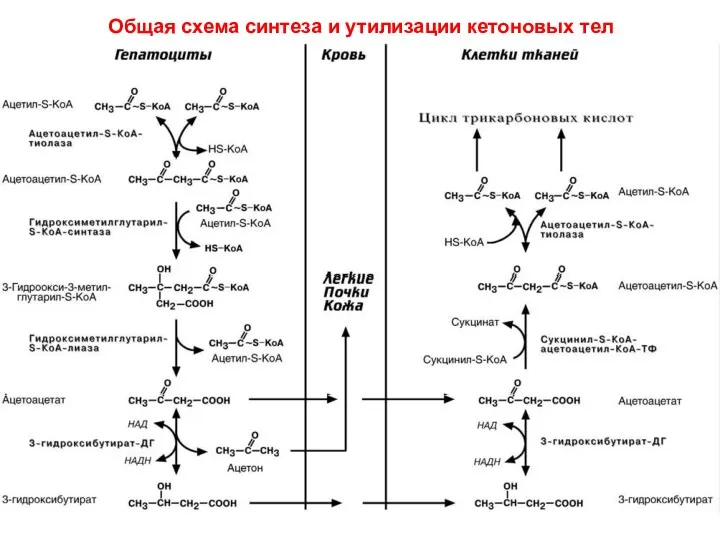
Общая схема синтеза и утилизации кетоновых тел
Общая схема синтеза и утилизации кетоновых тел

7) СИНТЕЗ ХОЛЕСТЕРОЛА
Холестерол – стероид, характерный только для животных организмов.
7) СИНТЕЗ ХОЛЕСТЕРОЛА
Холестерол – стероид, характерный только для животных организмов.

Синтез ХС идёт в цитозоле клеток –
один из самых длинных
Синтез ХС идёт в цитозоле клеток – один из самых длинных
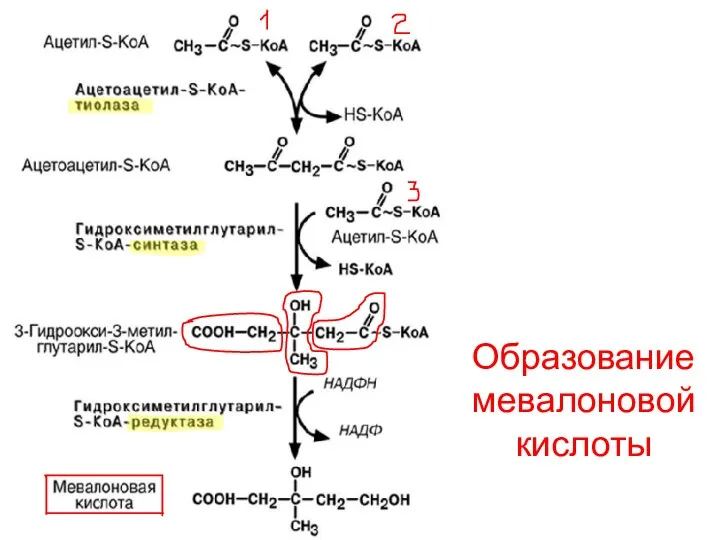
Образование мевалоновой кислоты
Образование мевалоновой кислоты

II этап – мевалоновая кислота
конденсируется в сквален
Сначала мевалоновая
II этап – мевалоновая кислота
конденсируется в сквален
Сначала мевалоновая
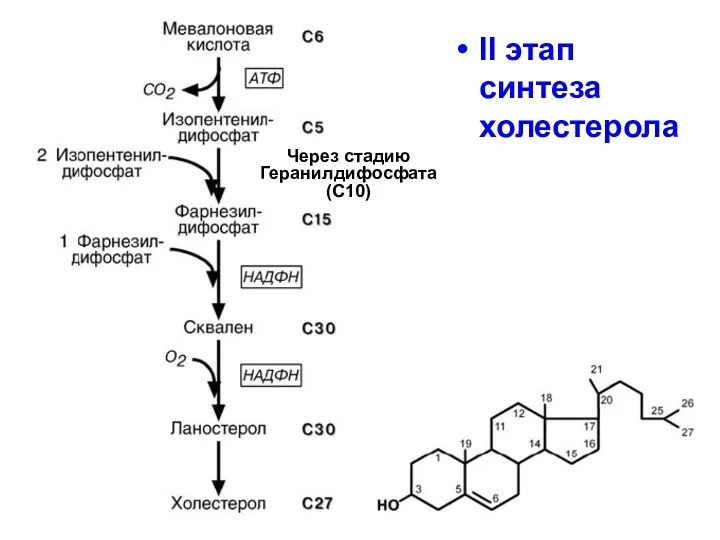
II этап синтеза холестерола
Через стадию Геранилдифосфата (С10)
II этап синтеза холестерола
Через стадию Геранилдифосфата (С10)

ГМГ-редуктаза ключевой фермент синтеза
холестерола, его активность регулируется:
высокие концентрации ХС
ГМГ-редуктаза ключевой фермент синтеза
холестерола, его активность регулируется:
высокие концентрации ХС

Пути холестерола
Встраивание в мембраны клеток
На синтезы:
-- желчных кислот
-- витамина D
-- эфиров
Пути холестерола
Встраивание в мембраны клеток
На синтезы:
-- желчных кислот
-- витамина D
-- эфиров
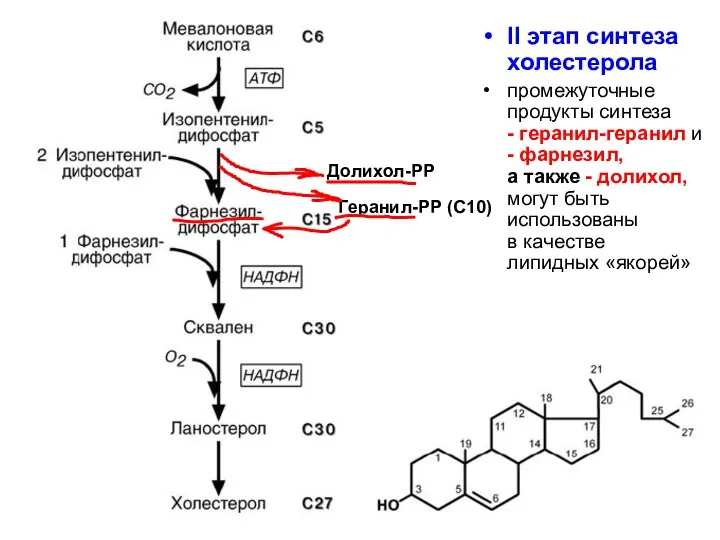
II этап синтеза холестерола
промежуточные продукты синтеза
- геранил-геранил и
- фарнезил,
а
II этап синтеза холестерола
промежуточные продукты синтеза
- геранил-геранил и
- фарнезил,
а
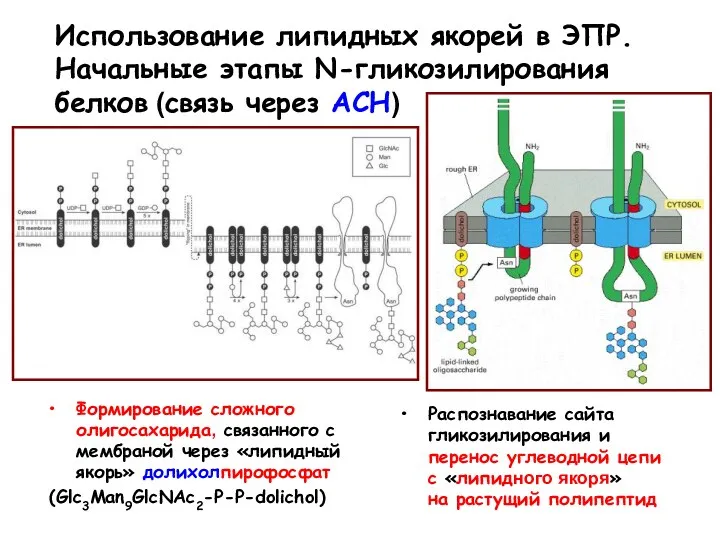
Использование липидных якорей в ЭПР.
Начальные этапы N-гликозилирования
белков (связь через АСН)
Формирование
Использование липидных якорей в ЭПР.
Начальные этапы N-гликозилирования
белков (связь через АСН)
Формирование
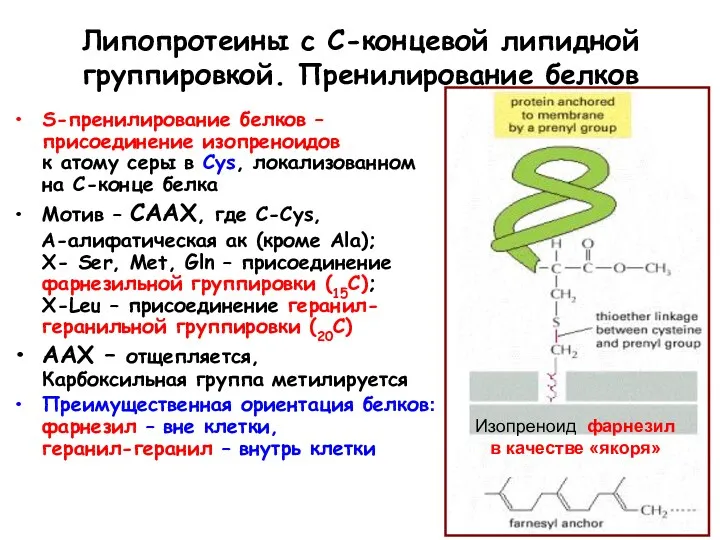
Липопротеины с С-концевой липидной группировкой. Пренилирование белков
S-пренилирование белков – присоединение изопреноидов
Липопротеины с С-концевой липидной группировкой. Пренилирование белков
S-пренилирование белков – присоединение изопреноидов
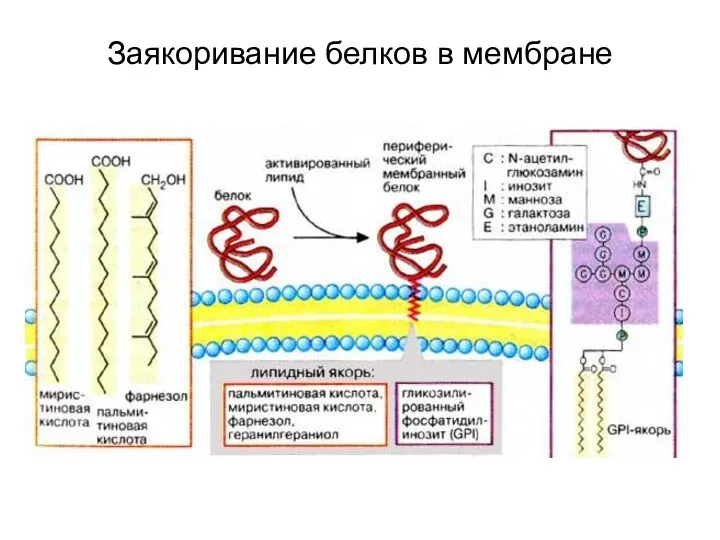
Заякоривание белков в мембране
Заякоривание белков в мембране

Основные виды патологии липидного обмена
Ожирение - избыточное отложение жира чаще алиментарного,
Основные виды патологии липидного обмена
Ожирение - избыточное отложение жира чаще алиментарного,

Осложнения ожирения
заболевание эндокринной системы → дает ослож-
нения сразу на несколько систем
Осложнения ожирения
заболевание эндокринной системы → дает ослож-
нения сразу на несколько систем

Основные виды патологии липидного обмена
Желчекаменная болезнь
Холестерол является компонентом желчных кислот
Основные виды патологии липидного обмена
Желчекаменная болезнь Холестерол является компонентом желчных кислот
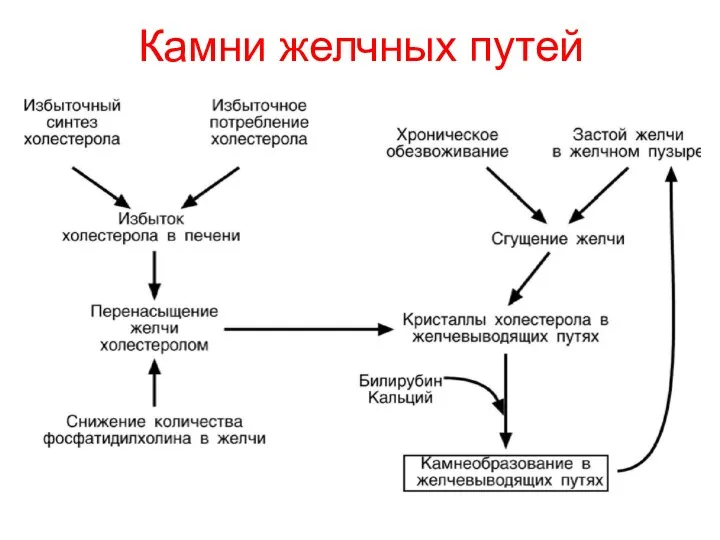
Камни желчных путей
Камни желчных путей

Основные виды патологии липидного обмена
Дислипопротеинемии – могут быть как наследственными, так
Основные виды патологии липидного обмена
Дислипопротеинемии – могут быть как наследственными, так

Нарушения обмена разных ЛП
можно приобрести при ряде заболеваний
(сахарный
Нарушения обмена разных ЛП можно приобрести при ряде заболеваний (сахарный

Типы дислипопротеинемий
I тип: повышена концентрация ХМ
При наследственной недостаточности ЛП-липазы ХМ не
Типы дислипопротеинемий
I тип: повышена концентрация ХМ
При наследственной недостаточности ЛП-липазы ХМ не

II тип: повышено содержание ЛПНП
или суммарно ЛПОНП + ЛПНП
II тип: повышено содержание ЛПНП или суммарно ЛПОНП + ЛПНП

IV тип. В крови повышено содержание ЛПОНП, ХС, ТАГ.
Это характерно
IV тип. В крови повышено содержание ЛПОНП, ХС, ТАГ. Это характерно

Основные виды патологии липидного обмена
Атеросклероз
Причиной атеросклероза может быть наруше-ние как
Основные виды патологии липидного обмена
Атеросклероз
Причиной атеросклероза может быть наруше-ние как
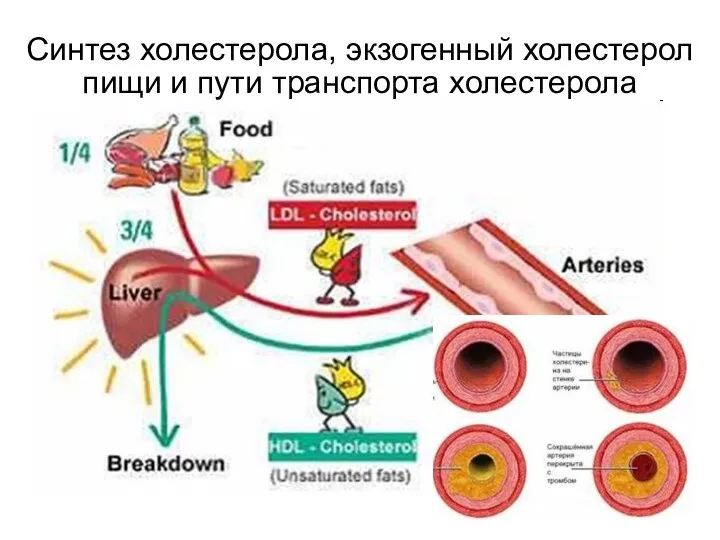
Синтез холестерола, экзогенный холестерол пищи и пути транспорта холестерола
Синтез холестерола, экзогенный холестерол пищи и пути транспорта холестерола

Формирование атеросклероза
При повреждении эндотелия ЛП проникают в сосудистую стенку, поглощаются фагоцитами.
Формирование атеросклероза
При повреждении эндотелия ЛП проникают в сосудистую стенку, поглощаются фагоцитами.

• холестериновый коэффициент атерогенности
k = ХС общ – ХС
• холестериновый коэффициент атерогенности
k = ХС общ – ХС


Все основные виды
патологии липидного обмена
так или иначе взаимосвязаны
Все основные виды
патологии липидного обмена
так или иначе взаимосвязаны
 Хімічні властивості кислот
Хімічні властивості кислот Радикальные реакции. (Лекция 9)
Радикальные реакции. (Лекция 9) Полимерлер-біздің болашағымыз
Полимерлер-біздің болашағымыз Почему нефть называют черным золотом
Почему нефть называют черным золотом Карбоновые кислоты. Изомерия. Физические, химические свойства. Получение, применение
Карбоновые кислоты. Изомерия. Физические, химические свойства. Получение, применение Производство водорода
Производство водорода Общие свойства металлов
Общие свойства металлов Основные положения теории растворов электролитов, используемых в аналитической химии. (Лекция 3)
Основные положения теории растворов электролитов, используемых в аналитической химии. (Лекция 3) Растворы электролитов и неэлектролитов. Ионное произведение воды
Растворы электролитов и неэлектролитов. Ионное произведение воды Минералы и горные породы
Минералы и горные породы Enantioselective Total Synthesis
Enantioselective Total Synthesis Основы химической термодинамики и кинетики химических реакций
Основы химической термодинамики и кинетики химических реакций Общая характеристика галогенов
Общая характеристика галогенов Алканы. Получение, свойства и применение
Алканы. Получение, свойства и применение Термодинамика химических процессов
Термодинамика химических процессов Углеводы (особенности строения, реакционной способности и методы синтеза альдегидо- и кетоспиртов)
Углеводы (особенности строения, реакционной способности и методы синтеза альдегидо- и кетоспиртов) Свойства воды. Оценка качества
Свойства воды. Оценка качества Вычисление массы растворённого вещества, содержащегося в определённой массе раствора с известной массовой долей
Вычисление массы растворённого вещества, содержащегося в определённой массе раствора с известной массовой долей Гетерогенді химиялық реакциялар
Гетерогенді химиялық реакциялар Кобальт. Нахождение в природе. Получение
Кобальт. Нахождение в природе. Получение Группа галогенов в периодической системе
Группа галогенов в периодической системе Подгруппа азота
Подгруппа азота Липиды. Классификация
Липиды. Классификация Химический элемент титан
Химический элемент титан Металлы в природе. Получение
Металлы в природе. Получение Кислоты. Состав кислот
Кислоты. Состав кислот Курс биохимии. Биохимия крови
Курс биохимии. Биохимия крови Гетерофункциональные соединения
Гетерофункциональные соединения