- Цитохромы P-450, b-5

Содержание
- 2. Цитохромы Р450 (КФ 1.14.14.1) – семейство гем-содержащих монооксигеназ, осуществляющих метаболизм ксенобиотиков, в том числе лекарственных препаратов.
- 3. Цитохром Р450 (англ.Cytochrome P450, CYP) Название указывает на то, что он окрашен (Р – от английского
- 4. Использование термина «цитохром» применительно к гемопротеинам класса Р450 нельзя считать удачным, так как функцией цитохромов является
- 5. Молекулярная масса различных цит. Р450 колеблется 44 - 60 кДа. Мономеры гемопротеина состоят из одной полипептидной
- 6. Цит. Р450 из P. putida содержит 4 антипараллельных спиральных участка, смесь спиралей и неупорядоченных структур, перемежающихся
- 7. Все мембранные цитохромы Р450 на N-концевом фрагменте пептидной цепи имеют короткий гидрофобный участок, содержащий от 12
- 8. Цитохромы P450 отсутствуют только у облигатно анаэробных организмов. Описано не менее 11 500 ? белков системы
- 9. Система цит. P450 участвует в окислении многочисленных соединений, как эндогенных, так и экзогенных. Цит. Р450-зависимые монооксигеназы
- 10. Ферменты семейства цитохрома P450 разнообразны и различаются: по функциям, типами ферментативной активности, регуляторами активности (ингибитораи, индукторами).
- 11. Цитохром Р450, наряду с монооксигеназной активностью, может проявлять оксидазную (А.И. Арчаков с сотр.), т.е. катализировать удаление
- 12. Цит. P-450 кодируются суперсемейством генов. У человека в системе цит. Р-450 выявлено 57 генов и более
- 13. В настоящее время известны тысячи изоформ (изоферментов) цит. Р-450. Изоформы, имеющие более 40 % общего аминокислотного
- 14. Цит. P450 катализируют ω-окисление насыщенных жирных кислот (ж.к.), перекисное окисление ненасыщеных ж.к., гидроксилирование стероидных гормонов, желчных
- 15. Общие индукторы
- 16. Фенобарбитал активирует синтез цит. Р450, УДФ-глюкуронилтрансферазы и эпоксид гидролазы. Например, у животных, которым вводили индуктор фенобарбитал,
- 17. В настоящее время описано более 250 химических соединений, вызывающих индукцию микросомальных ферментов. Индукторы монооксигеназных систем разделяются
- 18. Имеются химические вещества, способные ингибировать как ферменты 1-й фазы биотрансформации (изоферменты цитохрома Р-450) и 2-й фазы
- 19. Элетронтранспортные цепи ЭПР 1 цепь включает: 1) цитохром P450, имеет центры связывания для O2 и гидрофобного
- 21. Еще одна схема организации электронтранспортной цепи ЭПР Источником электронов и протонов в цепи является НАДФН+Н+, который
- 22. Пример RH – субстрат цит. Р-450; стрелками показаны реакции переноса электронов. Восстановленную форму цит.-b5 окисляет стеароил-КоА-монооксигеназа,
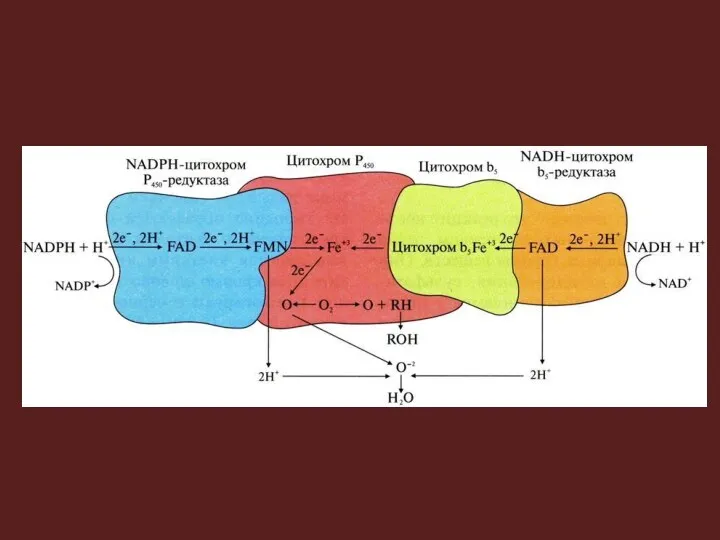
- 23. НАДФ-Н-цитохром Р-450-редуктаза – флавопротеин. Один моль фермента содержит по одному молю флавинмононуклеотида (ФМН) и флавинадениндинуклеотида (ФАД).
- 24. Механизм гидроксилирования субстрата при участии цитохрома Р-450 Условно можно выделить 5 этапов: 1. окисляемое вещество (S)
- 25. МОДЕЛЬ 2. Упрощенная схема гидроксилирования ксенобиотиков микросомальными монооксигеназами ( цитохром Р-450)
- 26. В отличие от митохондриальной дыхательной цепи, в монооксигеназной цепи при переносе электронов не происходит аккумулирования энергии
- 28. Цитохромы Р-450 являются мембранными белками и при исследовании их каталитической активности требуется сложное реконструирование монооксигеназной системы
- 29. В настоящее время разработаны электрохимические биосенсорные системы на основе иммобилизованных на электроде цитохромов Р-450. Электрокаталитическая реакция
- 30. Зависимость изменения каталитического тока при амперметрическом титровании цит. Р-450 3А4 (CYP 3А4) тестостероном при контролируемом напряжении
- 31. Разработка биосенсоров на основе электрохимических цитохром Р450-содержащих систем позволяет выявлять субстраты (ксенобиотики), исследовать эффекты лекарственных препаратов
- 32. Преимущества электрохимического метода исследования цитохром Р450-монооксигеназной системы 1) электрохимическая система не требует использования дорогостоящих и неустойчивых
- 33. Методы оценки состояния системы биотранс- формации ксенобиотиков: Высокоэффективная жидкостная хроматография (ВЭЖХ). Дает возможность исследовать аналиты в
- 34. 2) ПЦР-ПДРФ анализ полиморфизма и мутантных форм генов цит.Р-450. Результаты анализа полиморфизма длины рестрикционных фрагментов (ПДРФ-анализ)
- 35. 3) ДНК-чипы Позволяют одновременно определять очень большое количество полиморфизмов в одной пробе. На твердом чипе очень
- 36. 4) Компьютерные программы для моделирования взаимодействия лигандов с цитохромами Р450 Для изучения взаимодействия субстрата и фермента
- 37. При описании взаимодействия цит.Р-450 и лигандов оценивается роль пространственных и энергетических факторов, т.к. вклад этих факторов
- 38. Р-ция наз. региоспецифичной, если в качестве единств. продукта (в пределах ошибки) образуется один из двух или
- 39. QSAR модели. Количественные взаимоотношения структура-активность (QSAR - англ. сокр. от Quantitative Structure-Activity Relationships) позволяют по описанию
- 40. Суперсемейство цит. Р-450 катализирует большое количество реакций, проходящих по разным механизмам, поэтому классические QSAR методы не
- 41. Основные цитохромы Р450, ответственные за метаболизм лекарств в организме человека исследуемые in silico – это подсемейство
- 42. Цитохром b5 – гемопротеин, участвует в разнообразных биохимических окислительно-восстановительных реакциях в качестве переносчика электронов. Молекула микросомального
- 43. Гидрофобный домен цитохрома b5 заякорен в липидном бислое (ЭПР или митохондриальной), спирализован, образован остатками аминокислот C-
- 44. Цитохром b5 наружной мембраны митохондриий, по сравнению с микросомальным обладает более низким редокс-потенциалом, молекула более устойчива
- 45. Роль цитохрома b5 в реакциях, катализируемых изоформами системы цитохрома Р-450. Возможные механизмы стимулирующего влияния цит. b5
- 46. цит. b5 взаимодействует с цит. Р-450 с образованием комплекса двух гемопротеинов и последующей передачей двух электронов
- 47. Влияние цит. b5 на изменение скорости реакции, спектра метаболитов и образование активных форм кислорода в реакциях
- 48. цит. b5 у разных видов (человек и хомячок) может не изменять скорости окисления соединения (нитрозамин) или
- 49. Влияние цитохрома b5 на изменение скорости реакции, спектра метаболитов и образование активных форм кислорода в реакциях
- 51. Скачать презентацию
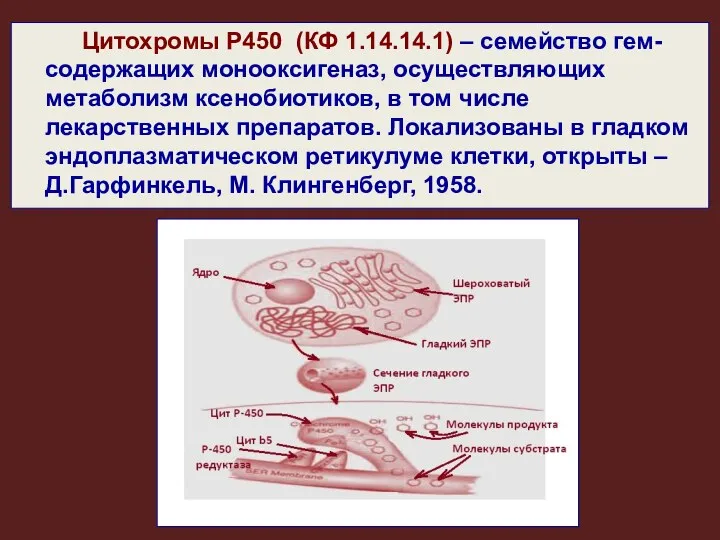
Цитохромы Р450 (КФ 1.14.14.1) – семейство гем-содержащих монооксигеназ, осуществляющих метаболизм
Цитохромы Р450 (КФ 1.14.14.1) – семейство гем-содержащих монооксигеназ, осуществляющих метаболизм

Цитохром Р450 (англ.Cytochrome P450, CYP)
Название указывает на то, что он окрашен
Название указывает на то, что он окрашен

Использование термина «цитохром» применительно к гемопротеинам класса Р450 нельзя считать
Использование термина «цитохром» применительно к гемопротеинам класса Р450 нельзя считать

Молекулярная масса различных цит. Р450 колеблется 44 - 60 кДа.
Мономеры
Молекулярная масса различных цит. Р450 колеблется 44 - 60 кДа.
Мономеры

Цит. Р450 из P. putida содержит
4 антипараллельных спиральных участка,
Цит. Р450 из P. putida содержит
4 антипараллельных спиральных участка,

Все мембранные цитохромы Р450 на N-концевом фрагменте пептидной цепи имеют короткий
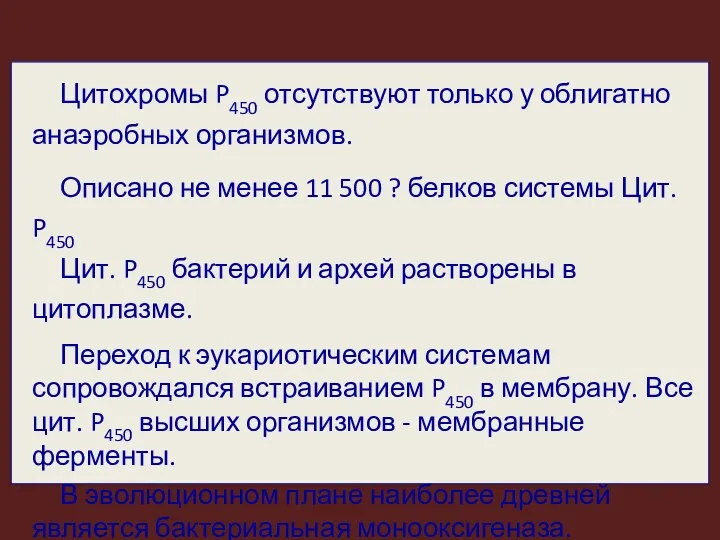
Цитохромы P450 отсутствуют только у облигатно анаэробных организмов.
Описано не менее
Описано не менее

Система цит. P450 участвует в окислении многочисленных соединений, как эндогенных,
Система цит. P450 участвует в окислении многочисленных соединений, как эндогенных,

Ферменты семейства цитохрома P450 разнообразны и различаются:
по функциям,
типами
Ферменты семейства цитохрома P450 разнообразны и различаются:
по функциям,
типами

Цитохром Р450, наряду с монооксигеназной активностью, может проявлять оксидазную (А.И.
Цитохром Р450, наряду с монооксигеназной активностью, может проявлять оксидазную (А.И.

Цит. P-450 кодируются суперсемейством генов.
У человека в системе цит.
Цит. P-450 кодируются суперсемейством генов.
У человека в системе цит.

В настоящее время известны тысячи изоформ (изоферментов) цит. Р-450.
Изоформы, имеющие более
Изоформы, имеющие более

Цит. P450 катализируют ω-окисление насыщенных жирных кислот (ж.к.), перекисное окисление
Цит. P450 катализируют ω-окисление насыщенных жирных кислот (ж.к.), перекисное окисление
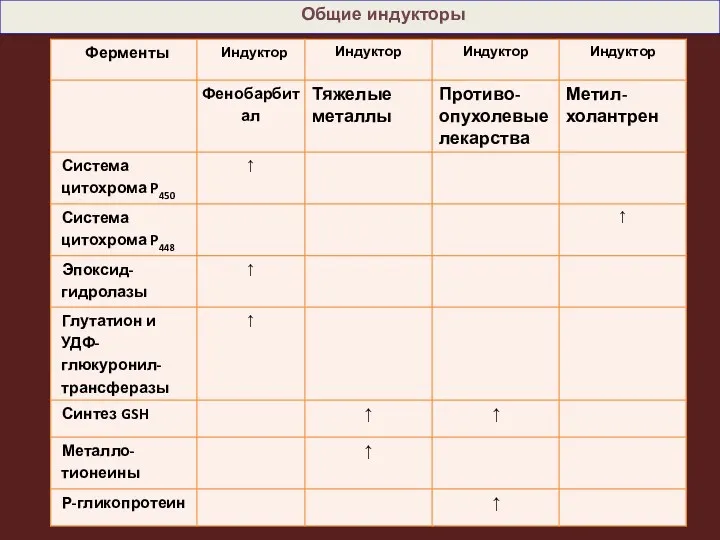
Общие индукторы
Общие индукторы

Фенобарбитал активирует синтез цит. Р450, УДФ-глюкуронилтрансферазы и эпоксид гидролазы.
Например,
Фенобарбитал активирует синтез цит. Р450, УДФ-глюкуронилтрансферазы и эпоксид гидролазы.
Например,

В настоящее время описано более 250 химических соединений, вызывающих индукцию
В настоящее время описано более 250 химических соединений, вызывающих индукцию

Имеются химические вещества, способные ингибировать как ферменты 1-й фазы биотрансформации (изоферменты

Элетронтранспортные цепи ЭПР
1 цепь включает:
1) цитохром P450, имеет центры связывания для
Элетронтранспортные цепи ЭПР
1 цепь включает:
1) цитохром P450, имеет центры связывания для
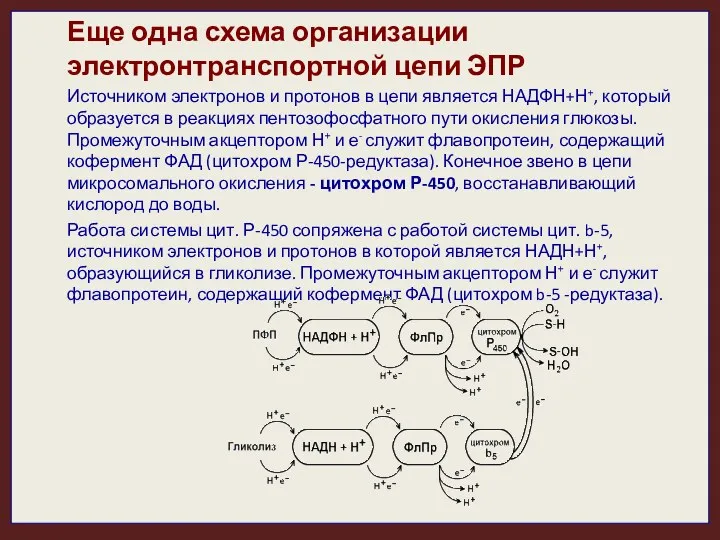
Еще одна схема организации электронтранспортной цепи ЭПР
Источником электронов и протонов в
Еще одна схема организации электронтранспортной цепи ЭПР
Источником электронов и протонов в
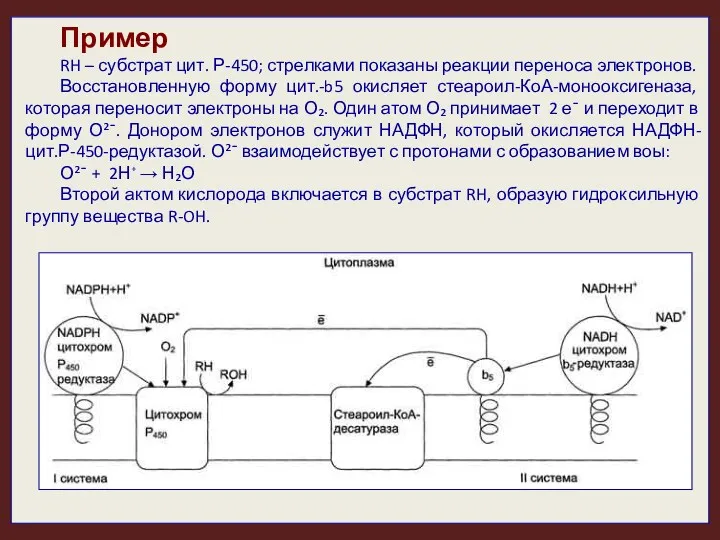
Пример
RH – субстрат цит. Р-450; стрелками показаны реакции переноса электронов.
Восстановленную
Пример
RH – субстрат цит. Р-450; стрелками показаны реакции переноса электронов.
Восстановленную

НАДФ-Н-цитохром Р-450-редуктаза – флавопротеин.
Один моль фермента содержит по одному молю
Один моль фермента содержит по одному молю
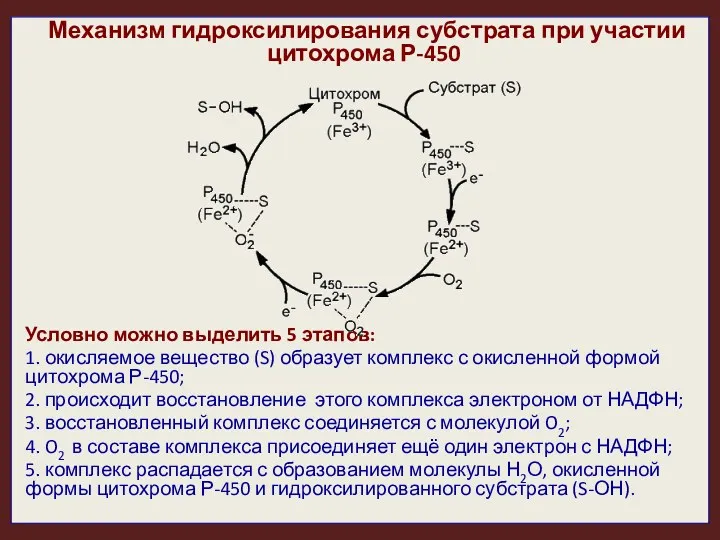
Механизм гидроксилирования субстрата при участии цитохрома Р-450
Условно можно выделить
Механизм гидроксилирования субстрата при участии цитохрома Р-450
Условно можно выделить
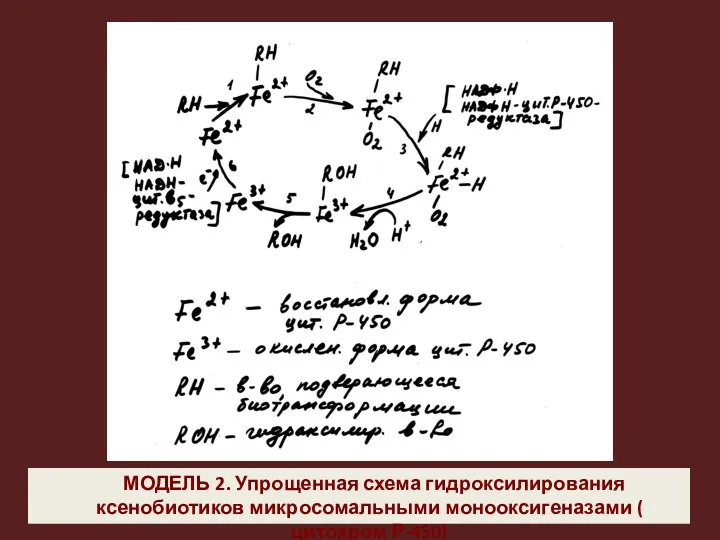
МОДЕЛЬ 2. Упрощенная схема гидроксилирования ксенобиотиков микросомальными монооксигеназами ( цитохром
МОДЕЛЬ 2. Упрощенная схема гидроксилирования ксенобиотиков микросомальными монооксигеназами ( цитохром

В отличие от митохондриальной дыхательной цепи, в монооксигеназной цепи при переносе
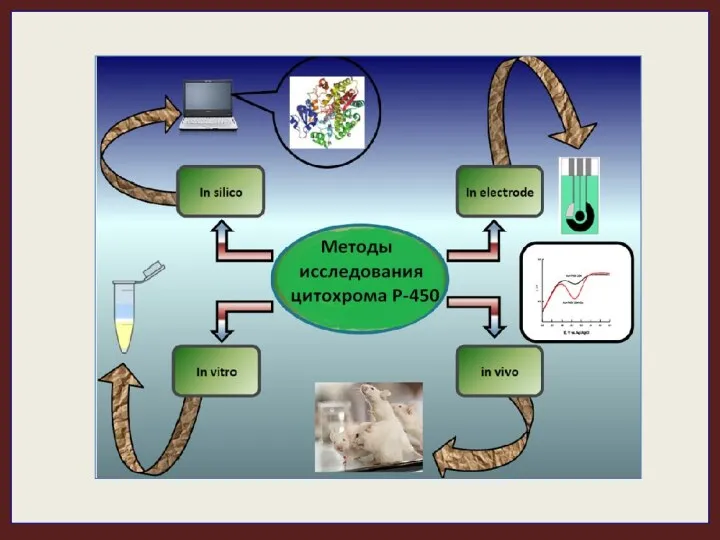

Цитохромы Р-450 являются мембранными белками и при исследовании их каталитической активности

В настоящее время разработаны электрохимические биосенсорные системы на основе иммобилизованных на
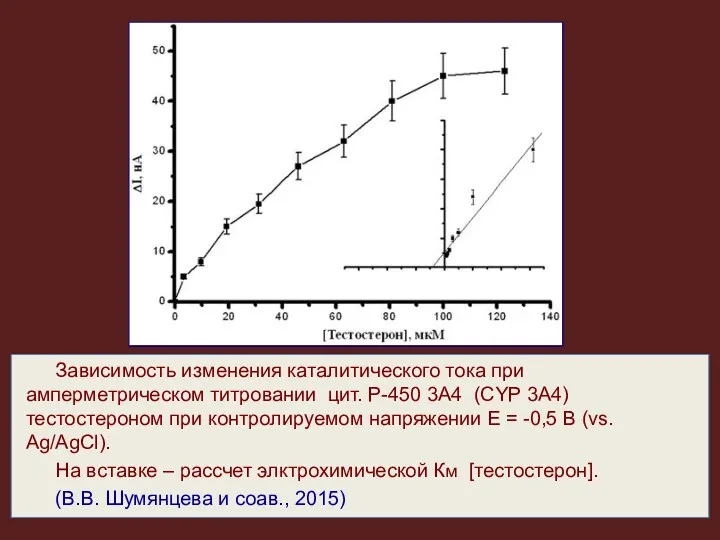
Зависимость изменения каталитического тока при амперметрическом титровании цит. Р-450 3А4 (CYP
Зависимость изменения каталитического тока при амперметрическом титровании цит. Р-450 3А4 (CYP

Разработка биосенсоров на основе электрохимических цитохром Р450-содержащих систем позволяет выявлять субстраты

Преимущества электрохимического метода исследования цитохром Р450-монооксигеназной системы
1) электрохимическая система не требует
1) электрохимическая система не требует

Методы оценки состояния системы биотранс- формации ксенобиотиков:
Высокоэффективная жидкостная хроматография (ВЭЖХ).
Дает
Высокоэффективная жидкостная хроматография (ВЭЖХ).
Дает
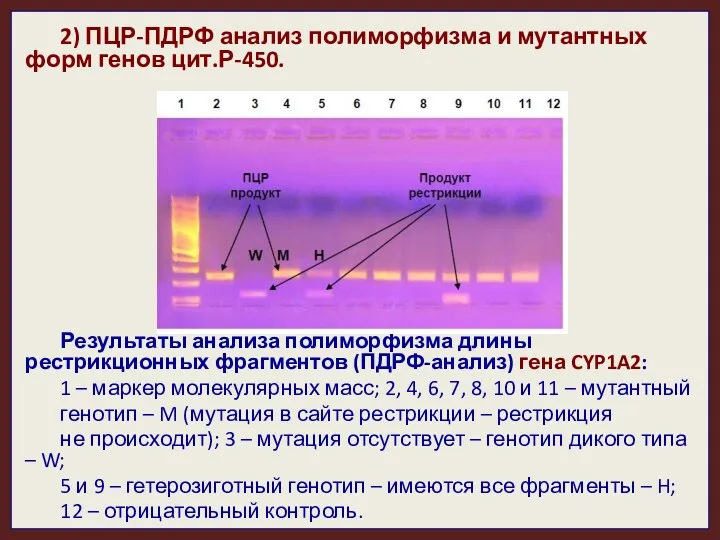
2) ПЦР-ПДРФ анализ полиморфизма и мутантных форм генов цит.Р-450.
Результаты анализа полиморфизма
Результаты анализа полиморфизма

3) ДНК-чипы
Позволяют одновременно определять очень большое количество полиморфизмов в одной
Позволяют одновременно определять очень большое количество полиморфизмов в одной

4) Компьютерные программы для моделирования взаимодействия лигандов с цитохромами Р450
Для изучения
Для изучения

При описании взаимодействия цит.Р-450 и лигандов оценивается роль пространственных и энергетических

Р-ция наз. региоспецифичной, если в качестве единств. продукта (в пределах ошибки)

QSAR модели. Количественные взаимоотношения структура-активность (QSAR - англ. сокр. от Quantitative

Суперсемейство цит. Р-450 катализирует большое количество реакций, проходящих по разным механизмам,

Основные цитохромы Р450, ответственные за метаболизм лекарств в организме человека исследуемые
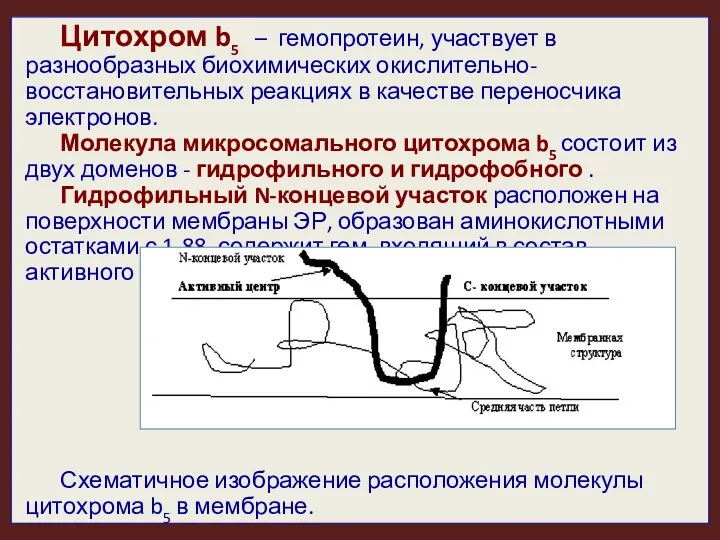
Цитохром b5 – гемопротеин, участвует в разнообразных биохимических окислительно-восстановительных реакциях в
Цитохром b5 – гемопротеин, участвует в разнообразных биохимических окислительно-восстановительных реакциях в

Гидрофобный домен цитохрома b5 заякорен в липидном бислое (ЭПР или митохондриальной),

Цитохром b5 наружной мембраны митохондриий, по сравнению с микросомальным обладает более

Роль цитохрома b5 в реакциях, катализируемых изоформами системы цитохрома Р-450.
Возможные механизмы
Возможные механизмы

цит. b5 взаимодействует с цит. Р-450 с образованием комплекса двух гемопротеинов

Влияние цит. b5 на изменение скорости реакции, спектра метаболитов и образование

цит. b5 у разных видов (человек и хомячок) может не изменять

Влияние цитохрома b5 на изменение скорости реакции, спектра метаболитов и образование
Влияние цитохрома b5 на изменение скорости реакции, спектра метаболитов и образование
 Гидроксипроизводные углеводородов. Спирты. Фенолы
Гидроксипроизводные углеводородов. Спирты. Фенолы Механохимиялық активтеу әдісі
Механохимиялық активтеу әдісі Табиғи нанокристалдар
Табиғи нанокристалдар Соли. Определение солей
Соли. Определение солей Класифікація та властивості оксидів
Класифікація та властивості оксидів Кристаллы. Формы
Кристаллы. Формы Окислительно-восстановительные реакции
Окислительно-восстановительные реакции Растворы электролитов
Растворы электролитов Общая химия.(Лекция№1). Основные законы и понятия химии
Общая химия.(Лекция№1). Основные законы и понятия химии Фосфор. Урок в 9 классе
Фосфор. Урок в 9 классе Основания. Обобщение
Основания. Обобщение Кремний и его соединения. Силикатная промышленность
Кремний и его соединения. Силикатная промышленность Нефть и ее роль в мировой экономике
Нефть и ее роль в мировой экономике Көмірсутектерді пиролиздеу арқылы қарапайым олефиндерді алу
Көмірсутектерді пиролиздеу арқылы қарапайым олефиндерді алу Кривые титрования: построение и анализ (лекция 6)
Кривые титрования: построение и анализ (лекция 6) Cromatografia ionică
Cromatografia ionică Техника безопасности для учащихся в кабинете химии
Техника безопасности для учащихся в кабинете химии Окисно-відновні реакції. Процеси окиснення, відновлення, окисник та відновник
Окисно-відновні реакції. Процеси окиснення, відновлення, окисник та відновник Термокаталитические процессы. Каталитический крекинг. Классификация и основные факторы процесса
Термокаталитические процессы. Каталитический крекинг. Классификация и основные факторы процесса Ағзаның барлық жасушасындағы белок алмасу. Аминқышқылдарының дезаминденуі, пурин және пиримидиндердің ыдырауынан
Ағзаның барлық жасушасындағы белок алмасу. Аминқышқылдарының дезаминденуі, пурин және пиримидиндердің ыдырауынан Вода. Состав и строение молекулы
Вода. Состав и строение молекулы Негізгі топша элементтерінің химиясы
Негізгі топша элементтерінің химиясы Виды химической связи
Виды химической связи Одноатомные спирты. Простые эфиры
Одноатомные спирты. Простые эфиры Органическая химия в ряду других наук
Органическая химия в ряду других наук Физико-химические методы в анализе ЛВ. Метод ионометрии в анализе ЛВ. Определение спирта в фармацевтических препаратах
Физико-химические методы в анализе ЛВ. Метод ионометрии в анализе ЛВ. Определение спирта в фармацевтических препаратах Періодична система хімічних елементів. Хімія. 8 клас
Періодична система хімічних елементів. Хімія. 8 клас Жидкие кристаллы
Жидкие кристаллы