- Биохимическая трансформация веществ (2)

Содержание
- 2. Биотрансформация чужеродных соединений происходит во всех биосистемах (органоид, клетка, ткань, орган, организм, надорганизменные биосистемы – биогеоценозы,
- 3. Фазы биотрансформации Реакции 1-й фазы Реакции 2-й фазы (реакции синтеза) конъюгация гидролиз, восстановление, окисление Конъюгация с
- 4. СИСТЕМА ОБЕЗВРЕЖИВАНИЯ ЛИПОФИЛЬНЫХ КСЕНОБИОТИКОВ
- 5. Основные пути биотрансформации чужеродных соединений. I ФАЗА Окисление: а) микросомальное – алифатичекое или ароматическое гидроксилирование, –
- 6. Восстановление: а) восстановление нитросоединений, азосоединений микросомальными ферментами; б) микросомальное восстановительное галогенирование; в) немикросомальное восстановление. Гидролиз с
- 7. Ферменты биотрансформации ксенобиотиков присутствуют, в основном, в микросомах и в цитозоле, незначительная часть – в митохондриях,
- 8. Ферментативные реакции 1-й фазы биотрансформации карбоксиэстераза, ацетилхолинэстераза псевдохолинэстераза эпоксидная гидролаза l фаза метаболизма - этап биотрансформации,
- 9. Пример. Гидролиз эпоксидов (связи углерод-кислород в оксирановом кольце) осуществляется эпоксидгидратазой. В результате реакции образуются дигидродиолы. (Стильбеноксид
- 10. 2 . Реакции восстановления. Восстанавливаются некоторые металлы, альдегиды, кетоны, дисульфиды, сульфоксиды, хиноны, алкены, азо- и нитросоединения.
- 11. Восстановление азо- и нитросоединений – осуществляют цитохром Р450, НАДФН-хинон оксидоредуктазы.
- 12. Восстановление карбонильных соединений Алкогольдегидрогеназа, группа ферментов — карбонильные редуктазы
- 13. Восстановление дисульфидов – ферменты: глутатионредуктаза, глутатион - S-трансфераза, или неферментативно
- 14. Восстановление сульфоксидов – осуществляют цитохром Р450, необходим НАДФН Восстановление хинонов - ферменты: НАДФН-хиноноксидоредуктаза, необходимы флавопротеины цитозоля
- 15. Дегалогенирование: окислительное дегалогенирование двойное дегалогенирование дегидрогалогенирование ХЛОРОФОРМ ФОСГЕН CHCl3→ ClCOCl +HCl Дегалогенирование
- 16. Галотан (фторотан) – высокоактивное ингаляционное средство для наркоза, подвергается различным метаболическим превращениям в организме животных и
- 17. Последние данные указывают, что метаболизм галотана условно делится на два пути: включает окисление молекулы с образованием
- 18. 3. Реакции окисления. Альдегиддегидрогеназы (АлДГ) - окисление альдегидов до карбоновых кислот (кофактор НАД+). Дигидродиолдегидрогеназы - окисление
- 19. Пероксидазы 1) обезвреживают пероксиды; 2) могут превращать ксенобиотики в токсичные метаболиты; 3) могут осуществлять прямой перенос
- 20. Флавинмонооксигеназы - окисляют нуклеофильный азот, серу и фосфор в молекулах ксенобиотиков.
- 21. Цитохром Р450 катализирует реакции окисления: -гидроксилирование алифатических и ароматических углеводородов; эпоксидирование двойной связи; окисление гетероатомов (О-,
- 22. Реакции N-деалкилирования
- 23. Реакции O-деалкилирования
- 24. Реакции S-деалкилирования
- 25. Эпоксидирование и гидроксилирование ароматических соединений Эпоксидирование алифатических и алициклических соединений
- 26. Гидроксилирование циклических ароматических углеводородов
- 27. Гидроксилирование циклических предельных углеводородов
- 28. Гидроксилирование гетероциклических углеводородов
- 29. Гидроксилирование алифатических соединений: а) предельных углеводородов
- 30. б) алкильной боковой цепи
- 31. N-окисление (N-гидроксилирование) Окисление тиоэфиров
- 32. Обезвреживание этанола Поступивший в организм этанол кровью переносится во все органы и ткани организма. Его катаболизм
- 33. Окисление этанола NAD-зависимой алкогольдегидрогеназой Алкогольдегидрогеназа катализирует обратимую реакцию, направление которой зависит от концентрации ацетальдегида и соотношения
- 34. Изоферменты алкогольдегидрогеназы. Класс I АДГ-изоферментов (α-АДГ, β- АДГ и γ - АДГ) – окисление этанола и
- 35. 2. Окисление этанола при участии цитохром Р450 - зависимой микросомальной этанолокисляющей системы Цитохром Р450-зависимая микросомальная этанолокисляющая
- 36. При хроническом алкоголизме окисление этанола ускоряется на 50 – 70 % за счёт гипертрофии ЭР и
- 37. 3. Окисление этанола каталазой Второстепенную роль в окислении этанола играет каталаза, находящаяся в пероксисомах. Этот фермент
- 38. Ацетальдегид, образовавшийся из этанола, окисляется до уксусной кислоты. Работают два фермента: 1) FAD -зависимая альдегидоксидаза: СН3СНО
- 39. 2) Ацетальдегиддегидрогеназа (АлДГ) – окисляет субстрат при участии кофермента NAD+. СН3СНО + Н2О + NAD+ →
- 40. Например, митохрондриальная изоформа АлДГ гепатоцитов, обладает более высоким сродством к ацетальдегиду (имеет низкую константу Михаэлиса КМ
- 41. Эффекты этанола в печени
- 42. Эффекты этанола в печени (пояснение схемы). 1 → 2 → 3 - окисление этанола до ацетата
- 43. Следствия химической модификации молекулы ксенобиотика
- 44. Следствия химической модификации молекулы ксенобиотика
- 45. За возможность образования легко выводимого из организма вещества путем окисления моноаминоксидазами клетка «платит» образованием в ряде
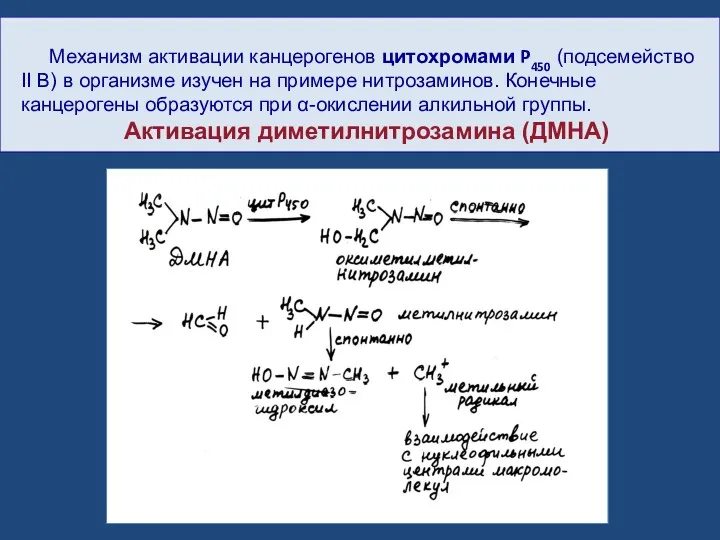
- 46. Механизм активации канцерогенов цитохромами P450 (подсемейство II В) в организме изучен на примере нитрозаминов. Конечные канцерогены
- 48. Скачать презентацию
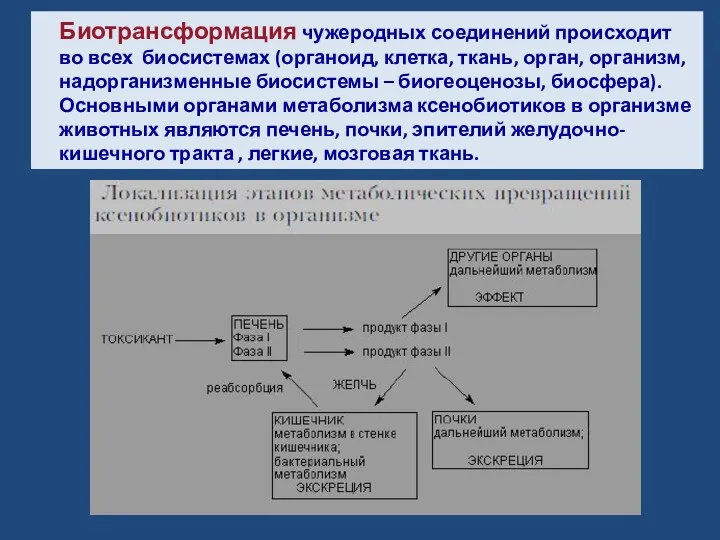
Биотрансформация чужеродных соединений происходит во всех биосистемах (органоид, клетка, ткань, орган,
Биотрансформация чужеродных соединений происходит во всех биосистемах (органоид, клетка, ткань, орган,
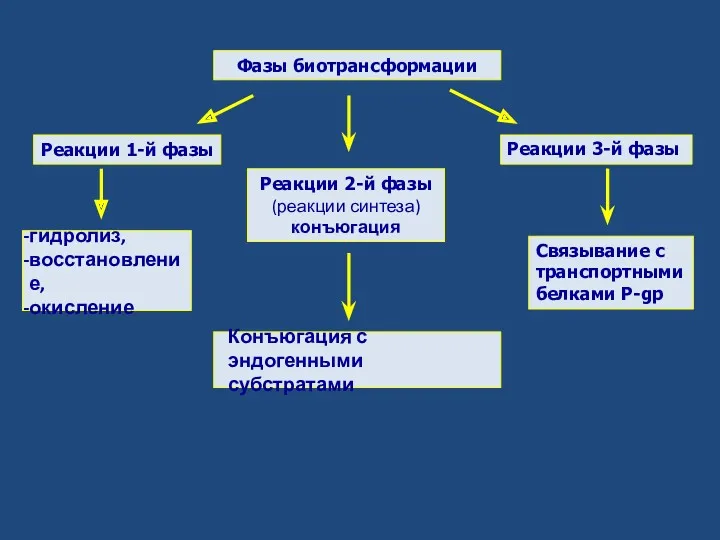
Фазы биотрансформации
Реакции 1-й фазы
Реакции 2-й фазы
(реакции синтеза)
конъюгация
гидролиз,
восстановление,
окисление
Конъюгация с эндогенными
Фазы биотрансформации
Реакции 1-й фазы
Реакции 2-й фазы
(реакции синтеза)
конъюгация
гидролиз,
восстановление,
окисление
Конъюгация с эндогенными
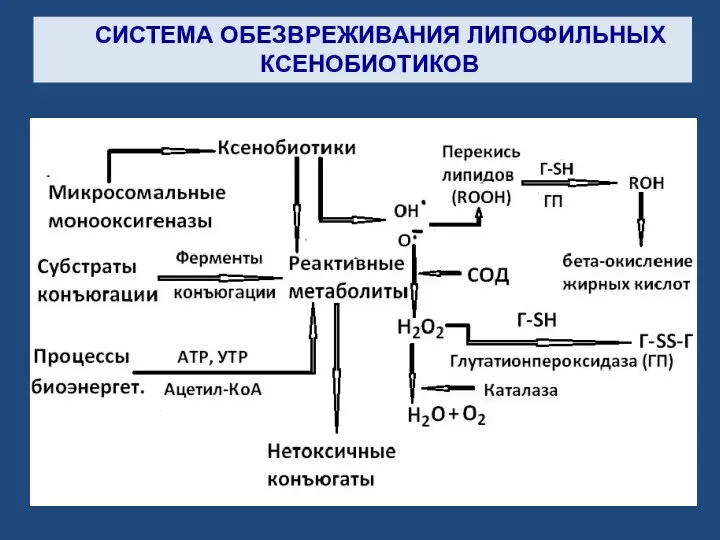
СИСТЕМА ОБЕЗВРЕЖИВАНИЯ ЛИПОФИЛЬНЫХ КСЕНОБИОТИКОВ
СИСТЕМА ОБЕЗВРЕЖИВАНИЯ ЛИПОФИЛЬНЫХ КСЕНОБИОТИКОВ

Основные пути биотрансформации чужеродных соединений.
I ФАЗА
Окисление:
а) микросомальное
I ФАЗА
Окисление:
а) микросомальное

Восстановление:
а) восстановление нитросоединений, азосоединений микросомальными ферментами;
б) микросомальное восстановительное галогенирование;
в) немикросомальное восстановление.
Гидролиз
а) восстановление нитросоединений, азосоединений микросомальными ферментами;
б) микросомальное восстановительное галогенирование;
в) немикросомальное восстановление.
Гидролиз

Ферменты биотрансформации ксенобиотиков присутствуют, в основном, в микросомах и в цитозоле,
Ферменты биотрансформации ксенобиотиков присутствуют, в основном, в микросомах и в цитозоле,
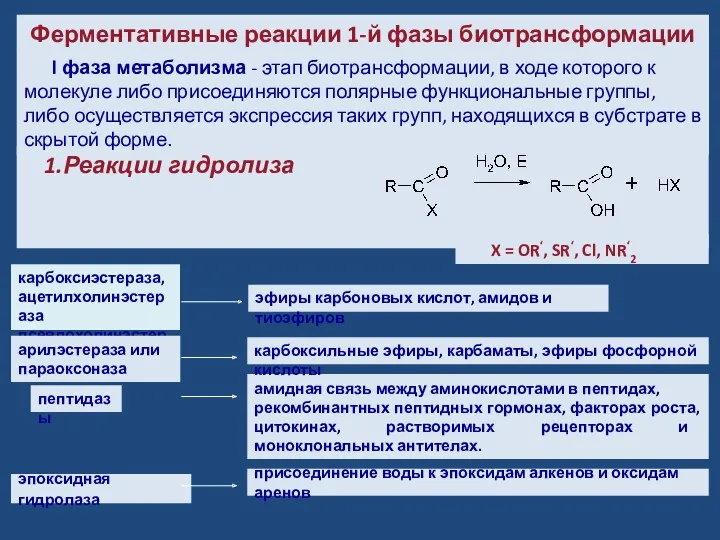
Ферментативные реакции 1-й фазы биотрансформации
карбоксиэстераза,
ацетилхолинэстераза
псевдохолинэстераза
эпоксидная гидролаза
l фаза метаболизма
Ферментативные реакции 1-й фазы биотрансформации
карбоксиэстераза,
ацетилхолинэстераза
псевдохолинэстераза
эпоксидная гидролаза
l фаза метаболизма
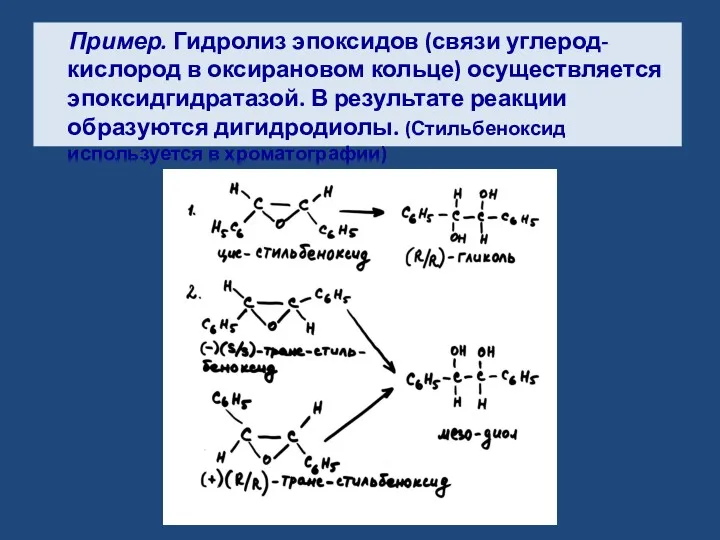
Пример. Гидролиз эпоксидов (связи углерод-кислород в оксирановом кольце) осуществляется эпоксидгидратазой. В
Пример. Гидролиз эпоксидов (связи углерод-кислород в оксирановом кольце) осуществляется эпоксидгидратазой. В

2 . Реакции восстановления.
Восстанавливаются некоторые металлы, альдегиды, кетоны, дисульфиды, сульфоксиды, хиноны,
2 . Реакции восстановления.
Восстанавливаются некоторые металлы, альдегиды, кетоны, дисульфиды, сульфоксиды, хиноны,
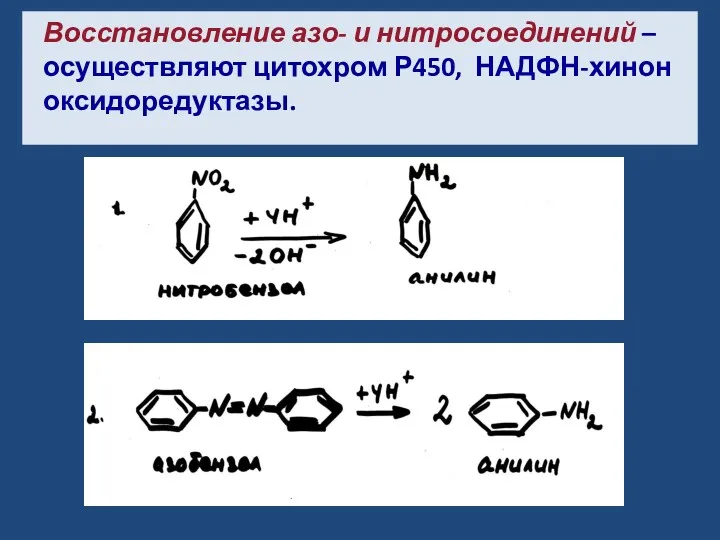
Восстановление азо- и нитросоединений – осуществляют цитохром Р450, НАДФН-хинон оксидоредуктазы.
Восстановление азо- и нитросоединений – осуществляют цитохром Р450, НАДФН-хинон оксидоредуктазы.
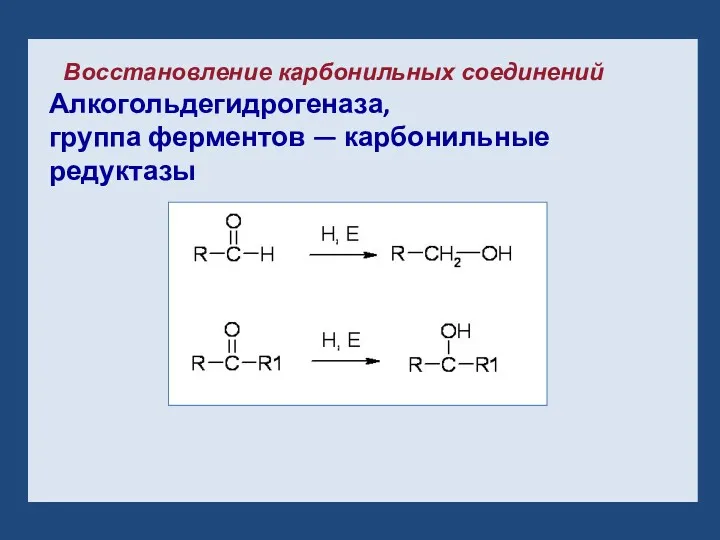
Восстановление карбонильных соединений
Алкогольдегидрогеназа,
группа ферментов — карбонильные редуктазы
Алкогольдегидрогеназа,
группа ферментов — карбонильные редуктазы

Восстановление дисульфидов – ферменты:
глутатионредуктаза,
глутатион - S-трансфераза, или
неферментативно
Восстановление дисульфидов – ферменты:
глутатионредуктаза,
глутатион - S-трансфераза, или
неферментативно
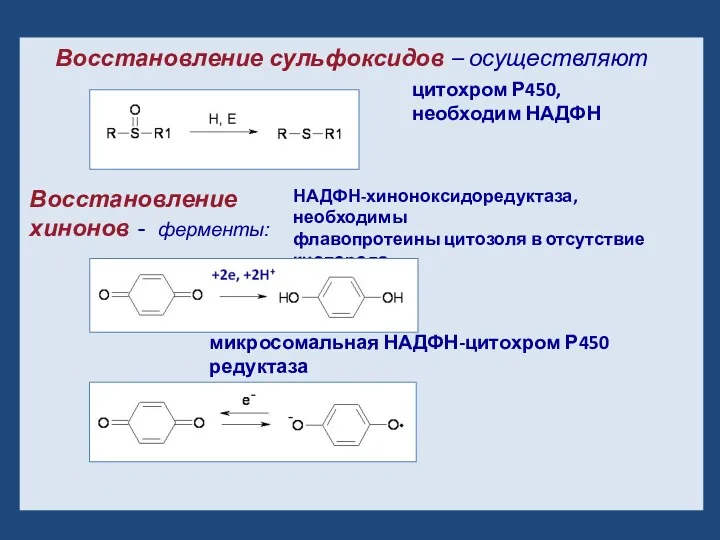
Восстановление сульфоксидов – осуществляют
цитохром Р450, необходим НАДФН
Восстановление хинонов - ферменты:
НАДФН-хиноноксидоредуктаза,
Восстановление сульфоксидов – осуществляют
цитохром Р450, необходим НАДФН
Восстановление хинонов - ферменты:
НАДФН-хиноноксидоредуктаза,
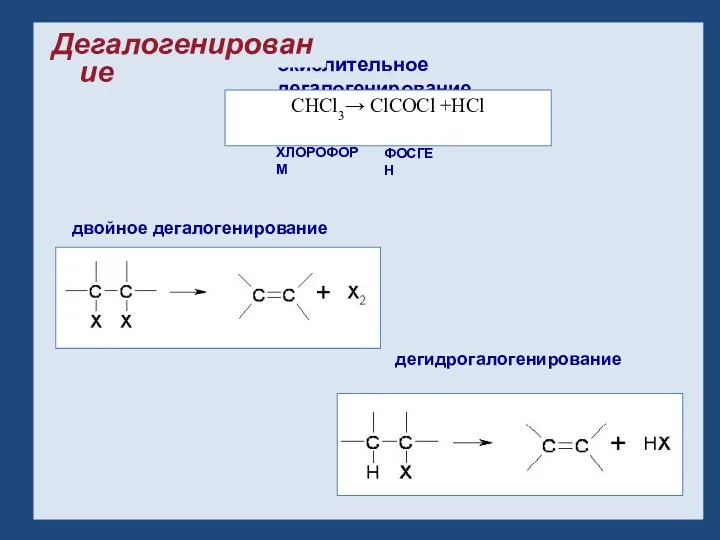
Дегалогенирование:
окислительное дегалогенирование
двойное дегалогенирование
дегидрогалогенирование
ХЛОРОФОРМ
ФОСГЕН
CHCl3→ ClCOCl +HCl
Дегалогенирование
Дегалогенирование:
окислительное дегалогенирование
двойное дегалогенирование
дегидрогалогенирование
ХЛОРОФОРМ
ФОСГЕН
CHCl3→ ClCOCl +HCl
Дегалогенирование
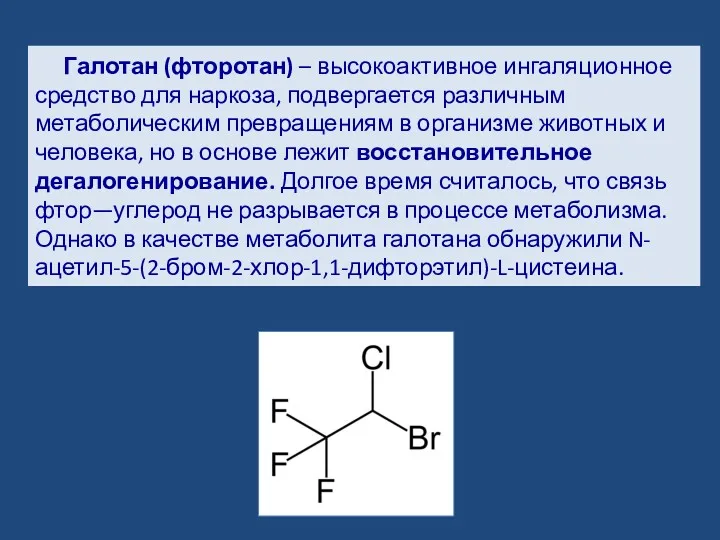
Галотан (фторотан) – высокоактивное ингаляционное средство для наркоза, подвергается различным метаболическим
Галотан (фторотан) – высокоактивное ингаляционное средство для наркоза, подвергается различным метаболическим

Последние данные указывают, что метаболизм галотана условно делится на два пути:
Последние данные указывают, что метаболизм галотана условно делится на два пути:

3. Реакции окисления.
Альдегиддегидрогеназы (АлДГ) - окисление альдегидов до карбоновых кислот (кофактор
Альдегиддегидрогеназы (АлДГ) - окисление альдегидов до карбоновых кислот (кофактор
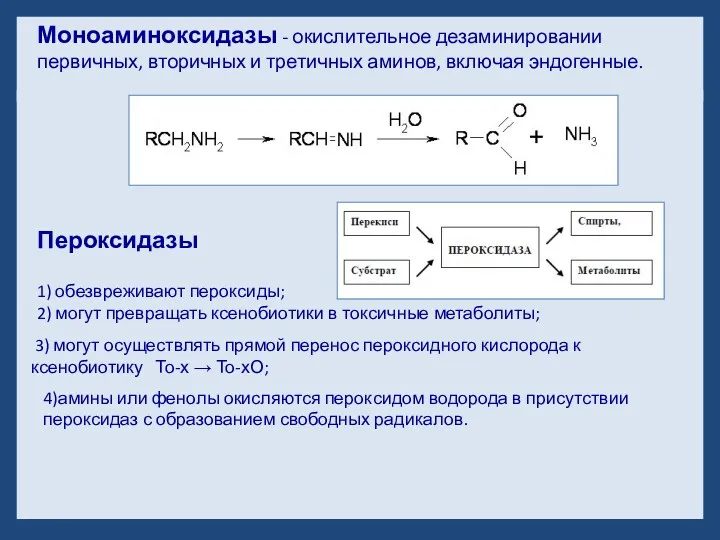
Пероксидазы
1) обезвреживают пероксиды;
2) могут превращать ксенобиотики в токсичные метаболиты;
Пероксидазы
1) обезвреживают пероксиды;
2) могут превращать ксенобиотики в токсичные метаболиты;
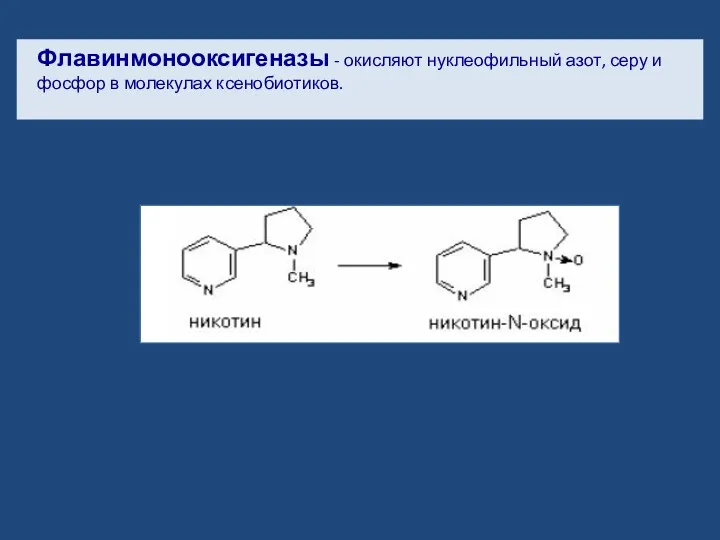
Флавинмонооксигеназы - окисляют нуклеофильный азот, серу и фосфор в молекулах ксенобиотиков.
Флавинмонооксигеназы - окисляют нуклеофильный азот, серу и фосфор в молекулах ксенобиотиков.

Цитохром Р450 катализирует реакции окисления:
-гидроксилирование алифатических и ароматических углеводородов;
эпоксидирование двойной
-гидроксилирование алифатических и ароматических углеводородов;
эпоксидирование двойной
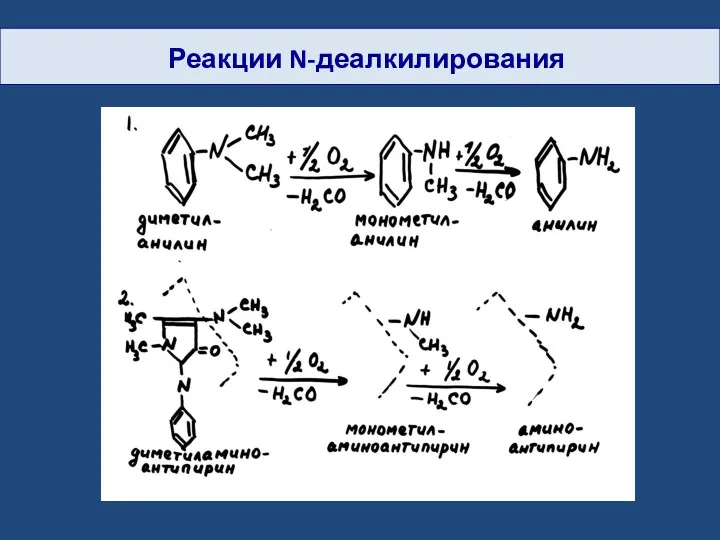
Реакции N-деалкилирования
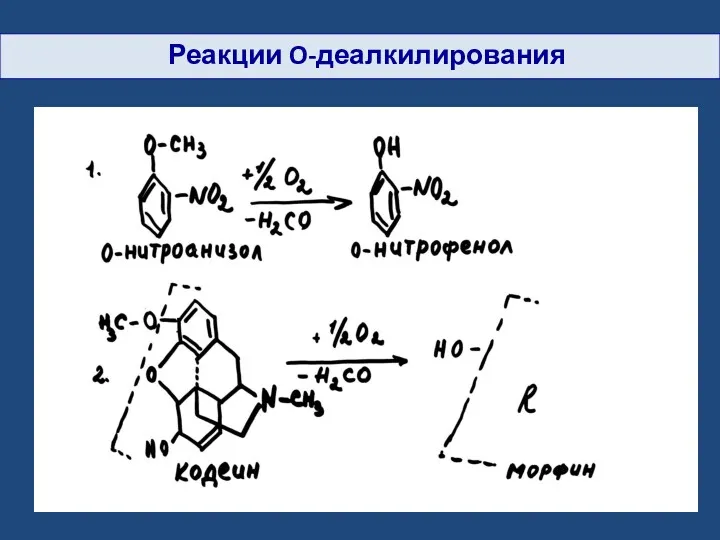
Реакции O-деалкилирования
Реакции O-деалкилирования
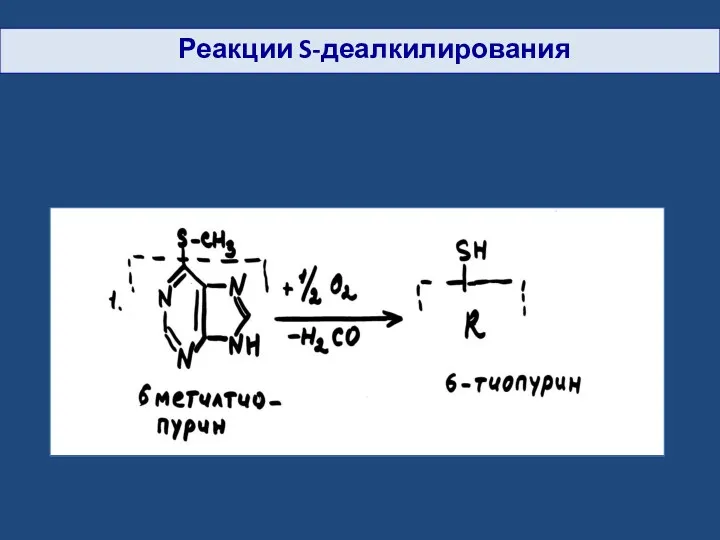
Реакции S-деалкилирования
Реакции S-деалкилирования
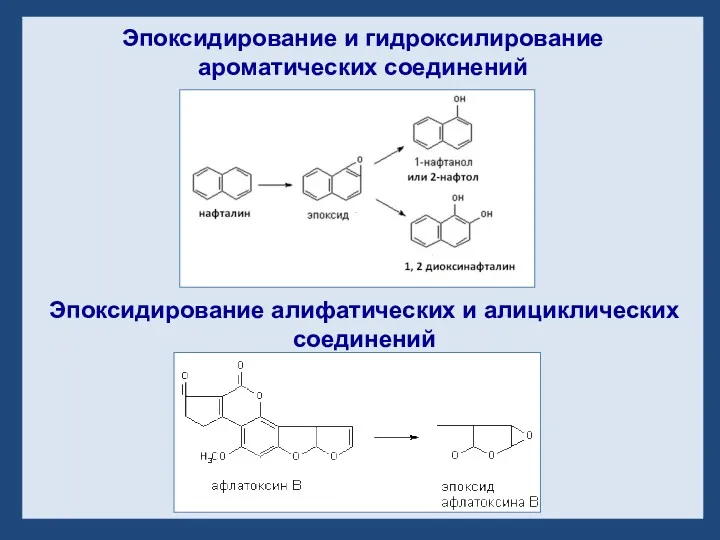
Эпоксидирование и гидроксилирование ароматических соединений
Эпоксидирование алифатических и алициклических соединений
Эпоксидирование и гидроксилирование ароматических соединений
Эпоксидирование алифатических и алициклических соединений
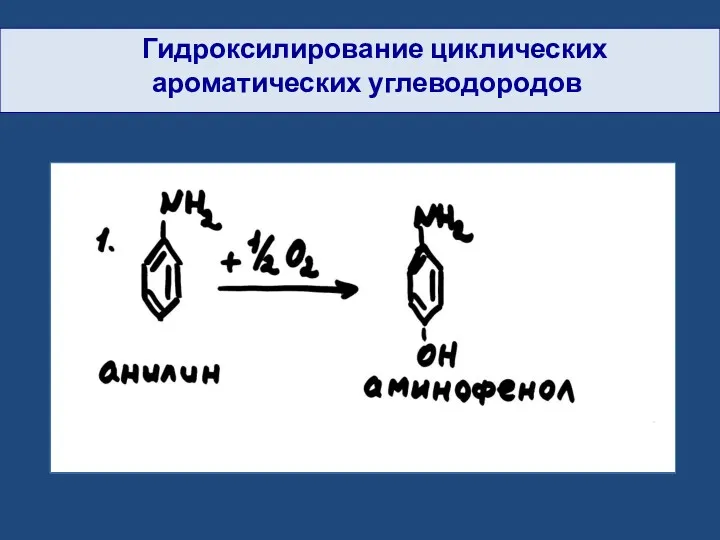
Гидроксилирование циклических ароматических углеводородов
Гидроксилирование циклических ароматических углеводородов
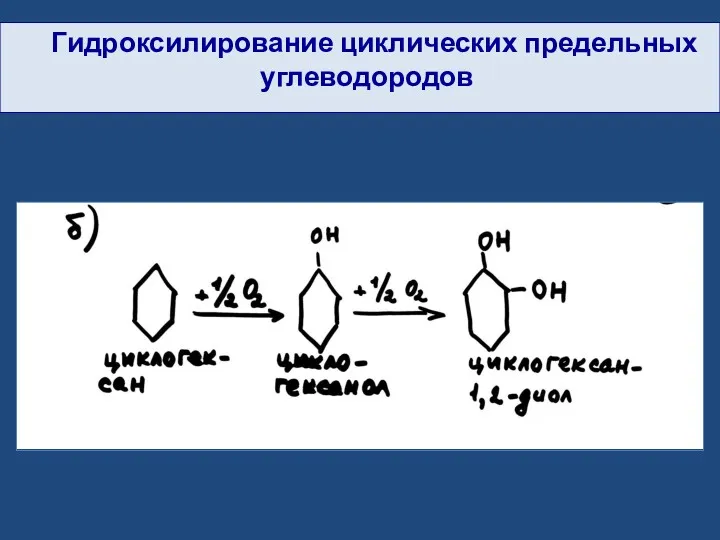
Гидроксилирование циклических предельных углеводородов
Гидроксилирование циклических предельных углеводородов
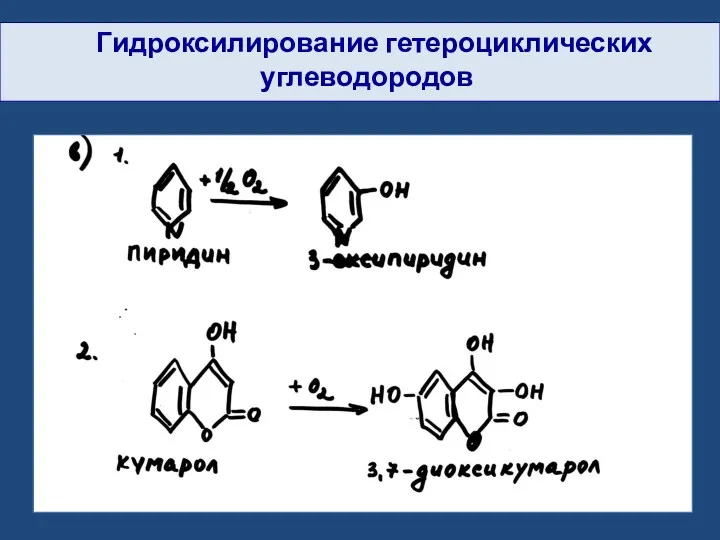
Гидроксилирование гетероциклических углеводородов
Гидроксилирование гетероциклических углеводородов

Гидроксилирование алифатических соединений:
а) предельных углеводородов
Гидроксилирование алифатических соединений:
а) предельных углеводородов
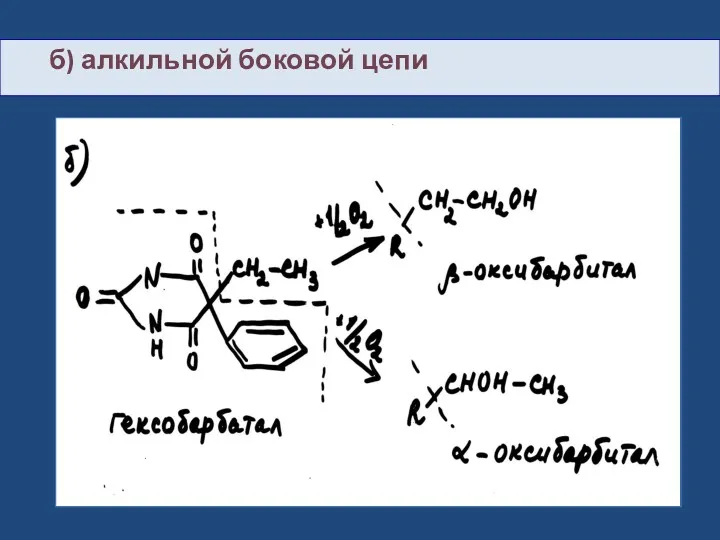
б) алкильной боковой цепи
б) алкильной боковой цепи
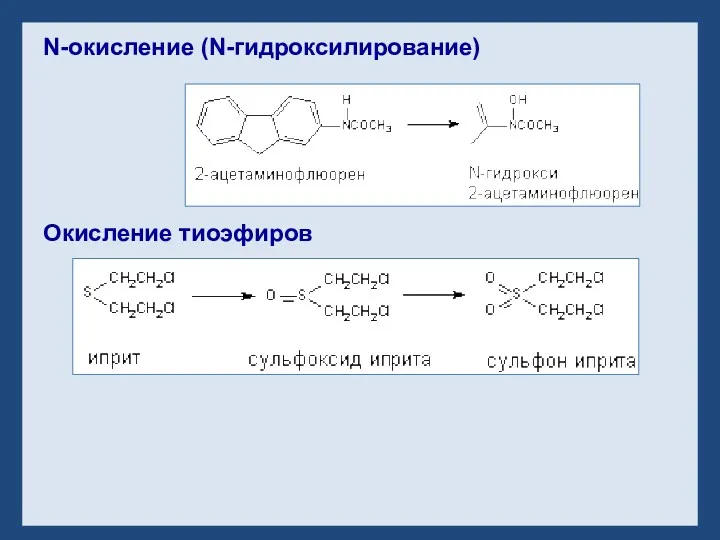
N-окисление (N-гидроксилирование)
Окисление тиоэфиров
N-окисление (N-гидроксилирование)
Окисление тиоэфиров

Обезвреживание этанола
Поступивший в организм этанол кровью переносится во все органы и
Поступивший в организм этанол кровью переносится во все органы и

Окисление этанола NAD-зависимой алкогольдегидрогеназой
Алкогольдегидрогеназа катализирует обратимую реакцию, направление которой зависит от
Алкогольдегидрогеназа катализирует обратимую реакцию, направление которой зависит от

Изоферменты алкогольдегидрогеназы.
Класс I АДГ-изоферментов (α-АДГ, β- АДГ и γ - АДГ)
Класс I АДГ-изоферментов (α-АДГ, β- АДГ и γ - АДГ)

2. Окисление этанола при участии цитохром Р450 - зависимой микросомальной этанолокисляющей
2. Окисление этанола при участии цитохром Р450 - зависимой микросомальной этанолокисляющей

При хроническом алкоголизме окисление этанола ускоряется на 50 – 70 %
При хроническом алкоголизме окисление этанола ускоряется на 50 – 70 %

3. Окисление этанола каталазой
Второстепенную роль в окислении этанола играет каталаза,
Второстепенную роль в окислении этанола играет каталаза,

Ацетальдегид, образовавшийся из этанола, окисляется до уксусной кислоты. Работают два фермента:

2) Ацетальдегиддегидрогеназа (АлДГ) – окисляет субстрат при участии кофермента NAD+.
СН3СНО
СН3СНО

Например, митохрондриальная изоформа АлДГ гепатоцитов, обладает более высоким сродством к ацетальдегиду
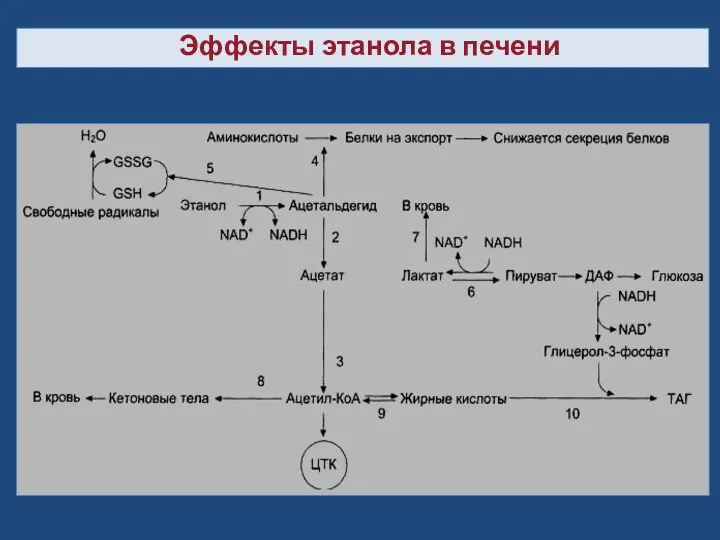
Эффекты этанола в печени
Эффекты этанола в печени

Эффекты этанола в печени (пояснение схемы).
1 → 2 → 3
1 → 2 → 3

Следствия химической модификации молекулы ксенобиотика
Следствия химической модификации молекулы ксенобиотика

Следствия химической модификации молекулы ксенобиотика
Следствия химической модификации молекулы ксенобиотика

 Металлы, их свойства и область применения
Металлы, их свойства и область применения Галогены - химические элементы 17-й группы периодической таблицы химических элементов Д.И. Менделеева
Галогены - химические элементы 17-й группы периодической таблицы химических элементов Д.И. Менделеева Тяжелые металлы
Тяжелые металлы Гидрокси(окси) кислоты
Гидрокси(окси) кислоты Значение пищи и ее состав
Значение пищи и ее состав Коррозия металлов
Коррозия металлов Изучение процесса коррозии железа (домашний эксперимент)
Изучение процесса коррозии железа (домашний эксперимент) Борьба с биологической коррозией
Борьба с биологической коррозией Реакции ионного обмена
Реакции ионного обмена Спирты
Спирты Газовые смеси
Газовые смеси Химический элемент, минерал, горная порода. (Лекция 6)
Химический элемент, минерал, горная порода. (Лекция 6) Титриметрический анализ. Кривые титрования
Титриметрический анализ. Кривые титрования Адсорбция
Адсорбция Ендотермічні реакції на службі людини
Ендотермічні реакції на службі людини Поверхностные явления. Адсорбция
Поверхностные явления. Адсорбция Химические реакции или химические явления
Химические реакции или химические явления Спирты. Понятия о предельных одноатомных спиртах. Химические свойства этанола и его применение
Спирты. Понятия о предельных одноатомных спиртах. Химические свойства этанола и его применение Хром, марганец
Хром, марганец Химическая кинетика и катализ
Химическая кинетика и катализ Свойства фосфора
Свойства фосфора Общие сведения о полезных ископаемых. (Лекция 2)
Общие сведения о полезных ископаемых. (Лекция 2) Тұндыру әдістері
Тұндыру әдістері Термодинамика химических процессов
Термодинамика химических процессов Чистые вещества и смеси. Химия. 8 кл
Чистые вещества и смеси. Химия. 8 кл Обед под микроскопом. Минеральные вещества
Обед под микроскопом. Минеральные вещества Типы химических реакций на примере свойств воды
Типы химических реакций на примере свойств воды Реакции солей в растворе. Гидролиз солей. Водородный показатель рН
Реакции солей в растворе. Гидролиз солей. Водородный показатель рН