- Механизмы трансформации энергии в фотосинтезе

Содержание
- 2. ФОТОАКТИВАЦИЯ Mn/Ca кластер можно экстрагировать из ФС2 без инактивации связывающих участков. Это доказывается тем, что каталитический
- 3. Схема самосборки марганцевого кластера в процессе ФА Mn2+ Mn2+ Mn2+ Mn2+ Ca2+ Высокоаффинный Mn-связывающий участок Генерируемый
- 4. МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ Yachandra et al., 1996 Увеличение расстояния 2,7А, зафиксированное EXAFS.
- 5. МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ Hillier & Wydrzynski, 2000
- 6. МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ Brudvig G with coworkers, 2005
- 7. МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ Barber & Tran, 2013 На рисунке представлены два возможных механизма финальной стадии S
- 8. (Suga et al. 2014) При переходе из S1 в S2 состояние может окисляться либо марганец Mn4
- 12. Каталитический центр окисления воды расположен в глубине белковой структуры, поэтому должны существовать каналы для доставки субстрата
- 13. Пути транспорта молекул воды, протонов и кислорода между КВК и внутритилакоидным пространством (компьютерное моделирование) Murray&Barber, 2007
- 14. Каналы должен иметь такой минимальный диаметр, чтобы обеспечить прохождение кислорода или воды. Канал i – наиболее
- 15. Канал iii – наиболее полярный канал и содержит ряд аминокислот, характерных для протонных каналов. Поэтому, предполагается,
- 16. КАНАЛЫ Najafpour et al., 2012 Кислород Вода Н+
- 17. Синтез АТФ Аденозинтрифосфат Cинтез АТФ осуществляется мембранным ферментом АТФ-синтазой, использующим энергию протонов, перемещающихся по протонному градиенту
- 18. Свободная энергия. Преобразование энергии в живых системах. Высокоэнергетические фосфаты. Свободная энергия G – наиболее полезная термодинамическая
- 19. В клетках, освобождаемая при окислении энергия, преобразуется путем сопряжения процесса окисления с синтезом АТФ из АДФ
- 20. Пример механизма использования АТФ: синтез сложного эфира. RCOOH + HO-R’ ⇔ RCOOR’ + H2O ΔG0 =
- 21. Структура АТФ-синтазы Строма Люмен
- 22. Водорастворимый фактор сопряжения F1 cостоит из: 3 субъединиц α неподвижная часть (статор) 3 субъединиц β неподвижная
- 23. Функционирование АТФ синтазы Связывание субстрата, синтез АТФ и высвобождение АТФ. Каждая β субъединица фактора F1 имеет
- 24. α АТФ γ γ Р Р Скорость вращения ротора – до 700 оборотов в сек по
- 25. Функционирование АТФ синтазы Механизм вращения ротора. Как осуществляется вращение γ-субъединицы? γ-субъединица, участвующая синтезе АТФ, входит в
- 26. 5. Каналы расположены в гидрофобной неподвижной субъединице а, примыкающей к кольцу ротора. Кольцо ротора. 1. Кольцо
- 27. 5. После поворота кольца ротора на 360о остаток Asp депротонируется и протон выбрасывается наружу по выходному
- 28. Протонирование/депротонирование c-Asp кислоты сопровождается поворотом ТМC с субъединицы, который обеспечивается взаимодействием с остатком Arg субъединицы а.
- 29. In the starting position, the stator arginine is tightly bound to binding site S1. An incoming
- 30. PQ PQ FNR или FQR (?) Циклический транспорт электронов NDH – NAD(P)H dehydrogenase FNR - ferredoxin-NADP
- 31. Finazzi et al. 2003 Циклический транспорт электронов
- 32. Allen, 2003
- 33. Методы исследования фотосистемы 2. Флуоресценция и люминесценция. Флуоресценция в ФС2 является результатом главным образом дезактивации возбужденной
- 34. Методы исследования фотосистемы 2. Флуоресценция и люминесценция. Люминесценция – регистрация квантов света, появляющихся при рекомбинации зарядов
- 35. Yzox DCMU P680+ Mn(II) P680* Pheo Q-a Qb P680 Скорость уменьшения выхода флуоресценции зависит от скорости
- 36. Кинетика уменьшения выхода флуоресценции, индуцированной вспышкой света: Qa- → Qb t1/2 = 100 – 250 мкс
- 37. Методы исследования фотосистемы 2. Кинетика индукции флуоресценции Strasser, 1997
- 38. Методы исследования фотосистемы 2. Замедленная флуоресценция Guo & Tan, 2013
- 39. Методы исследования фотосистемы 2. ЭПР ЭПР – практически все электрон-транспортные компоненты ФС2 дают сигнал ЭПР, но
- 40. Полярография – метод измерения концентрации кислорода в среде (соответственно, возможность измерения скорости окисления воды фотосистемой 2
- 41. Эволюция фотосистем. ФС1: центральные белки PsaA и PsaB имеют по 11 трансмембранных спиралей и каждый белок
- 42. Рентгеновский анализ кристаллов ФС1 и ФС2 показал очень интересные результаты: расположение в мембране спиралей белков D1
- 43. Расположение трансмембранных спиралей ФС1 и ФС2.
- 44. Эволюция фотосистем. Структура белка ФС1 ФС2 СР43/CP47 D1/D2 PsaA/PsaB
- 45. Большое сходство в структурной организации фотосистем 1-го и 2-го типов позволило ряду авторов предположить, что эти
- 46. Эволюция. Происхождение фотосистем. Редокс переключатель (Allen, 2005) Fig. 2. Retention of type I and type II
- 48. Эволюция ФС2 Анаэробные зеленые бактерии Allen & Martin, 2007 В цианобактериях Oscillatoria limnetica в присутствии H2S
- 49. Эволюция ФС2 4500 500 АРХЕЙ ПРОТЕРОЗОЙ ПАЛЕОЗОЙ ⇒ 2500 Млн. лет О2 Fe2+ 0.01 атм 10-20M
- 50. Возможные доноры электронов до использования воды: Fe(II) H2S H2O2 – легко окисляемый донор, для реакции окисления
- 51. СИММЕТРИЯ МЕЖДУ СООН КОНЦЕВЫМИ УЧАСТКАМИ ПОЛИПЕПТИДОВ D1 ИD2 D1 …. SVVDSQGRVINTWAD II NRANLGMEVMHERNAHNFDLDLA D2 …PEFETFYTKNIIINEGIRAWMAAQDQPHENLIFPEEVLPRG 344
- 52. Примеры использования фотосистем в биотехнологии Глобальное потребление энергии в настоящее время около 17 TW/год. Около 40%
- 53. Примеры использования фотосистем в биотехнологии Фотоводород. Фотосинтетические бактерии, цианобактерии и одноклеточные зеленые водоросли могут на свету
- 54. Механизм выделения водорода в одноклеточных зеленых водорослях Гидрогеназа
- 55. Источником электронов для гидрогеназы является ферредоксин, восстанавливаемый ФС1. В свою очередь, ФС1 восстанавливается через цепь переносчиков
- 56. 2. Биосенсоры. Для обнаружения гербицидов и тяжелых металлов в воде, почве, продуктах. Giardi & Pace, 2005
- 57. 3. Био-фотосенсор. Фотосистема 1 из термоустойчивых цианобактерий соединена молекулярной “проволочкой” с транзистором. (Terasaki et al. 2007).
- 58. Фотосистема 1 соединена молекулярной “проволочкой” с гидрогеназой (Lubner et al., 2010).
- 60. Скачать презентацию

ФОТОАКТИВАЦИЯ
Mn/Ca кластер можно экстрагировать из ФС2 без инактивации связывающих участков.
Это доказывается
ФОТОАКТИВАЦИЯ
Mn/Ca кластер можно экстрагировать из ФС2 без инактивации связывающих участков.
Это доказывается
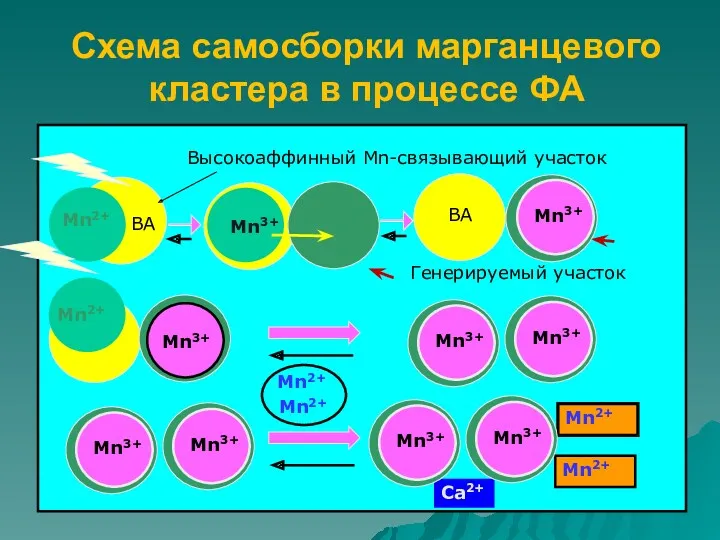
Схема самосборки марганцевого кластера в процессе ФА
Mn2+
Mn2+
Mn2+
Mn2+
Ca2+
Высокоаффинный Mn-связывающий участок
Генерируемый участок
ВА
ВА
Mn3+
Схема самосборки марганцевого кластера в процессе ФА
Mn2+
Mn2+
Mn2+
Mn2+
Ca2+
Высокоаффинный Mn-связывающий участок
Генерируемый участок
ВА
ВА
Mn3+
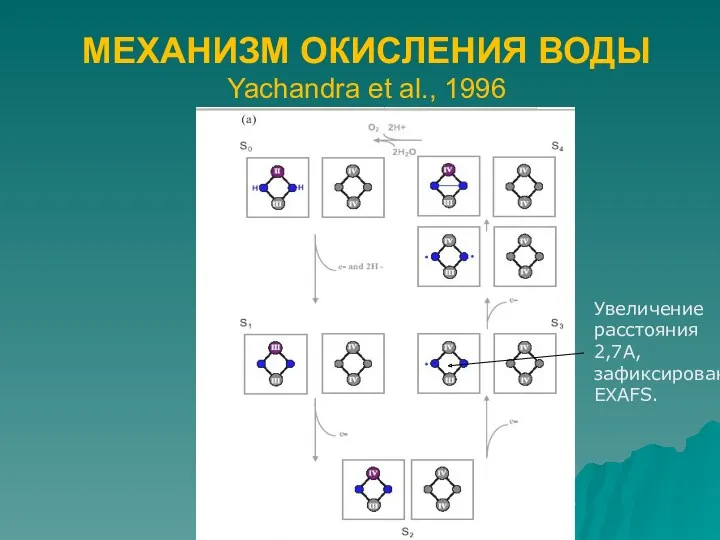
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Yachandra et al., 1996
Увеличение
расстояния
2,7А,
зафиксированное
EXAFS.
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Yachandra et al., 1996
Увеличение
расстояния
2,7А,
зафиксированное
EXAFS.
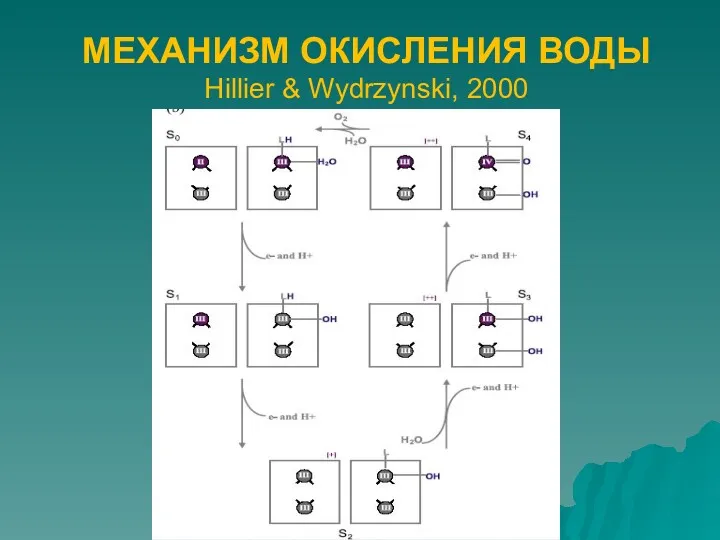
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Hillier & Wydrzynski, 2000
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Hillier & Wydrzynski, 2000
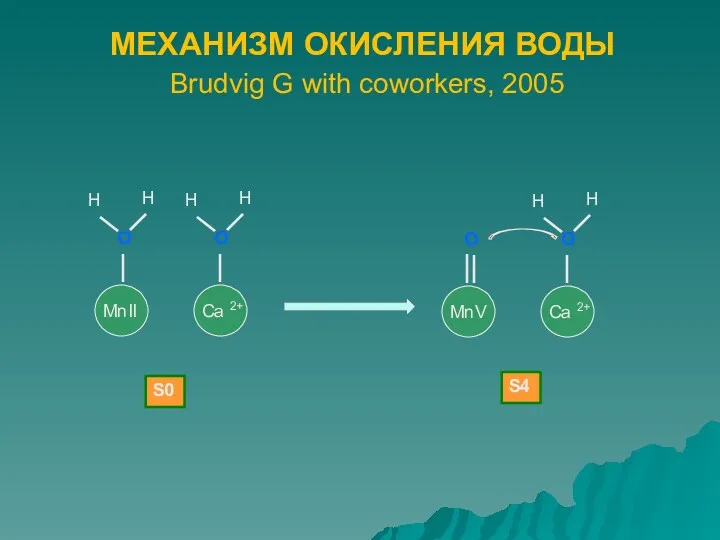
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Brudvig G with coworkers, 2005
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Brudvig G with coworkers, 2005
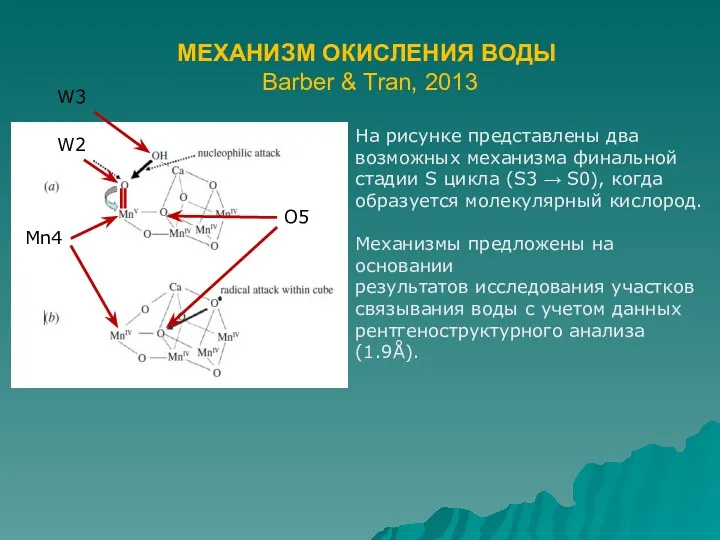
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Barber & Tran, 2013
На рисунке представлены два
возможных
МЕХАНИЗМ ОКИСЛЕНИЯ ВОДЫ
Barber & Tran, 2013
На рисунке представлены два
возможных
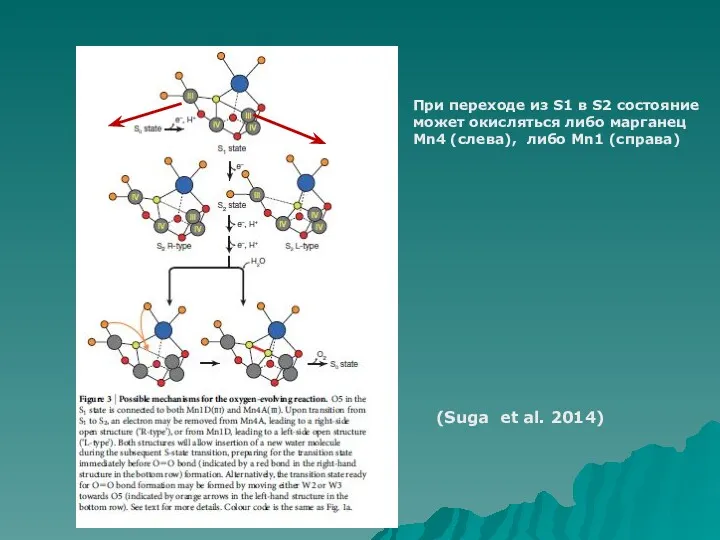
(Suga et al. 2014)
При переходе из S1 в S2 состояние
может окисляться
(Suga et al. 2014)
При переходе из S1 в S2 состояние
может окисляться
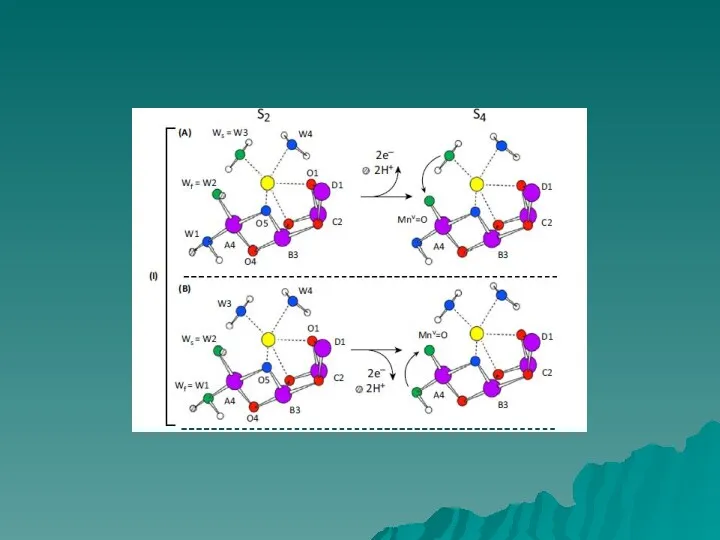
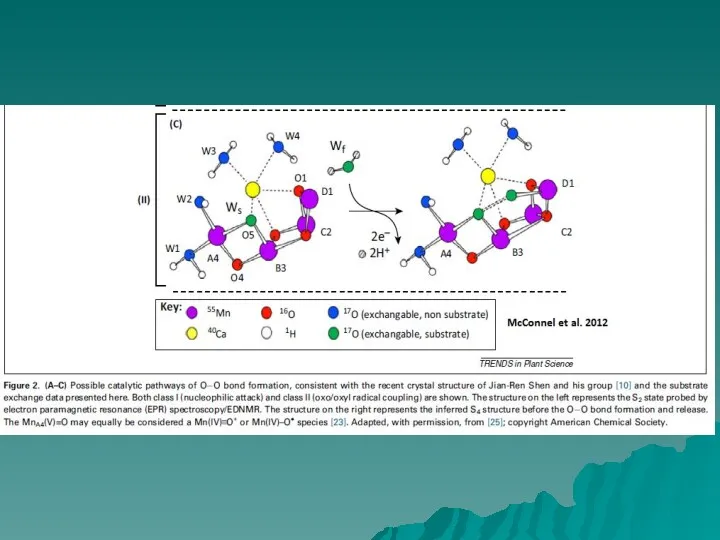
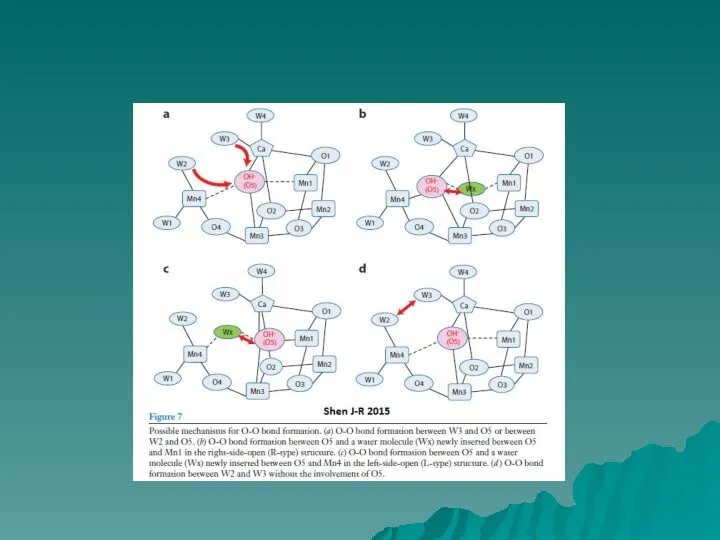

Каталитический центр окисления воды расположен в глубине белковой структуры, поэтому должны
Каталитический центр окисления воды расположен в глубине белковой структуры, поэтому должны
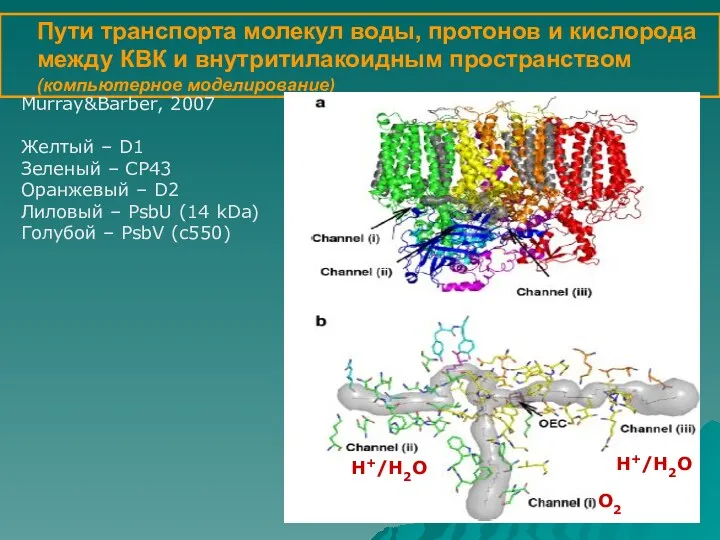
Пути транспорта молекул воды, протонов и кислорода между КВК и внутритилакоидным
Пути транспорта молекул воды, протонов и кислорода между КВК и внутритилакоидным

Каналы должен иметь такой минимальный диаметр, чтобы обеспечить прохождение кислорода или
Каналы должен иметь такой минимальный диаметр, чтобы обеспечить прохождение кислорода или

Канал iii – наиболее полярный канал и содержит ряд аминокислот, характерных
Канал iii – наиболее полярный канал и содержит ряд аминокислот, характерных
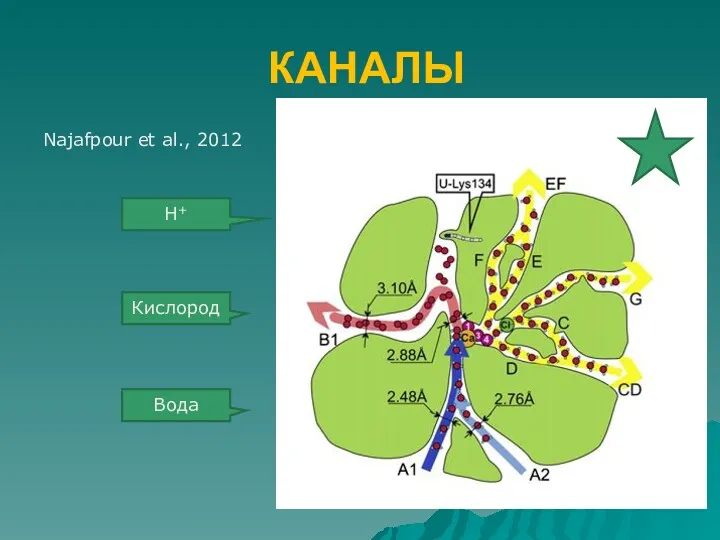
КАНАЛЫ
Najafpour et al., 2012
Кислород
Вода
Н+
КАНАЛЫ
Najafpour et al., 2012
Кислород
Вода
Н+
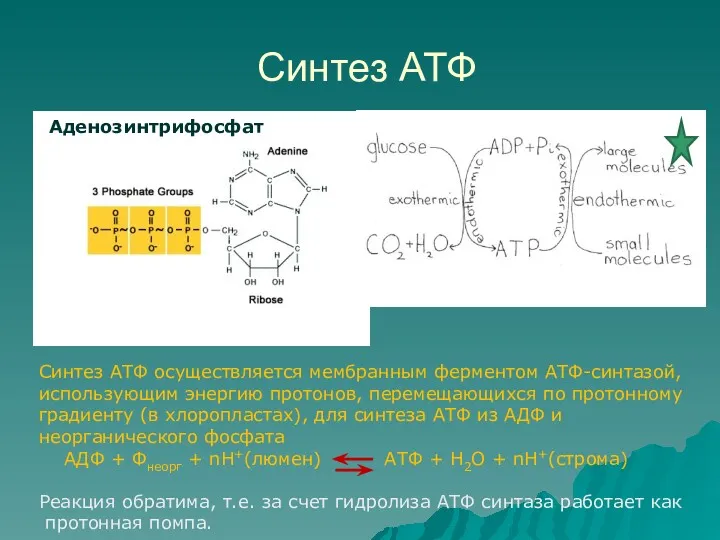
Синтез АТФ
Аденозинтрифосфат
Cинтез АТФ осуществляется мембранным ферментом АТФ-синтазой,
использующим энергию протонов, перемещающихся по
Синтез АТФ
Аденозинтрифосфат
Cинтез АТФ осуществляется мембранным ферментом АТФ-синтазой,
использующим энергию протонов, перемещающихся по

Свободная энергия. Преобразование энергии в живых системах. Высокоэнергетические фосфаты.
Свободная энергия G
Свободная энергия. Преобразование энергии в живых системах. Высокоэнергетические фосфаты.
Свободная энергия G

В клетках, освобождаемая при окислении энергия, преобразуется путем сопряжения процесса окисления
В клетках, освобождаемая при окислении энергия, преобразуется путем сопряжения процесса окисления

Пример механизма использования АТФ: синтез сложного эфира.
RCOOH + HO-R’ ⇔ RCOOR’
Пример механизма использования АТФ: синтез сложного эфира.
RCOOH + HO-R’ ⇔ RCOOR’
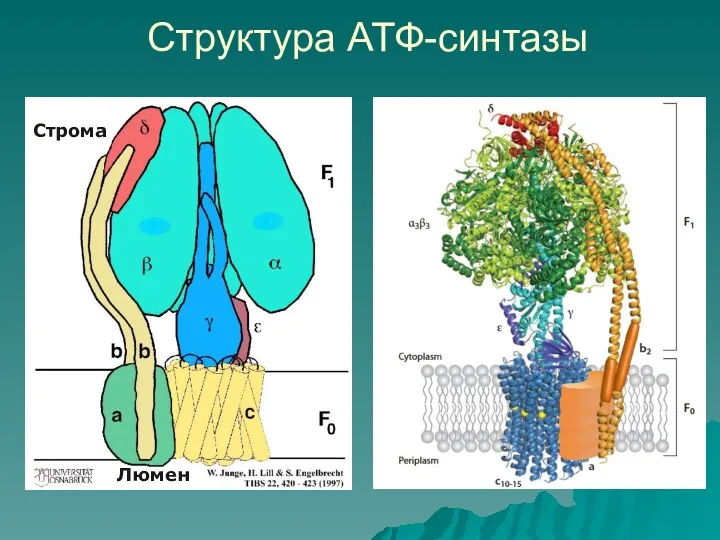
Структура АТФ-синтазы
Строма
Люмен
Структура АТФ-синтазы
Строма
Люмен

Водорастворимый фактор сопряжения F1 cостоит из:
3 субъединиц α неподвижная часть
Водорастворимый фактор сопряжения F1 cостоит из:
3 субъединиц α неподвижная часть

Функционирование АТФ синтазы
Связывание субстрата, синтез АТФ и высвобождение АТФ.
Каждая β субъединица
Функционирование АТФ синтазы
Связывание субстрата, синтез АТФ и высвобождение АТФ.
Каждая β субъединица
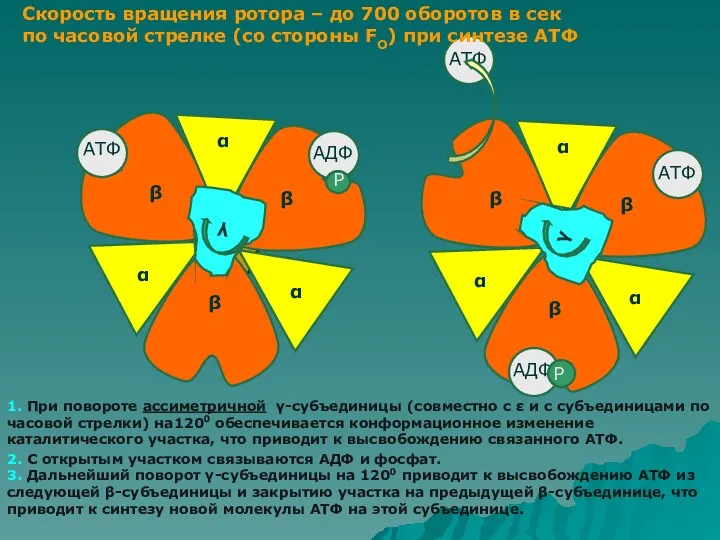
α
АТФ
γ
γ
Р
Р
Скорость вращения ротора – до 700 оборотов в сек
по часовой стрелке
α
АТФ
γ
γ
Р
Р
Скорость вращения ротора – до 700 оборотов в сек
по часовой стрелке
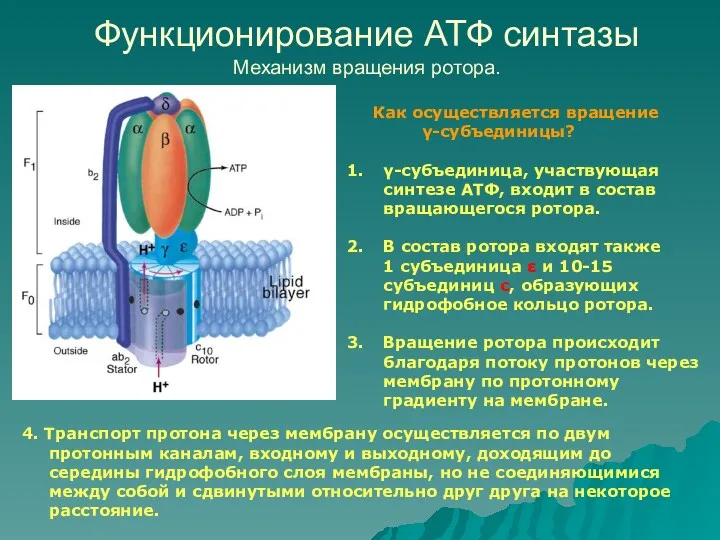
Функционирование АТФ синтазы
Механизм вращения ротора.
Как осуществляется вращение
γ-субъединицы?
γ-субъединица, участвующая
синтезе
Функционирование АТФ синтазы
Механизм вращения ротора.
Как осуществляется вращение
γ-субъединицы?
γ-субъединица, участвующая
синтезе
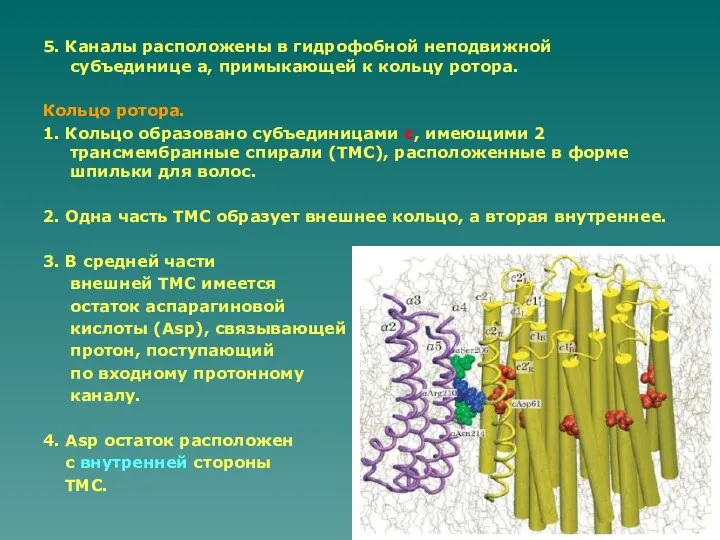
5. Каналы расположены в гидрофобной неподвижной субъединице а, примыкающей к кольцу
5. Каналы расположены в гидрофобной неподвижной субъединице а, примыкающей к кольцу

5. После поворота кольца ротора на 360о остаток Asp депротонируется и

Протонирование/депротонирование c-Asp кислоты
сопровождается поворотом ТМC с субъединицы, который обеспечивается
Протонирование/депротонирование c-Asp кислоты
сопровождается поворотом ТМC с субъединицы, который обеспечивается

In the starting position, the stator arginine is tightly bound to
In the starting position, the stator arginine is tightly bound to
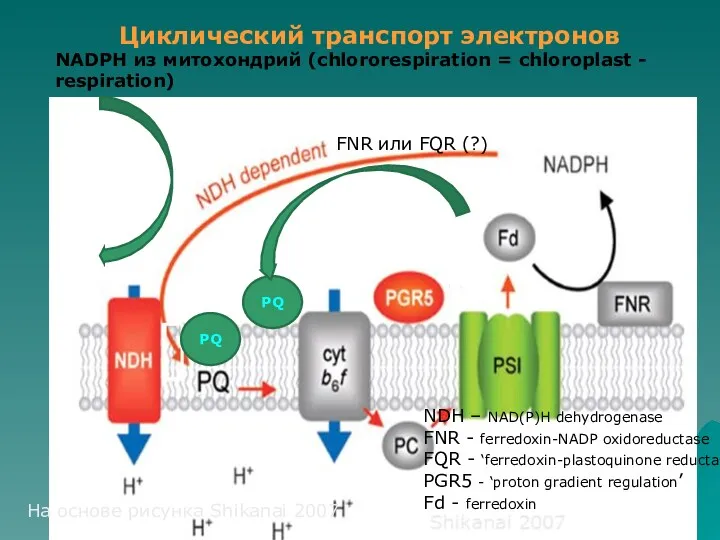
PQ
PQ
FNR или FQR (?)
Циклический транспорт электронов
NDH – NAD(P)H dehydrogenase
FNR - ferredoxin-NADP
PQ
PQ
FNR или FQR (?)
Циклический транспорт электронов
NDH – NAD(P)H dehydrogenase
FNR - ferredoxin-NADP
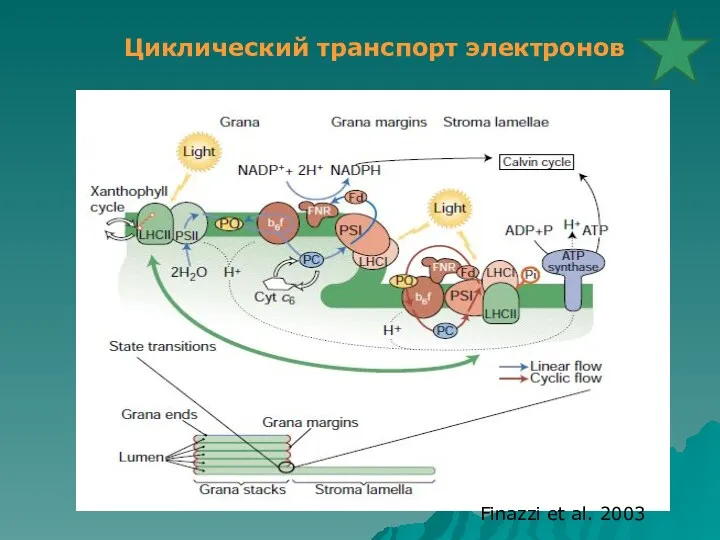
Finazzi et al. 2003
Циклический транспорт электронов
Finazzi et al. 2003
Циклический транспорт электронов
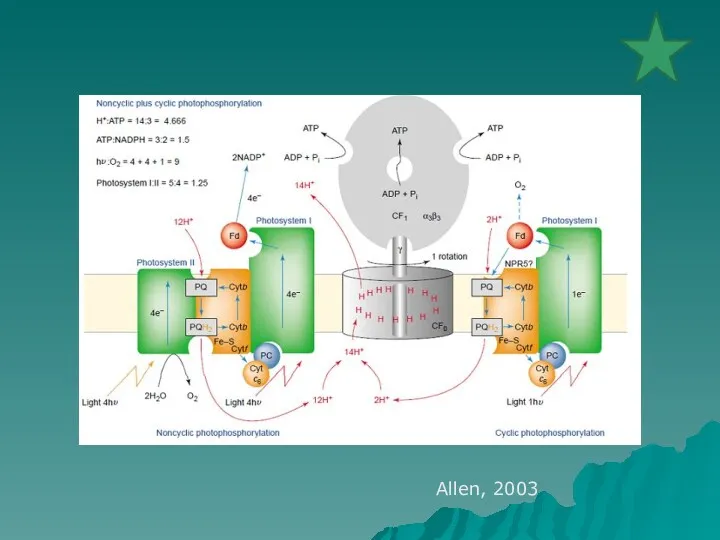
Allen, 2003
Allen, 2003

Методы исследования фотосистемы 2. Флуоресценция и люминесценция.
Флуоресценция в ФС2 является результатом
Методы исследования фотосистемы 2. Флуоресценция и люминесценция.
Флуоресценция в ФС2 является результатом
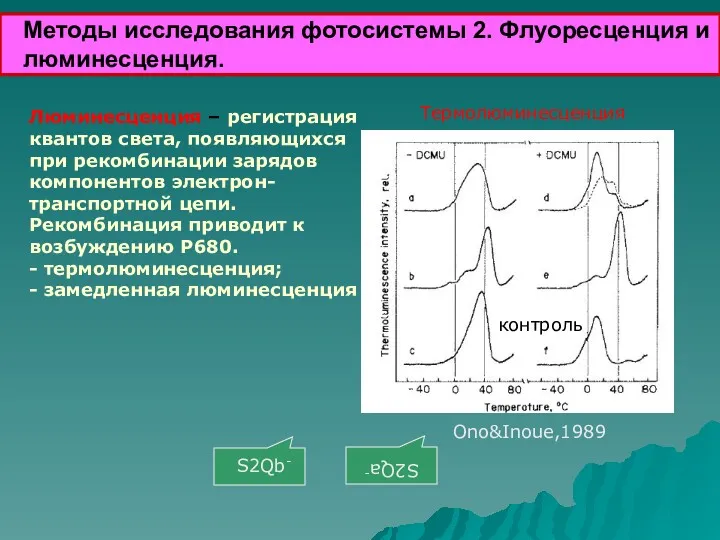
Методы исследования фотосистемы 2. Флуоресценция и люминесценция.
Люминесценция – регистрация квантов света,
Методы исследования фотосистемы 2. Флуоресценция и люминесценция.
Люминесценция – регистрация квантов света,
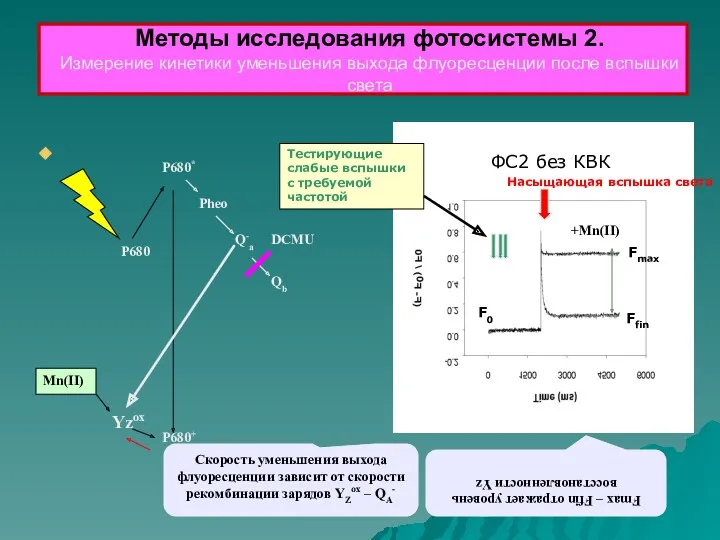
Yzox
DCMU
P680+
Mn(II)
P680*
Pheo
Q-a
Qb
P680
Скорость уменьшения выхода флуоресценции зависит от скорости рекомбинации зарядов YZox
Yzox
DCMU
P680+
Mn(II)
P680*
Pheo
Q-a
Qb
P680
Скорость уменьшения выхода флуоресценции зависит от скорости рекомбинации зарядов YZox
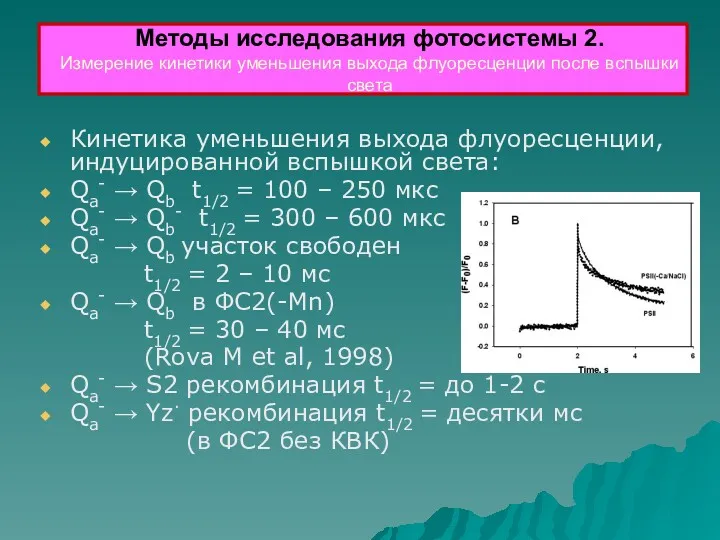
Кинетика уменьшения выхода флуоресценции, индуцированной вспышкой света:
Qa- → Qb t1/2 =
Кинетика уменьшения выхода флуоресценции, индуцированной вспышкой света:
Qa- → Qb t1/2 =
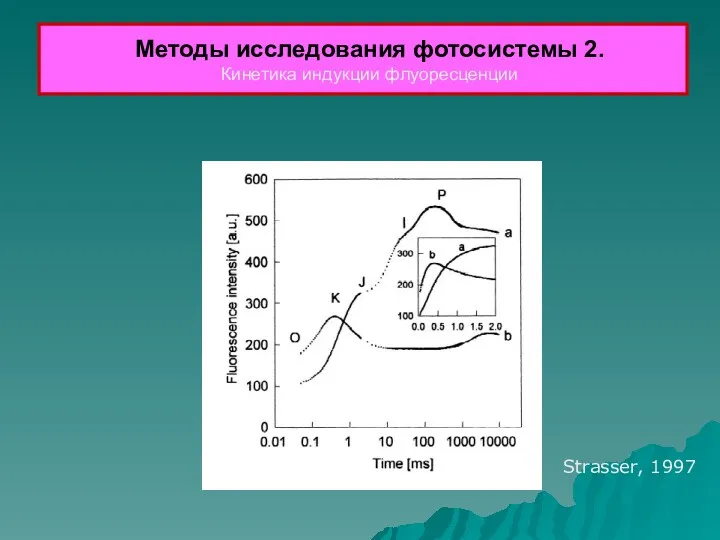
Методы исследования фотосистемы 2.
Кинетика индукции флуоресценции
Strasser, 1997
Методы исследования фотосистемы 2.
Кинетика индукции флуоресценции
Strasser, 1997

Методы исследования фотосистемы 2.
Замедленная флуоресценция
Guo & Tan, 2013
Методы исследования фотосистемы 2.
Замедленная флуоресценция
Guo & Tan, 2013
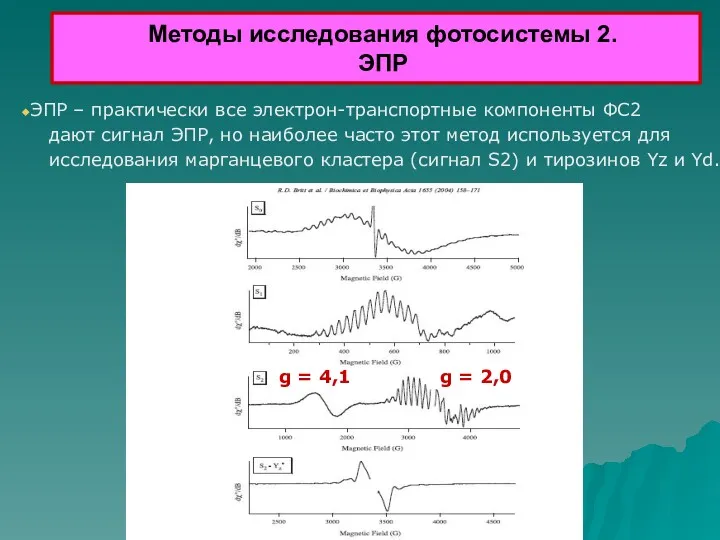
Методы исследования фотосистемы 2.
ЭПР
ЭПР – практически все электрон-транспортные компоненты ФС2
Методы исследования фотосистемы 2.
ЭПР
ЭПР – практически все электрон-транспортные компоненты ФС2

Полярография – метод измерения концентрации кислорода в среде (соответственно, возможность измерения
Полярография – метод измерения концентрации кислорода в среде (соответственно, возможность измерения

Эволюция фотосистем.
ФС1: центральные белки PsaA и PsaB имеют по 11 трансмембранных
Эволюция фотосистем.
ФС1: центральные белки PsaA и PsaB имеют по 11 трансмембранных

Рентгеновский анализ кристаллов ФС1 и ФС2 показал очень интересные результаты:
расположение
Рентгеновский анализ кристаллов ФС1 и ФС2 показал очень интересные результаты:
расположение
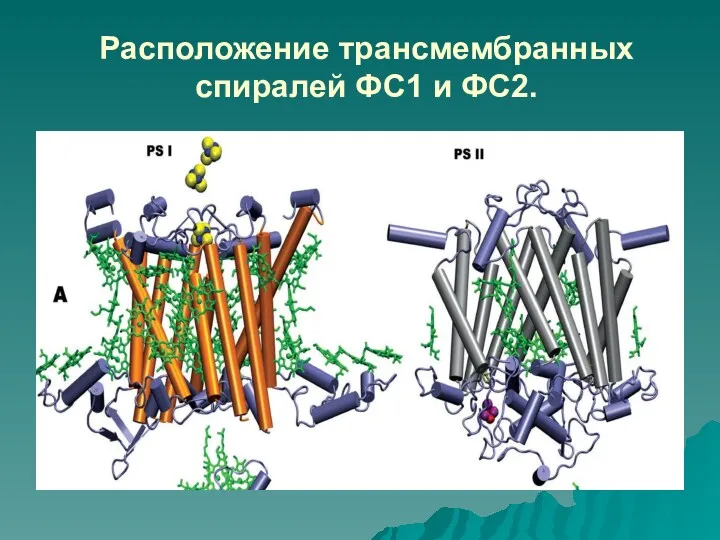
Расположение трансмембранных спиралей ФС1 и ФС2.
Расположение трансмембранных спиралей ФС1 и ФС2.
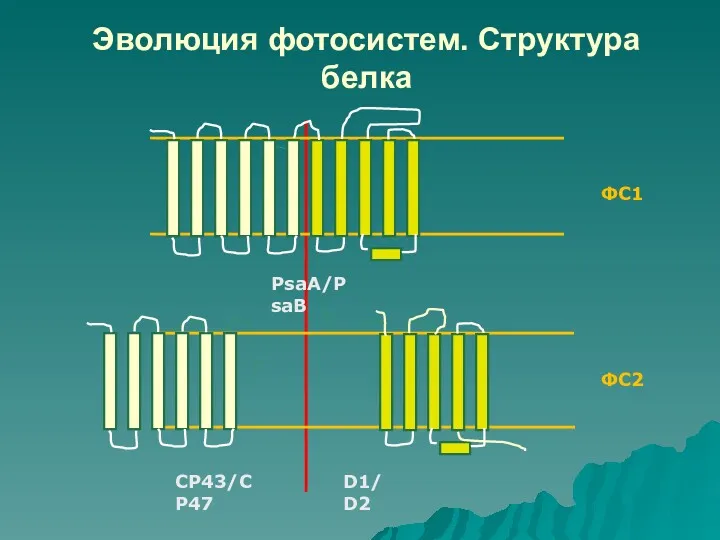
Эволюция фотосистем. Структура белка
ФС1
ФС2
СР43/CP47
D1/D2
PsaA/PsaB
Эволюция фотосистем. Структура белка
ФС1
ФС2
СР43/CP47
D1/D2
PsaA/PsaB

Большое сходство в структурной организации фотосистем 1-го и 2-го типов позволило
Большое сходство в структурной организации фотосистем 1-го и 2-го типов позволило
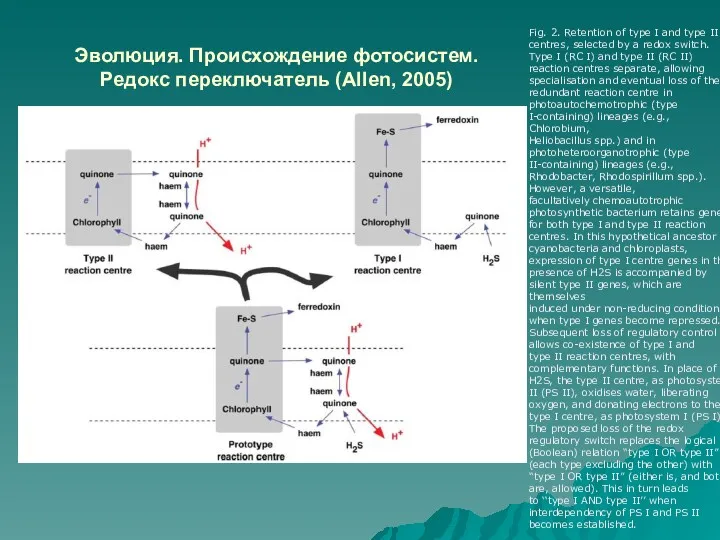
Эволюция. Происхождение фотосистем.
Редокс переключатель (Allen, 2005)
Fig. 2. Retention of type I
Эволюция. Происхождение фотосистем.
Редокс переключатель (Allen, 2005)
Fig. 2. Retention of type I
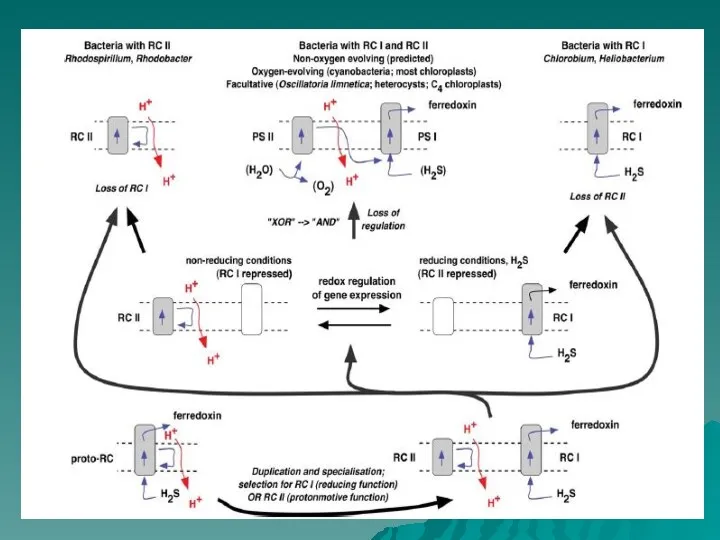
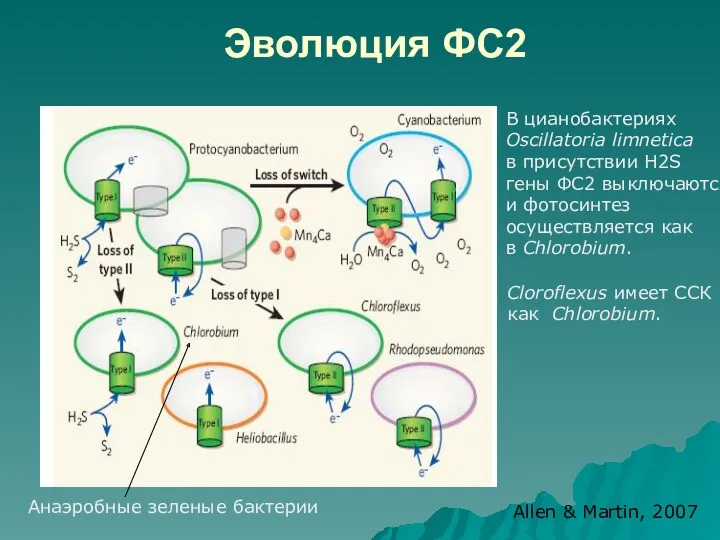
Эволюция ФС2
Анаэробные зеленые бактерии
Allen & Martin, 2007
В цианобактериях
Oscillatoria limnetica
в присутствии H2S
гены
Эволюция ФС2
Анаэробные зеленые бактерии
Allen & Martin, 2007
В цианобактериях
Oscillatoria limnetica
в присутствии H2S
гены
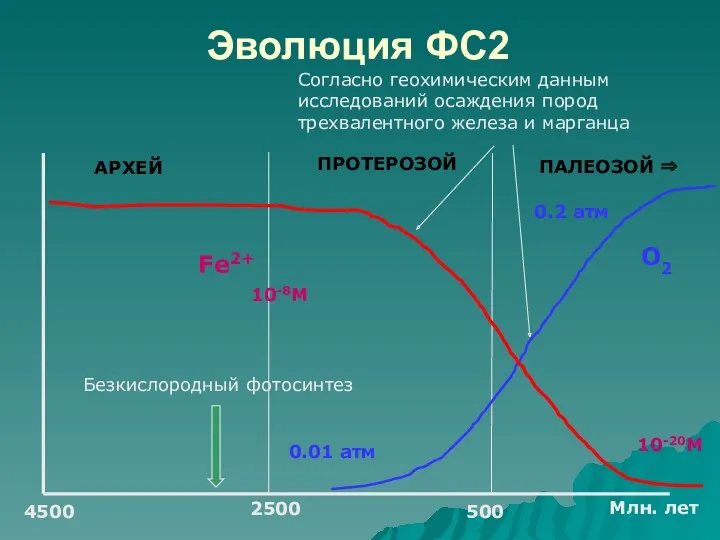
Эволюция ФС2
4500
500
АРХЕЙ
ПРОТЕРОЗОЙ
ПАЛЕОЗОЙ ⇒
2500
Млн. лет
О2
Fe2+
0.01 атм
10-20M
0.2 атм
10-8M
Безкислородный фотосинтез
Согласно геохимическим данным
исследований
Эволюция ФС2
4500
500
АРХЕЙ
ПРОТЕРОЗОЙ
ПАЛЕОЗОЙ ⇒
2500
Млн. лет
О2
Fe2+
0.01 атм
10-20M
0.2 атм
10-8M
Безкислородный фотосинтез
Согласно геохимическим данным
исследований

Возможные доноры электронов до использования воды:
Fe(II)
H2S
H2O2 – легко
Возможные доноры электронов до использования воды:
Fe(II)
H2S
H2O2 – легко

СИММЕТРИЯ МЕЖДУ СООН КОНЦЕВЫМИ УЧАСТКАМИ ПОЛИПЕПТИДОВ D1 ИD2
D1
…. SVVDSQGRVINTWAD II
СИММЕТРИЯ МЕЖДУ СООН КОНЦЕВЫМИ УЧАСТКАМИ ПОЛИПЕПТИДОВ D1 ИD2
D1
…. SVVDSQGRVINTWAD II

Примеры использования фотосистем в биотехнологии
Глобальное потребление энергии в настоящее время около
Примеры использования фотосистем в биотехнологии
Глобальное потребление энергии в настоящее время около

Примеры использования фотосистем в биотехнологии
Фотоводород.
Фотосинтетические бактерии, цианобактерии и одноклеточные зеленые
Примеры использования фотосистем в биотехнологии
Фотоводород.
Фотосинтетические бактерии, цианобактерии и одноклеточные зеленые
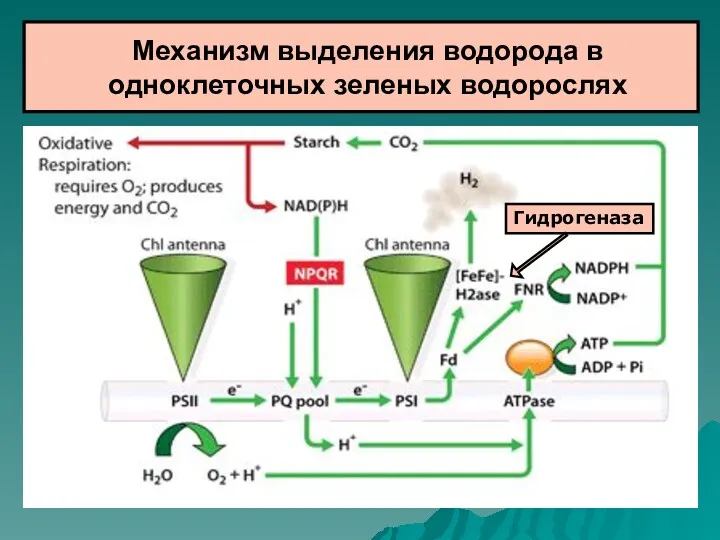
Механизм выделения водорода в одноклеточных зеленых водорослях
Гидрогеназа
Механизм выделения водорода в одноклеточных зеленых водорослях
Гидрогеназа

Источником электронов для гидрогеназы является ферредоксин, восстанавливаемый ФС1. В свою
Источником электронов для гидрогеназы является ферредоксин, восстанавливаемый ФС1. В свою
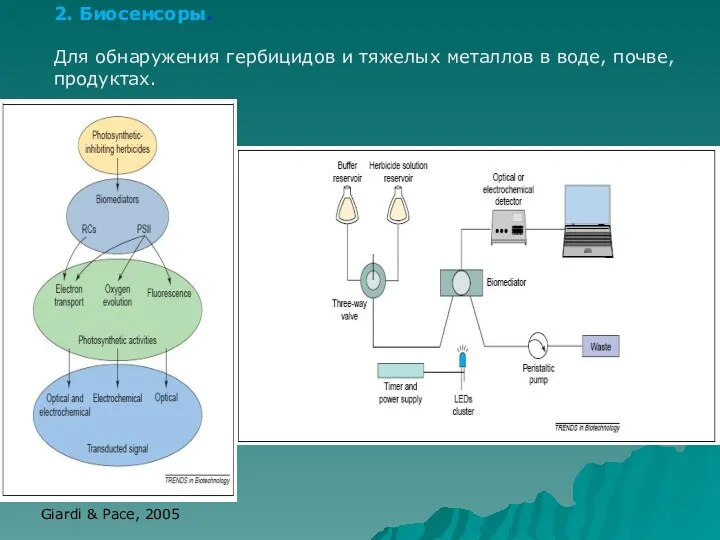
2. Биосенсоры.
Для обнаружения гербицидов и тяжелых металлов в воде, почве,
продуктах.
Giardi
2. Биосенсоры.
Для обнаружения гербицидов и тяжелых металлов в воде, почве,
продуктах.
Giardi
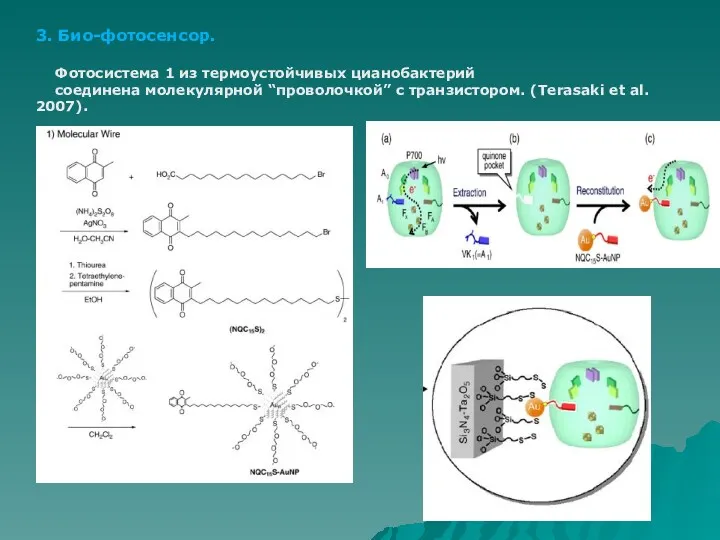
3. Био-фотосенсор.
Фотосистема 1 из термоустойчивых цианобактерий
соединена молекулярной “проволочкой”
3. Био-фотосенсор.
Фотосистема 1 из термоустойчивых цианобактерий
соединена молекулярной “проволочкой”
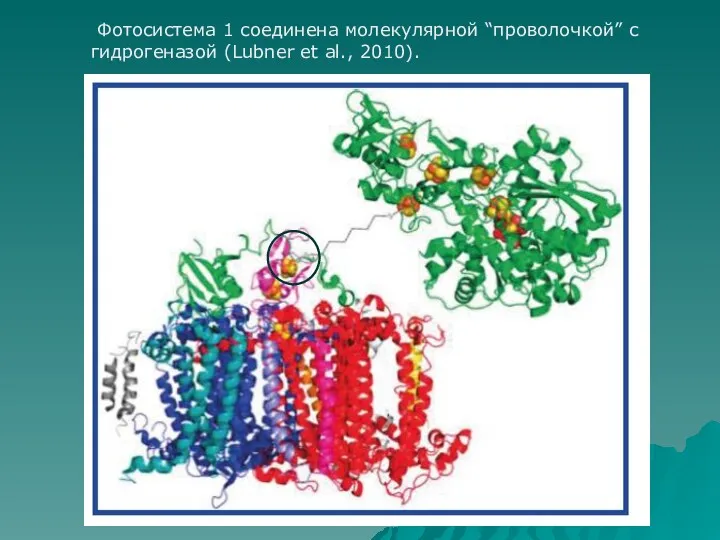
Фотосистема 1 соединена молекулярной “проволочкой” с гидрогеназой (Lubner et al.,
Фотосистема 1 соединена молекулярной “проволочкой” с гидрогеназой (Lubner et al.,
 Чистые вещества и смеси
Чистые вещества и смеси Мыло и синтетические моющие средства
Мыло и синтетические моющие средства Материаловедение. Лекция 1
Материаловедение. Лекция 1 Залізо. Фізичні та хімічні властивості
Залізо. Фізичні та хімічні властивості Почему мыло пенится
Почему мыло пенится Алюминий
Алюминий Гидрогеохимия урана и тория
Гидрогеохимия урана и тория Природные источники углеводородов
Природные источники углеводородов Химические волокна (7 класс)
Химические волокна (7 класс) Аргентум, или серебро
Аргентум, или серебро Алкины. Физические и химические свойства
Алкины. Физические и химические свойства Спирты: общая характеристика
Спирты: общая характеристика Бескислородные керамические материалы
Бескислородные керамические материалы Химическая посуда и лабораторное оборудование
Химическая посуда и лабораторное оборудование Objectives and background. Apply for an unknown mixture
Objectives and background. Apply for an unknown mixture Процессы, проходящие в растворах (продолжение темы)
Процессы, проходящие в растворах (продолжение темы) The Molecules of Life
The Molecules of Life Зеленая химия полиуретанов: механизм, катализ, проблемы использования возобновляемого сырья
Зеленая химия полиуретанов: механизм, катализ, проблемы использования возобновляемого сырья Электродные процессы
Электродные процессы Физические и химические свойства алмаза. Алмазная промышленность в России
Физические и химические свойства алмаза. Алмазная промышленность в России Кислоты
Кислоты Вклад ученых-химиков в победу в Великой Отечественной войне
Вклад ученых-химиков в победу в Великой Отечественной войне Окислительно-восстановительные реакции
Окислительно-восстановительные реакции Основні класи неорганічних сполук
Основні класи неорганічних сполук Совершенствование урока химии в условиях ФГОС
Совершенствование урока химии в условиях ФГОС Игра Химические элементы (формулы и названия)
Игра Химические элементы (формулы и названия) Химическая кинетика и катализ
Химическая кинетика и катализ Органічні сполуки. 3D моделі органічних сполук
Органічні сполуки. 3D моделі органічних сполук