- Аминокислоты. Пептиды. Белки

Содержание
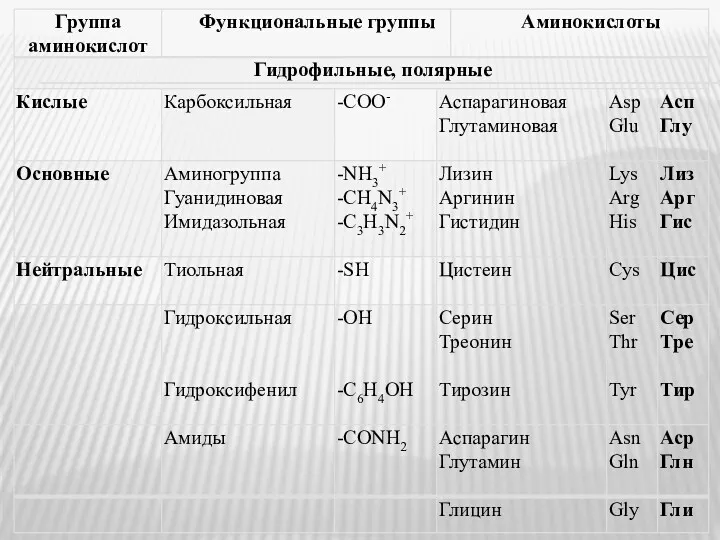
- 2. СТРУКТУРА И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ 20 аминокислот входят в состав белков (протеиногенные аминокислоты). Это α-аминокислоты, в
- 3. По структуре боковой группы R аминокислоты подразделяются на: моноаминомонокарбоновые алифатические (глицин, аланин, валин, лейцин, изолейцин); моноаминодикарбоновые
- 5. *Тирозин, или гидроксифенилаланин – ароматическая, гидрофильная, полярная аминокислота.
- 6. Протеиногенные аминокислоты делятся на: незаменимые – не могут синтезироваться в организме человека (треонин, метионин, валин, лейцин,
- 7. α-Аминоксилоты (кроме глицина) имеют в структуре хиральные (асимметричные) атомы С. Это обусловливает существование двух энантиомеров –
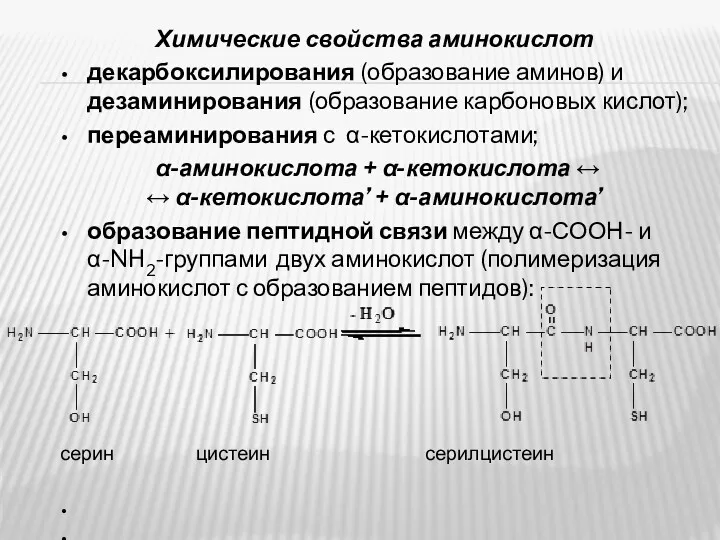
- 8. Химические свойства аминокислот декарбоксилирования (образование аминов) и дезаминирования (образование карбоновых кислот); переаминирования с α-кетокислотами; α-аминокислота +
- 9. образования амидов и сложных эфиров; взаимодействие аминогрупп с альдегидами (образование шиффовых оснований); образование N-гликозидов (при взаимодействии
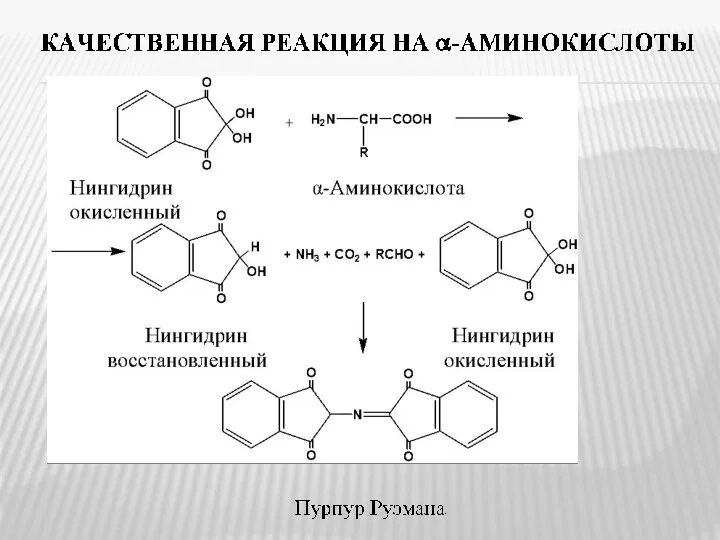
- 10. Универсальной качественной реакцией на α-аминокислоты, является их взаимодействие с нингидрином, сопровождающееся образованием окрашенного продукта фиолетового цвета
- 12. Амфотерные свойства аминокислот α-Аминокислоты в водных растворах существуют преимуществненно в виде биполярных, или цвиттер-ионов:
- 13. Степень диссоциации ионогенных групп зависит от рН. Значение рН раствора, при котором суммарный заряд молекулы аминокислоты
- 14. Значение рН водного раствора химически чистой аминокислоты называется изоионной точкой. Значения изоэлектрической и изоионной точек в
- 15. Заряд аминокислоты в растворе зависит от его рН Аминокислоты в растворах при любых значениях рН (кроме
- 16. Аминокислотные остатки в молекуле белка соединены пептидными связями. Длина пептидной связи = 0,132 нм длина одинарной
- 17. Свойства пептидной связи: пептидная группа жесткая планарная (плоская) структура и вращение вокруг пептидной связи невозможно; пептидная
- 18. По числу аминокислотных остатков: олигопептиды (до 10 аминокислотных остатков); полипептиды (от 10 до 50 аминокислотных остатков).
- 19. Полипептиды, состоящие более, чем из 50 аминокислотных остатков, относятся к белкам, или протеинам. В структуре белковой
- 22. В зависимости от степени асимметрии молекулы белка, имеющие пространственную структуру (конформацию), подразделяются на: - глобулярные (при
- 23. Формирование третичной структуры приводит к образованию функционально активной, или нативной, белковой структуры.
- 24. Физико-химические свойства белков Большинство белков – это водорастворимые вещества. В растворах белки проявляют коллоидные свойства и
- 25. Белки способны взаимодействовать и с катионами, и с анионами. Способность белков взаимодействовать с различными заряженными веществами
- 26. Денатурация – изменение пространственной структуры, которая происходит в связи с разрывом связей, поддерживающих и образующих пространственную
- 27. КЛАССИФИКАЦИЯ БЕЛКОВ В зависимости от состава белки делятся на простые и сложные. Простые белки состоят только
- 28. Сложные белки кроме белковой части имеют структуры небелковой природы. Хромопротеины –окрашенные белки: гемопротеины, флавопротеины, родопсин и
- 29. Функции белков Каталитическая функция. Структурная функция. Транспортная функция Защитная функция. Регуляторная функция. Двигательная функция.
- 30. ФЕРМЕНТЫ Ферменты - природные биокатализаторы белковой природы.
- 31. СВОЙСТВА ФЕРМЕНТОВ Общие со всеми катализаторами: 1. способность катализировать только термодинамически возможные процессы. 2. ускорение наступления
- 32. Специфические свойства: 1. более высокая активность ферментов по сравнению с неорганическими катализаторами. 2. высокую специфичность действия
- 33. СТРУКТУРА ФЕРМЕНТОВ Простые ферменты – однокомпонентные, состоят только из полипептидной части; Сложные ферменты (холофермент) – двухкомпонентные,
- 34. Классификация ферментов
- 35. Единицы и формы выражения активности ферментов 1 катал (каt) – количество фермента, которое катализирует превращение 1
- 38. Скачать презентацию

СТРУКТУРА И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ
20 аминокислот входят в состав белков (протеиногенные
СТРУКТУРА И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ
20 аминокислот входят в состав белков (протеиногенные

По структуре боковой группы R аминокислоты подразделяются на:
моноаминомонокарбоновые алифатические (глицин, аланин,
По структуре боковой группы R аминокислоты подразделяются на:
моноаминомонокарбоновые алифатические (глицин, аланин,

*Тирозин, или гидроксифенилаланин – ароматическая, гидрофильная, полярная аминокислота.
*Тирозин, или гидроксифенилаланин – ароматическая, гидрофильная, полярная аминокислота.

Протеиногенные аминокислоты делятся на:
незаменимые – не могут синтезироваться в организме человека
незаменимые – не могут синтезироваться в организме человека

α-Аминоксилоты (кроме глицина) имеют в структуре хиральные (асимметричные) атомы С.
Это обусловливает
α-Аминоксилоты (кроме глицина) имеют в структуре хиральные (асимметричные) атомы С.
Это обусловливает
Химические свойства аминокислот
декарбоксилирования (образование аминов) и дезаминирования (образование карбоновых кислот);
переаминирования
Химические свойства аминокислот
декарбоксилирования (образование аминов) и дезаминирования (образование карбоновых кислот);
переаминирования

образования амидов и сложных эфиров;
взаимодействие аминогрупп с альдегидами (образование шиффовых оснований);
образование
образования амидов и сложных эфиров;
взаимодействие аминогрупп с альдегидами (образование шиффовых оснований);
образование

Универсальной качественной реакцией на α-аминокислоты, является их взаимодействие с нингидрином, сопровождающееся
Универсальной качественной реакцией на α-аминокислоты, является их взаимодействие с нингидрином, сопровождающееся
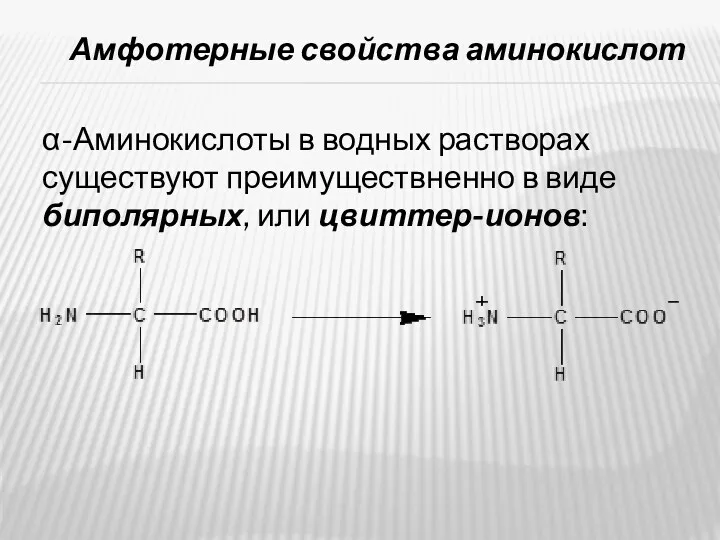
Амфотерные свойства аминокислот
α-Аминокислоты в водных растворах существуют преимуществненно в виде биполярных,
Амфотерные свойства аминокислот
α-Аминокислоты в водных растворах существуют преимуществненно в виде биполярных,

Степень диссоциации ионогенных групп зависит от рН.
Значение рН раствора, при котором
Степень диссоциации ионогенных групп зависит от рН.
Значение рН раствора, при котором

Значение рН водного раствора химически чистой аминокислоты называется изоионной точкой.
Значения
Значение рН водного раствора химически чистой аминокислоты называется изоионной точкой.
Значения

Заряд аминокислоты в растворе зависит от его рН
Аминокислоты в растворах при
Заряд аминокислоты в растворе зависит от его рН
Аминокислоты в растворах при
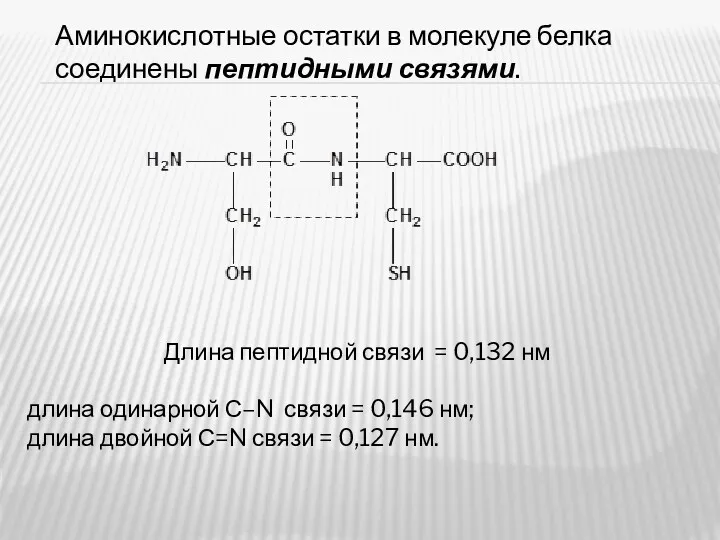
Аминокислотные остатки в молекуле белка соединены пептидными связями.
Длина пептидной связи
Аминокислотные остатки в молекуле белка соединены пептидными связями.
Длина пептидной связи

Свойства пептидной связи:
пептидная группа жесткая планарная (плоская) структура и вращение вокруг
Свойства пептидной связи:
пептидная группа жесткая планарная (плоская) структура и вращение вокруг

По числу аминокислотных остатков:
олигопептиды (до 10 аминокислотных остатков);
полипептиды (от 10
По числу аминокислотных остатков:
олигопептиды (до 10 аминокислотных остатков);
полипептиды (от 10

Полипептиды, состоящие более, чем из 50 аминокислотных остатков, относятся к белкам,
Полипептиды, состоящие более, чем из 50 аминокислотных остатков, относятся к белкам,
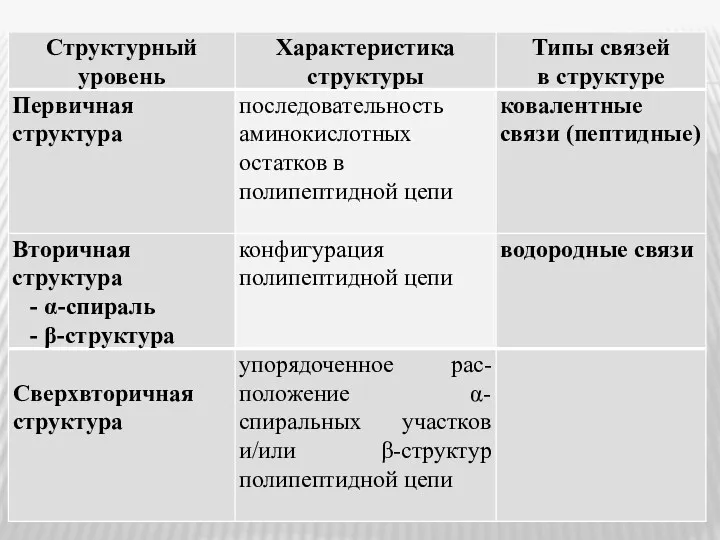


В зависимости от степени асимметрии молекулы белка, имеющие пространственную структуру (конформацию),
В зависимости от степени асимметрии молекулы белка, имеющие пространственную структуру (конформацию),

Формирование третичной структуры приводит к образованию функционально активной, или нативной, белковой
Формирование третичной структуры приводит к образованию функционально активной, или нативной, белковой

Физико-химические свойства белков
Большинство белков – это водорастворимые вещества.
В растворах белки
Физико-химические свойства белков
Большинство белков – это водорастворимые вещества.
В растворах белки

Белки способны взаимодействовать и с катионами, и с анионами.
Способность белков
Белки способны взаимодействовать и с катионами, и с анионами.
Способность белков

Денатурация – изменение пространственной структуры, которая происходит в связи с разрывом
Денатурация – изменение пространственной структуры, которая происходит в связи с разрывом

КЛАССИФИКАЦИЯ БЕЛКОВ
В зависимости от состава белки делятся
на простые и сложные.
Простые белки
КЛАССИФИКАЦИЯ БЕЛКОВ
В зависимости от состава белки делятся
на простые и сложные.
Простые белки

Сложные белки кроме белковой части имеют
структуры небелковой природы.
Хромопротеины –окрашенные
Сложные белки кроме белковой части имеют
структуры небелковой природы.
Хромопротеины –окрашенные

Функции белков
Каталитическая функция.
Структурная функция.
Транспортная функция
Защитная функция.
Регуляторная функция.
Двигательная функция.
Функции белков
Каталитическая функция.
Структурная функция.
Транспортная функция
Защитная функция.
Регуляторная функция.
Двигательная функция.

ФЕРМЕНТЫ
Ферменты - природные биокатализаторы белковой природы.
ФЕРМЕНТЫ
Ферменты - природные биокатализаторы белковой природы.

СВОЙСТВА ФЕРМЕНТОВ
Общие со всеми катализаторами:
1. способность катализировать только термодинамически возможные процессы.
2.
СВОЙСТВА ФЕРМЕНТОВ
Общие со всеми катализаторами:
1. способность катализировать только термодинамически возможные процессы.
2.

Специфические свойства:
1. более высокая активность ферментов по сравнению с неорганическими катализаторами.
Специфические свойства:
1. более высокая активность ферментов по сравнению с неорганическими катализаторами.

СТРУКТУРА ФЕРМЕНТОВ
Простые ферменты – однокомпонентные, состоят только из полипептидной части;
Сложные ферменты
СТРУКТУРА ФЕРМЕНТОВ
Простые ферменты – однокомпонентные, состоят только из полипептидной части;
Сложные ферменты

Классификация ферментов
Классификация ферментов

Единицы и формы выражения
активности ферментов
1 катал (каt) – количество фермента,
Единицы и формы выражения
активности ферментов
1 катал (каt) – количество фермента,
 Химические реакторы. Лекция №10
Химические реакторы. Лекция №10 Химические свойства солей
Химические свойства солей Кислоты: классификация и химические свойства
Кислоты: классификация и химические свойства Гравиметрический метод анализа
Гравиметрический метод анализа Энергетика химических процессов. Энтропия и энергия Гиббса
Энергетика химических процессов. Энтропия и энергия Гиббса Различные теории кислот и оснований
Различные теории кислот и оснований Особенности выбора связующих и наполнителей для производства деталей машин из ПКМ
Особенности выбора связующих и наполнителей для производства деталей машин из ПКМ Общая схема радиолиза алифатических углеводородов. Радиолиз органических кислот. Радиолиз кетонов эфиров
Общая схема радиолиза алифатических углеводородов. Радиолиз органических кислот. Радиолиз кетонов эфиров Композиционные материалы для изоляции электрических машин
Композиционные материалы для изоляции электрических машин Предмет и задачи химии. Вещества и их свойства. (Продолжение)
Предмет и задачи химии. Вещества и их свойства. (Продолжение) Галогены. Положение в периодической системе. Химические свойства
Галогены. Положение в периодической системе. Химические свойства Амины. Общая характеристика, номенклатура, изомерия. Получение. Физико-химические свойства. Отдельные представители
Амины. Общая характеристика, номенклатура, изомерия. Получение. Физико-химические свойства. Отдельные представители Ионные уравнения
Ионные уравнения S-элементтердің жалпы сипаттамасы
S-элементтердің жалпы сипаттамасы История нефти. Свойства нефти. Классификация нефтей
История нефти. Свойства нефти. Классификация нефтей Алкины. Самостоятельная работа
Алкины. Самостоятельная работа Признаки химических реакций. (Практическая работа 5,4)
Признаки химических реакций. (Практическая работа 5,4) Роль металлов в нашей жизни
Роль металлов в нашей жизни Насыщенные углеводороды
Насыщенные углеводороды Полимерные растворы. Условия образования
Полимерные растворы. Условия образования Сушка в химической промышленности. Свойства влажного воздуха
Сушка в химической промышленности. Свойства влажного воздуха Кислотно-основное равновесие в процессах жизнедеятельности. Ионное произведение воды. Водородный показатель
Кислотно-основное равновесие в процессах жизнедеятельности. Ионное произведение воды. Водородный показатель Химический эквивалент. Формульные единицы
Химический эквивалент. Формульные единицы i9tjjvxuvf7fj7k4drnlmj56tgok
i9tjjvxuvf7fj7k4drnlmj56tgok Введение в кристаллографию
Введение в кристаллографию Количество вещества, число Авогадро, молярная масса, молярный объём, уравнение связи
Количество вещества, число Авогадро, молярная масса, молярный объём, уравнение связи Строительное материаловедение. (Лекции 1-2)
Строительное материаловедение. (Лекции 1-2) Марганец. Железо
Марганец. Железо