- Хімія та обмін вуглеводів

Содержание
- 2. Функції вуглеводів Енергетична (при окисненні 1 г вуглеводів виділяється ≈ 16,9 кДж енергії) Структурна (в комплексі
- 3. ВУГЛЕВОДИ залежно від здатності до гідролізу ділять на: Моносахариди а) від кількості атомів Карбону в ланцюзі
- 4. Моносахариди Моносахариди – похідні багатоатомних спиртів, які містять карбонільну групу. В залежності від положення в молекулі
- 5. Альдози і кетози
- 6. Моносахариди зображуються проекційними формулами Фішера, тобто у вигляді проекції тетраедричної моделі атомів Карбону на площину креслення.
- 7. Ізомерія моносахаридів Ізомерія моносахаридів обумовлена: 1) наявністю альдегідної або кетонної групи; 2) наявністю асиметричних атомів Карбону;
- 8. Таутомерія – здатність речовин існувати в декількох формах, які переходять одна в одну і знаходяться в
- 9. α-D-глюкопіраноза D-глюкоза β-D-глюкопіраноза
- 10. Моносахариди, які мають циклічну форму, зручно відображати перспективними формулами Хеуорса. Вони представляють собою ідеалізовані п'яти- і
- 12. Дисахариди Це складні вуглеводи, молекула яких складається з двох (однакових або різних) залишків моносахариду. Дисахариди ділять
- 13. Невідновлюючі дисахариди (сахароза, трегалоза) Це дисахариди, утворені в результаті виділення води за рахунок напівацетальних гідроксилів обох
- 14. Відновлюючі дисахариди (мальтоза, целобіоза, лактоза) Це дисахариди, утворені в результаті виділення води за рахунок напівацетального гідроксилу
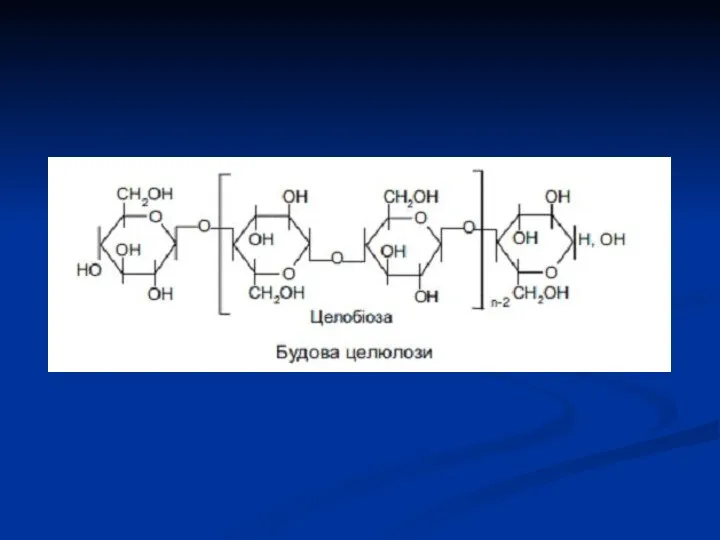
- 15. Целобіоза утворюється при неповному гідролізі полісахариду целюлози (клітковини). Целобіоза побудована з двох молекул D‑глюкози, які зв'язані
- 16. Дисахариди їжі
- 17. Дисахариди їжі
- 18. Полісахариди Полісахариди – це складні вуглеводи, молекули яких складаються з великої кількості моносахаридів (більше 10). За
- 19. Крохмаль представляє собою суміш двох полісахаридів, побудованих з α, D‑глюкопіранозних ланок: амілози (10-20%) і амілопектину (80-90%).
- 20. Амілоза: Амілопектин:
- 21. Будова крохмалю
- 22. Глікоген полісахарид, утворений залишками глюкози; основний запасний вуглевод людини і тварин. Глікоген (також іноді його називають
- 24. Целюлоза (клітковина) найбільш розповсюджений рослинний полісахарид. Він має велику механічну міцність і виконує роль опорного матеріалу
- 27. Хітин (фр. chitine, від грец. chiton: хітон - одяг, шкіра, оболонка) – природна ссполука з групи
- 29. Хітозан застосовують в якості корму для тварин, для виготовлення продуктів харчування і косметики, використовують в продуктах
- 31. Інулін (C6H10O5)n — органічна речовина з групи полісахаридів, полімер D-фруктози. Інулін — білий порошок, який легко
- 33. Гетерополісахариди Це складні вуглеводи, молекули яких побудовані із залишків різних моносахаридів, їх похідних та інших сполук.
- 34. Гіалуронова кислота Це гетерополісахарид, побудований із залишків молекул глюкуронової та оцтової кислот, глюкозаміну. Структурною одиницею вуглеводу
- 35. Гіалуронова кислота є хімічною основою склоподібного тіла ока, пупкового канатику, синовії, блискучої оболонки яйцеклітини, капсул деяких
- 37. Хондроітинсульфатна кислота Є продуктом полімеризації N-ацетилгалактозамінсульфату і глюкуронової кислоти, з'єднаних між собою β-1,3- і β-1,4-глікозидними зв'язками.
- 38. Хондроітинсульфатна кислота – обов'язкова складова частина хрящів (до 40% сухої маси), кісток, основної речовини сполучної тканини,
- 40. Гепарин Молекула цієї речовини утворена залишками α-Д-глюкозаміну, глюкуронової та сульфатної кислот.
- 41. Гепарин – білий аморфний порошок, розчинний у воді, стійкий до нагрівання. В організмі виробляється тучними клітинами
- 43. Переварювання вуглеводів у ротовій порожнині
- 44. Переварювання вуглеводів у кишечнику У дванадцятипалій кишці рН середовища шлункового вмісту нейтралізується, так як секрет підшлункової
- 45. Переварювання вуглеводів А) крохмаль декстрини мальтоза глікоген Б) мальтоза глюкоза кров В) сахароза глюкоза кров фруктоза
- 46. Переварювання вуглеводів
- 47. Всмоктування моносахаридів у кишечнику Транспорт моносахаридів у клітини слизової оболонки кишечника може здійснюватися різними способами: шляхом
- 48. Всмоктування моносахаридів з кишечника відбувається шляхом простої дифузії за допомогою спеціальних білків-переносників (транспортерів). Окрім того, глюкоза
- 49. Переварювання вуглеводів у жуйних
- 50. За добу утворюється 3 – 5 кг летких карбонових кислот, серед яких: Оцтова – 70 -
- 51. КОНЦЕНТРАЦІЯ ГЛЮКОЗИ В КРОВІ ДРХ – 30-40 мг% (1,7 – 2,2 ммоль/л) ВРХ – 60-80 мг%
- 52. Шляхи використання глюкози в тканинах
- 53. Шляхи окиснення глюкози І Анаеробне окиснення: Гліколіз Глікогеноліз ІІ Аеробне окиснення: Через цикл трикарбонових кислот (ЦТК)
- 54. Основні шляхи використання вуглеводів 1. Синтез глікогену. 2. Окиснення а) анаеробні − гліколіз і глікогеноліз б)
- 55. Реакції гліколізу 1.
- 56. 2. 3.
- 57. 4. 5.
- 58. 6.1. 6.2.
- 59. 6.3. 7.
- 60. 8. 9.
- 61. 10. 11.
- 62. Анаеробний гліколіз Анаеробним гліколізом називають процес розщеплення глюкози з утворенням в якості кінцевого продукту лактату. Цей
- 63. Глікогеноліз
- 64. Енергетичний баланс гліколізу і глікогенолізу
- 65. Катаболізм глюкози Окиснення глюкози до СО2 і Н2О (аеробний розпад). Аеробний розпад глюкози можна виразити сумарним
- 66. Етапи аеробного гліколізу В аеробному гліколізі можна виділити 2 етапи. 1. Підготовчий етап, в ході якого
- 67. Центральним процесом катаболізму є цикл лимонної кислоти Головна функція циклу лимонної кислоти - окиснення ацетогрупи, яка
- 68. Ц.Т.К. – послідовні перетворення ди- і трикарбонових кислот, в ході яких оцтова кислота, що включається в
- 69. Цикл лимонної кислоти (трикарбонових кислот, Кребса)
- 70. Загальна схема цитратного циклу
- 71. Цикл Кребса складається з 8 послідовних ферментативних реакцій: Синтез лимонної кислоти або цитрату (фермент -цитратсинтетаза); Синтез
- 72. 1. 2.
- 73. 3. ізолимонна кислота α-кетоглутарова кислота 4. α-кетоглутарова кислота
- 74. 5. 6.
- 75. 7. 8.
- 76. В результаті 1 витку циклу Кребса енергія виділяється у вигляді тепла і 12 молекул АТФ 1
- 77. Регуляція ЦТК Алостерична регуляція – здійснюється за рахунок 3 регуляторних ферментів: цитратсинтази (інгібується АТФ і НАДН,
- 78. Біологічна роль ЦТК: Інтегративна – він є метаболічним колектором, що об'єднує шляхи катаболізму вуглеводів, ліпідів та
- 79. Енергетична ефективність аеробного окиснення глюкози 1М глюкози 38 АТФ в тому числі: А) гліколіз 2 МК
- 80. ПЕНТОЗОФОСФАТНИЙ ШЛЯХ ПЕРЕТВОРЕННЯ ГЛЮКОЗИ Пентозофосфатний шлях, який називається також гексомонофосфатним шунтом, слугує альтернативним шляхом окиснення глюкозо-6-фосфату.
- 81. Пентозофосфатний шлях забезпечує клітини рибозою для синтезу пуринових та піримідинових нуклеотидів і гідрованим коферментом НАДФH, який
- 82. Окисний етап В окисній частині пентозофосфатного шляху глюкозо-6-фосфат підлягає окисному декарбоксилуванню, в результаті якого утворюються пентози.
- 83. Окисний етап пентозофосфатного шляху
- 84. Реакції окисного етапу є основним джерелом НАДФH в клітинах. Гідровані коферменти забезпечують Гідрогеном біосинтетичні процеси, окисно-відновні
- 85. Перетворення рибулозо-5-фосфату
- 86. МЕТАБОЛІЗМ ГЛЮКУРОНОВОЇ КИСЛОТИ Окрім гліколізу і пентозофосфатного шляху, що забезпечують засвоєння глюкози, в організмі тварин є
- 87. Схема синтезу вітаміну С
- 88. Біологічне значення глюкуронової кислоти проявляється в тому, що вона здатна зв'язуватися з певними ендогенними і екзогенними
- 89. Синтез глікогену (глікогенез)
- 90. Функції глікогену в печінці і м'язах
- 91. Регуляція глікогенезу Активує цей процес інсулін. Інгібують цей процес: глюкагон, адреналін.
- 92. Глюконеогенез це синтез глюкози з невуглеводних метаболітів (молочної кислоти, карбонових кислот, гліцерину, залишків амінокислот), тобто тих,
- 93. Включення субстратів у глюконеогенез
- 94. Цикл Корі (глюкозо-лактатний цикл). 1. Надходження лактату з м'язу, що скорочується, з током крові в печінку;
- 95. Лактат, що утворився в м'язах, які інтенсивно працюють або в клітинах з переважаючим анаеробним шляхом катаболізму
- 96. Цикл Корі виконує 2 важливі функції: 1 — забезпечує утилізацію лактату; 2 — запобігає накопиченню лактату
- 97. Синтез глюкози з амінокислот В умовах голодування частина білків м'язової тканини розпадається до амінокислот, які далі
- 98. Синтез глюкози з гліцеролу Гліцерол утворюється в результаті гідролізу триацил-гліцеролів, головним чином в жировій тканині. Використовувати
- 99. Гормони, регулятори рівня глюкози а) гіпоглікемічної дії: інсулін б) гіперглікемічної дії: Основні: адреналін – у всіх
- 100. Регуляція рівня цукру в крові Центр вуглеводного обміну гіпоталамусу гіпофіз АКТГ,ТТГ Залози внутрішньої секреції ГОРМОНИ адреналін
- 101. Біохімічні функції інсуліну Це головний анаболічний гормон. Підвищує проникність клітин печінки, нирок, жирової тканини та інших
- 102. Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу): глюкоза + АТФ → глюкозо-6-фосфат + АДФ Глюкозо-6-фосфат
- 103. Гіпоглікемічна дія інсуліну
- 104. Гіпофункція по інсуліну Захворювання цукровий діабет. Основні його ознаки: 1) гіперглікемія; 2) глюкозурія; 3) кетонемія; 4)
- 106. Скачать презентацию

Функції вуглеводів
Енергетична
(при окисненні 1 г вуглеводів виділяється ≈ 16,9 кДж
Функції вуглеводів
Енергетична
(при окисненні 1 г вуглеводів виділяється ≈ 16,9 кДж

ВУГЛЕВОДИ залежно від здатності до гідролізу ділять на:
Моносахариди
а) від кількості атомів
ВУГЛЕВОДИ залежно від здатності до гідролізу ділять на:
Моносахариди
а) від кількості атомів

Моносахариди
Моносахариди – похідні багатоатомних спиртів, які містять карбонільну групу. В залежності
Моносахариди
Моносахариди – похідні багатоатомних спиртів, які містять карбонільну групу. В залежності
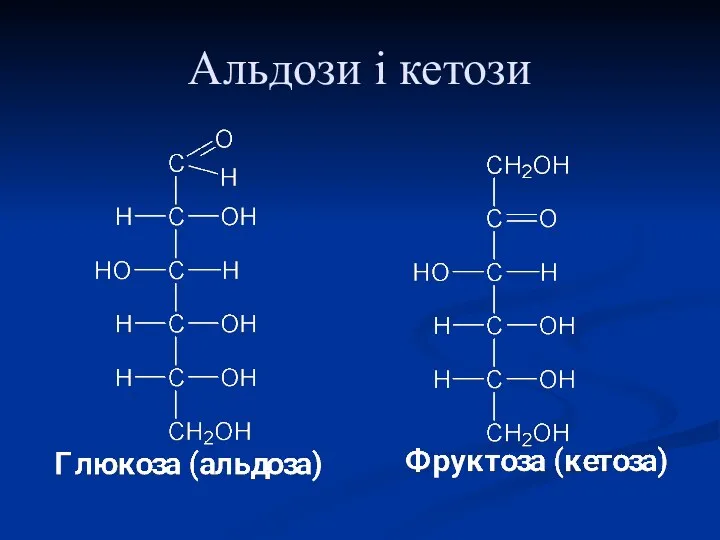
Альдози і кетози
Альдози і кетози

Моносахариди зображуються проекційними формулами Фішера, тобто у вигляді проекції тетраедричної моделі
Моносахариди зображуються проекційними формулами Фішера, тобто у вигляді проекції тетраедричної моделі

Ізомерія моносахаридів
Ізомерія моносахаридів обумовлена:
1) наявністю альдегідної або кетонної групи;
2)
Ізомерія моносахаридів
Ізомерія моносахаридів обумовлена:
1) наявністю альдегідної або кетонної групи;
2)

Таутомерія – здатність речовин існувати в декількох формах, які переходять одна
Таутомерія – здатність речовин існувати в декількох формах, які переходять одна
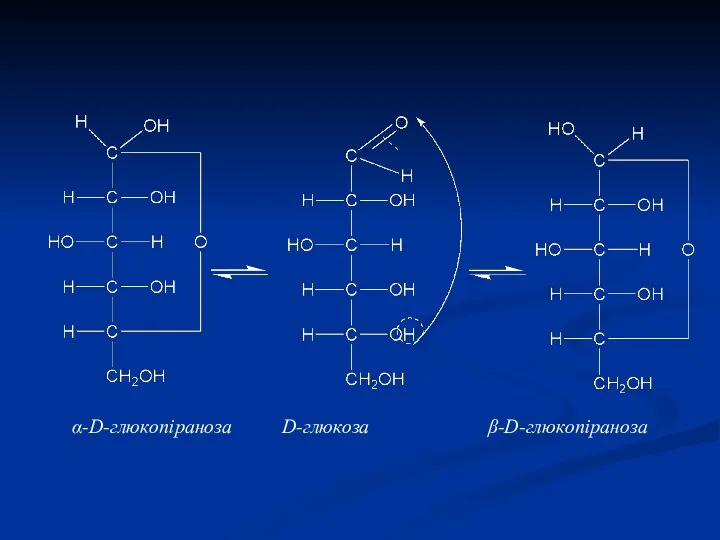
α-D-глюкопіраноза D-глюкоза β-D-глюкопіраноза
α-D-глюкопіраноза D-глюкоза β-D-глюкопіраноза

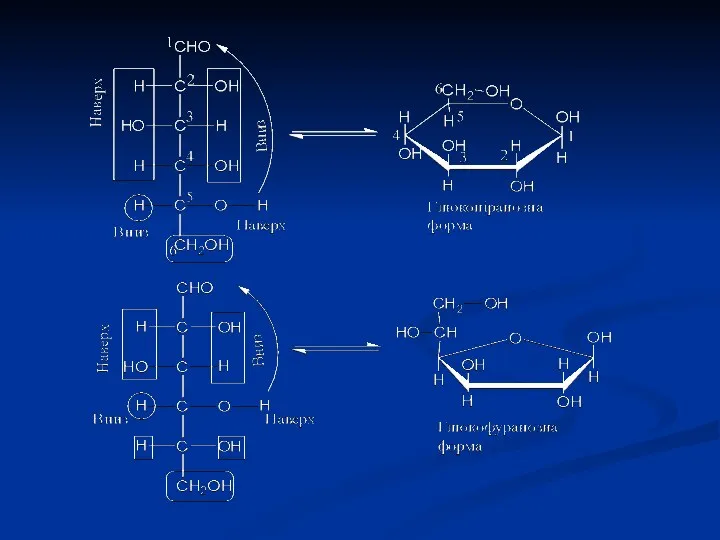
Моносахариди, які мають циклічну форму, зручно відображати перспективними формулами Хеуорса. Вони
Моносахариди, які мають циклічну форму, зручно відображати перспективними формулами Хеуорса. Вони

Дисахариди
Це складні вуглеводи, молекула яких складається з двох (однакових або
Дисахариди
Це складні вуглеводи, молекула яких складається з двох (однакових або

Невідновлюючі дисахариди (сахароза, трегалоза)
Це дисахариди, утворені в результаті виділення води за
Невідновлюючі дисахариди (сахароза, трегалоза)
Це дисахариди, утворені в результаті виділення води за

Відновлюючі дисахариди
(мальтоза, целобіоза, лактоза)
Це дисахариди, утворені в результаті виділення води
Відновлюючі дисахариди
(мальтоза, целобіоза, лактоза)
Це дисахариди, утворені в результаті виділення води

Целобіоза утворюється при неповному гідролізі полісахариду целюлози (клітковини). Целобіоза побудована з
Целобіоза утворюється при неповному гідролізі полісахариду целюлози (клітковини). Целобіоза побудована з

Дисахариди їжі
Дисахариди їжі

Дисахариди їжі
Дисахариди їжі

Полісахариди
Полісахариди – це складні вуглеводи, молекули яких складаються з великої кількості
Полісахариди
Полісахариди – це складні вуглеводи, молекули яких складаються з великої кількості

Крохмаль
представляє собою суміш двох полісахаридів, побудованих з α, D‑глюкопіранозних ланок: амілози
Крохмаль
представляє собою суміш двох полісахаридів, побудованих з α, D‑глюкопіранозних ланок: амілози
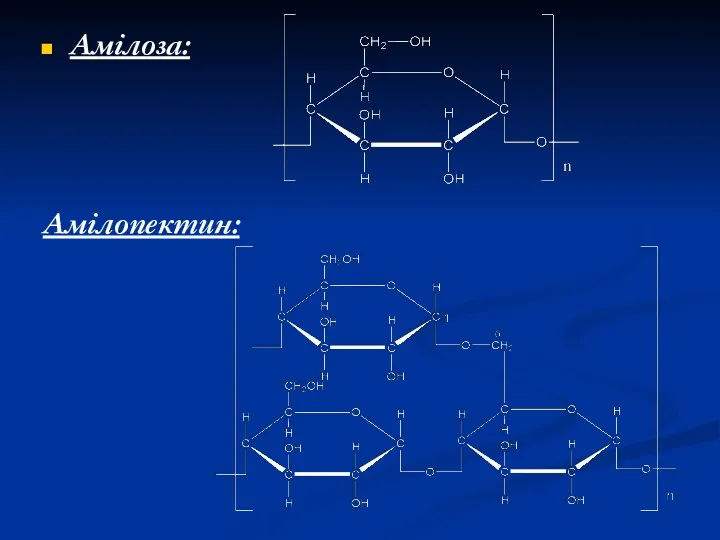
Амілоза:
Амілопектин:
Амілоза:
Амілопектин:
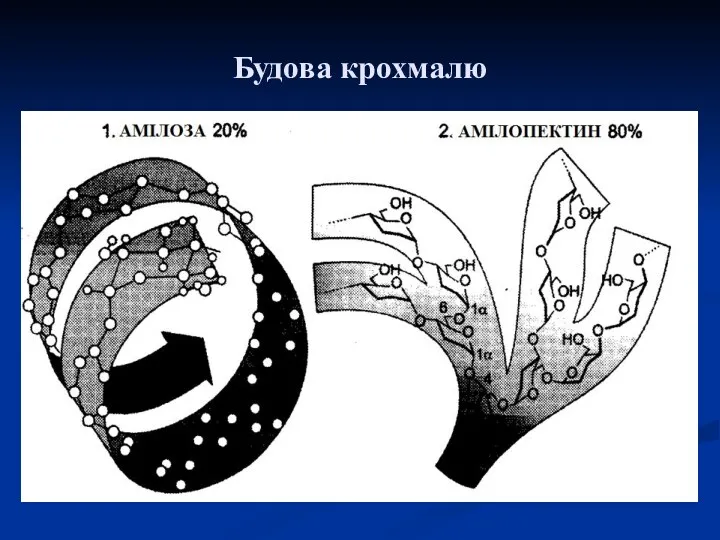
Будова крохмалю
Будова крохмалю

Глікоген
полісахарид, утворений залишками глюкози; основний запасний вуглевод людини і тварин.
Глікоген
Глікоген
полісахарид, утворений залишками глюкози; основний запасний вуглевод людини і тварин.
Глікоген
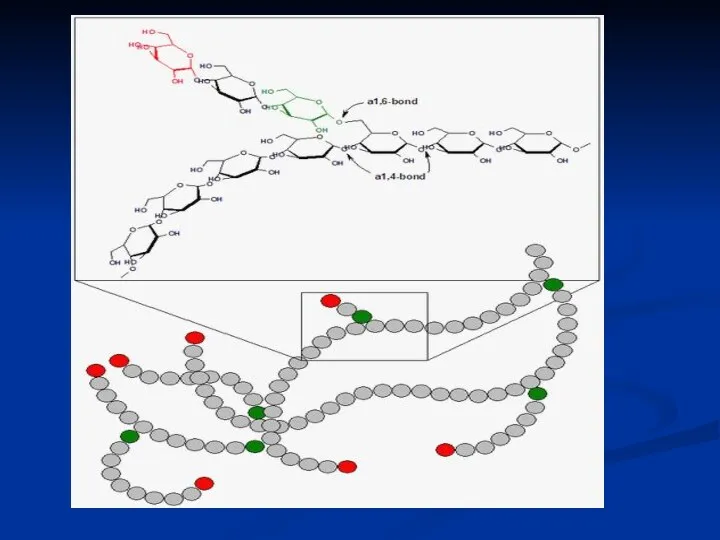

Целюлоза (клітковина)
найбільш розповсюджений рослинний полісахарид. Він має велику механічну міцність і
Целюлоза (клітковина)
найбільш розповсюджений рослинний полісахарид. Він має велику механічну міцність і


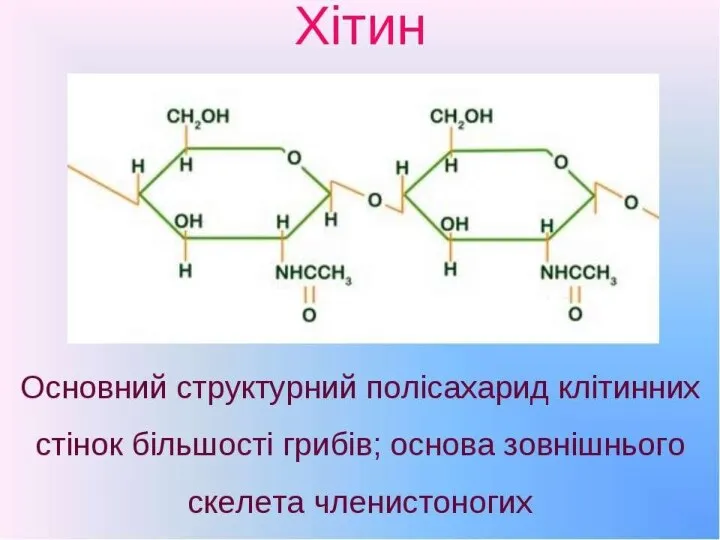
Хітин
(фр. chitine, від грец. chiton: хітон - одяг, шкіра, оболонка) –
Хітин
(фр. chitine, від грец. chiton: хітон - одяг, шкіра, оболонка) –

Хітозан застосовують в якості корму для тварин, для виготовлення продуктів харчування
Хітозан застосовують в якості корму для тварин, для виготовлення продуктів харчування


Інулін
(C6H10O5)n — органічна речовина з групи полісахаридів, полімер D-фруктози.
Інулін — білий
Інулін
(C6H10O5)n — органічна речовина з групи полісахаридів, полімер D-фруктози.
Інулін — білий


Гетерополісахариди
Це складні вуглеводи, молекули яких побудовані із залишків різних моносахаридів, їх
Гетерополісахариди
Це складні вуглеводи, молекули яких побудовані із залишків різних моносахаридів, їх
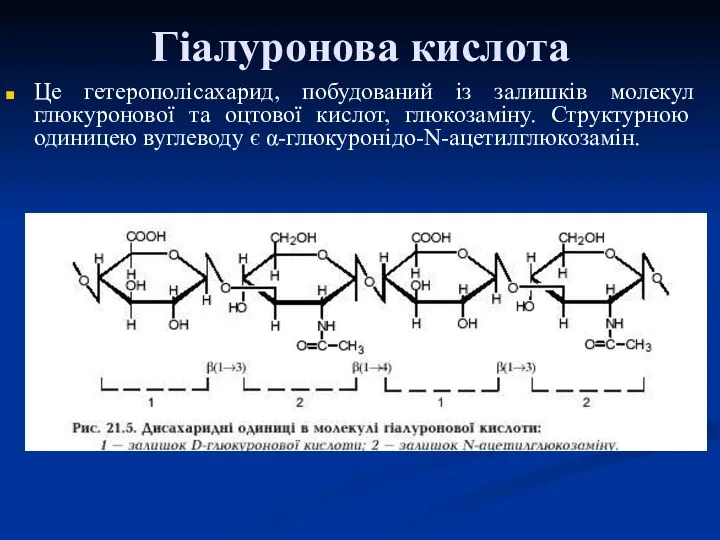
Гіалуронова кислота
Це гетерополісахарид, побудований із залишків молекул глюкуронової та оцтової кислот,
Гіалуронова кислота
Це гетерополісахарид, побудований із залишків молекул глюкуронової та оцтової кислот,

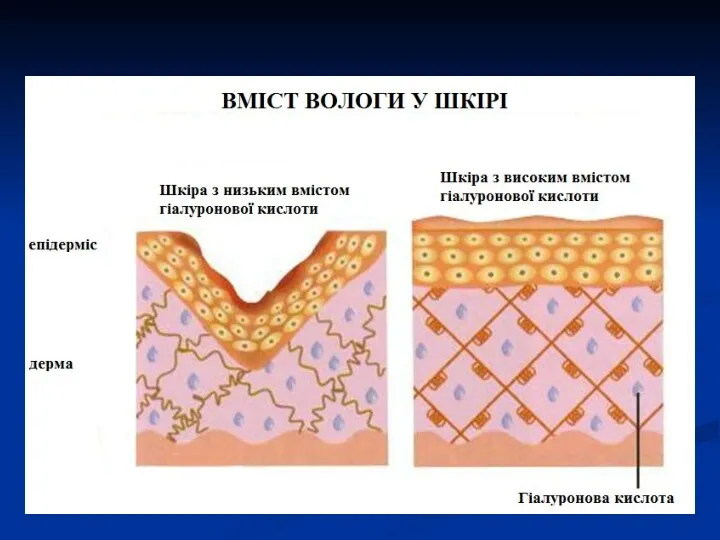
Гіалуронова кислота є хімічною основою склоподібного тіла ока, пупкового канатику, синовії,
Гіалуронова кислота є хімічною основою склоподібного тіла ока, пупкового канатику, синовії,
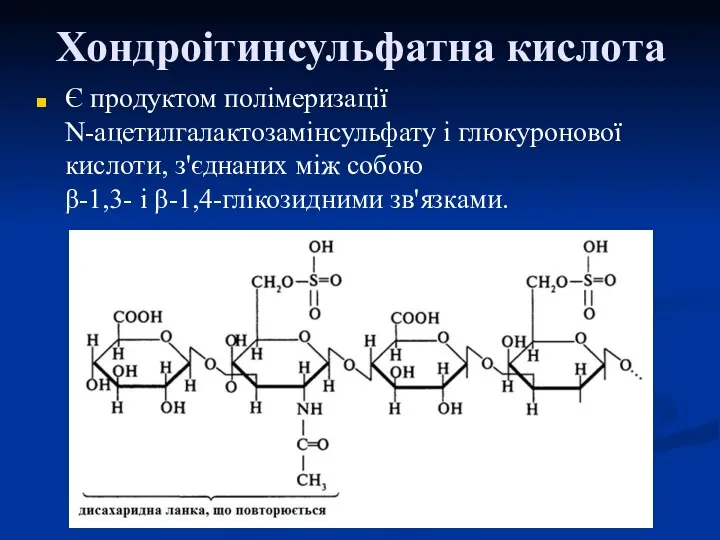
Хондроітинсульфатна кислота
Є продуктом полімеризації N-ацетилгалактозамінсульфату і глюкуронової кислоти, з'єднаних між собою
Хондроітинсульфатна кислота
Є продуктом полімеризації N-ацетилгалактозамінсульфату і глюкуронової кислоти, з'єднаних між собою

Хондроітинсульфатна кислота – обов'язкова складова частина хрящів (до 40% сухої маси),
Хондроітинсульфатна кислота – обов'язкова складова частина хрящів (до 40% сухої маси),
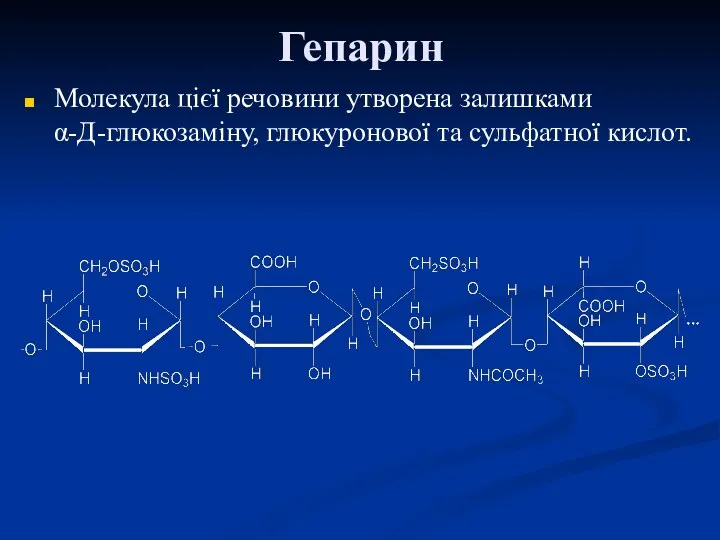
Гепарин
Молекула цієї речовини утворена залишками α-Д-глюкозаміну, глюкуронової та сульфатної кислот.
Гепарин
Молекула цієї речовини утворена залишками α-Д-глюкозаміну, глюкуронової та сульфатної кислот.

Гепарин – білий аморфний порошок, розчинний у воді, стійкий до нагрівання.
Гепарин – білий аморфний порошок, розчинний у воді, стійкий до нагрівання.

Переварювання вуглеводів у ротовій порожнині
Переварювання вуглеводів у ротовій порожнині

Переварювання вуглеводів у кишечнику
У дванадцятипалій кишці рН середовища шлункового вмісту нейтралізується,
Переварювання вуглеводів у кишечнику
У дванадцятипалій кишці рН середовища шлункового вмісту нейтралізується,

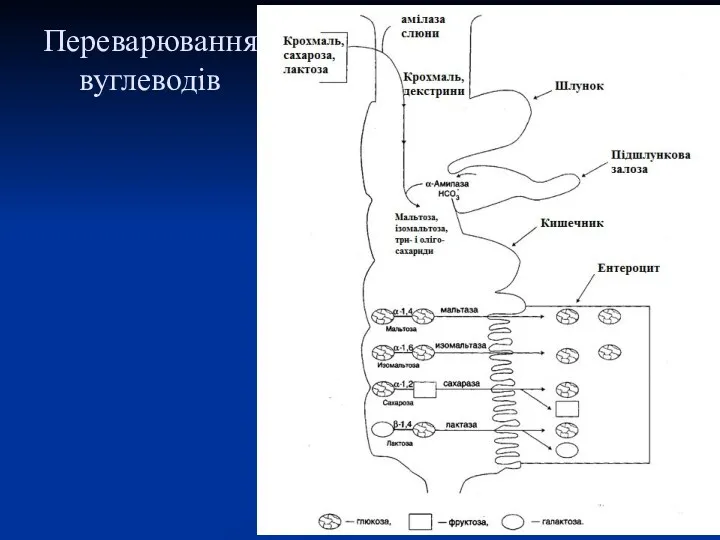
Переварювання вуглеводів
А) крохмаль декстрини мальтоза
глікоген
Б) мальтоза глюкоза кров
В) сахароза глюкоза
Переварювання вуглеводів
А) крохмаль декстрини мальтоза
глікоген
Б) мальтоза глюкоза кров
В) сахароза глюкоза
Переварювання
вуглеводів
Переварювання
вуглеводів

Всмоктування моносахаридів у кишечнику
Транспорт моносахаридів у клітини слизової оболонки кишечника може
Всмоктування моносахаридів у кишечнику
Транспорт моносахаридів у клітини слизової оболонки кишечника може
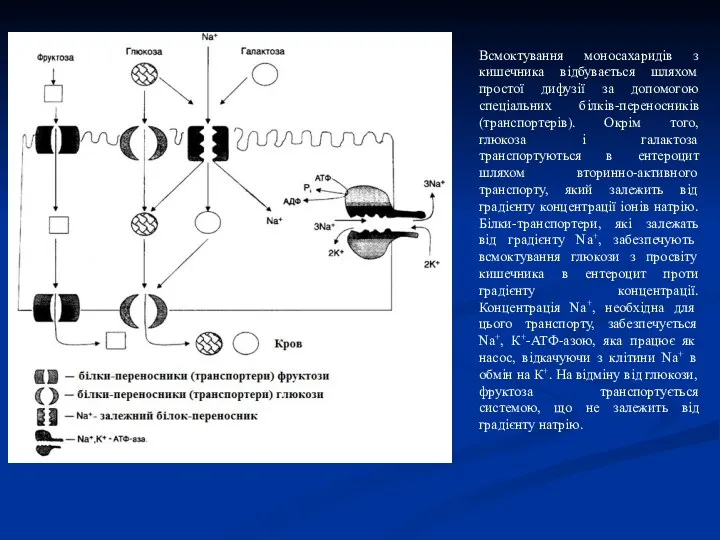
Всмоктування моносахаридів з кишечника відбувається шляхом простої дифузії за допомогою спеціальних
Всмоктування моносахаридів з кишечника відбувається шляхом простої дифузії за допомогою спеціальних

Переварювання вуглеводів у жуйних
Переварювання вуглеводів у жуйних

За добу утворюється 3 – 5 кг летких карбонових кислот, серед
За добу утворюється 3 – 5 кг летких карбонових кислот, серед

КОНЦЕНТРАЦІЯ ГЛЮКОЗИ В КРОВІ
ДРХ – 30-40 мг% (1,7 – 2,2 ммоль/л)
ВРХ
КОНЦЕНТРАЦІЯ ГЛЮКОЗИ В КРОВІ
ДРХ – 30-40 мг% (1,7 – 2,2 ммоль/л)
ВРХ
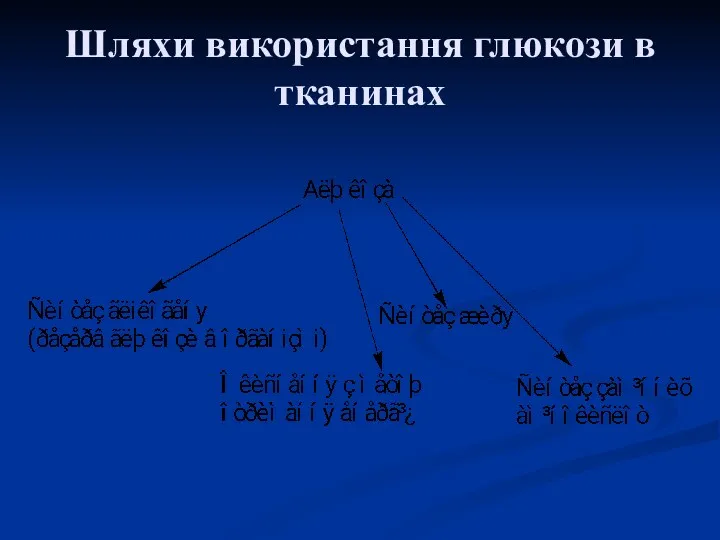
Шляхи використання глюкози в тканинах
Шляхи використання глюкози в тканинах

Шляхи окиснення глюкози
І Анаеробне окиснення:
Гліколіз
Глікогеноліз
ІІ Аеробне окиснення:
Через цикл трикарбонових кислот (ЦТК)
Пентозо-фосфатний
Шляхи окиснення глюкози
І Анаеробне окиснення:
Гліколіз
Глікогеноліз
ІІ Аеробне окиснення:
Через цикл трикарбонових кислот (ЦТК)
Пентозо-фосфатний

Основні шляхи використання вуглеводів
1. Синтез глікогену.
2. Окиснення
а) анаеробні −
Основні шляхи використання вуглеводів
1. Синтез глікогену.
2. Окиснення
а) анаеробні −
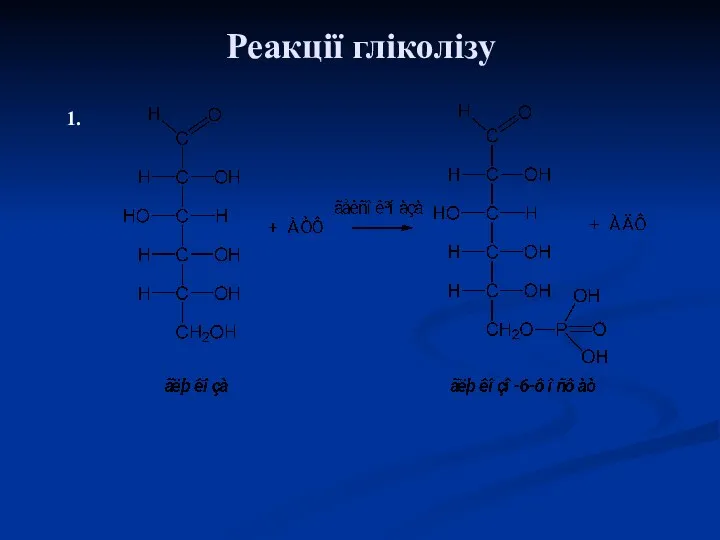
Реакції гліколізу
1.
Реакції гліколізу
1.
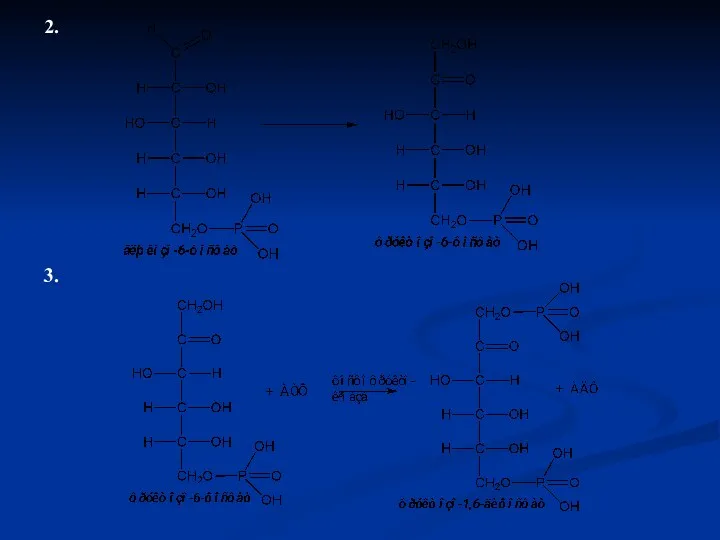
2.
3.
2.
3.
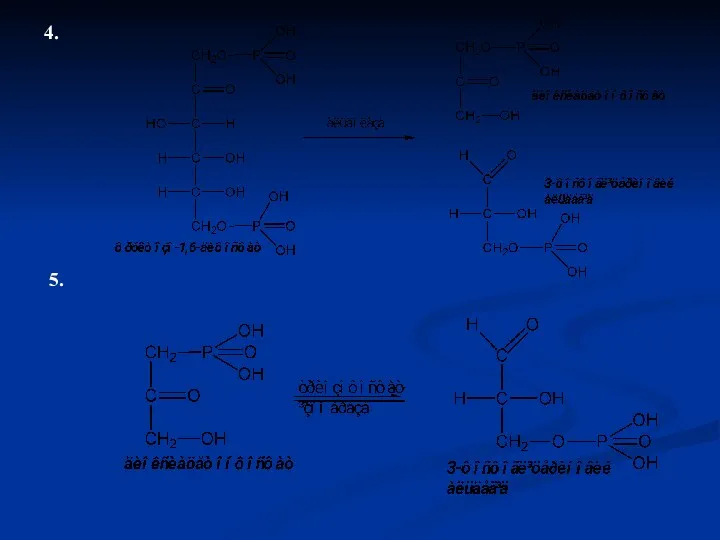
4.
5.
4.
5.
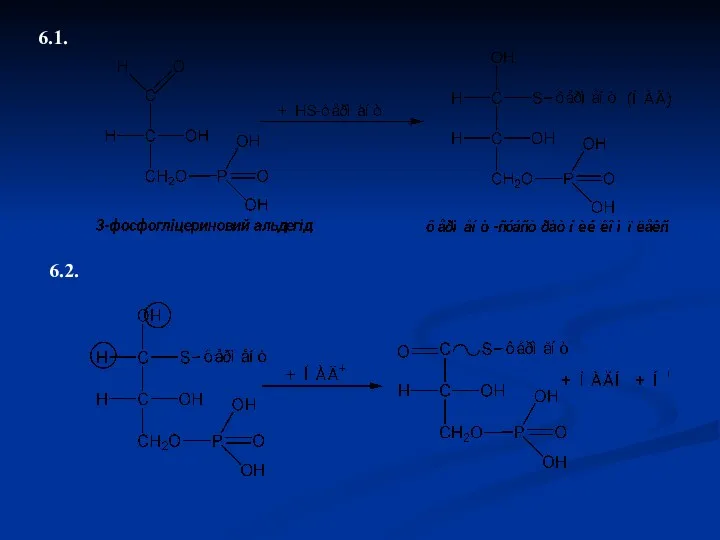
6.1.
6.2.
6.1.
6.2.
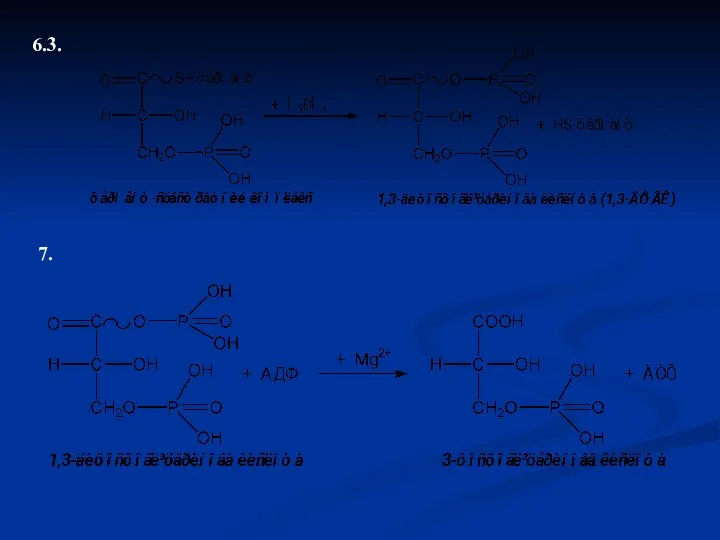
6.3.
7.
6.3.
7.
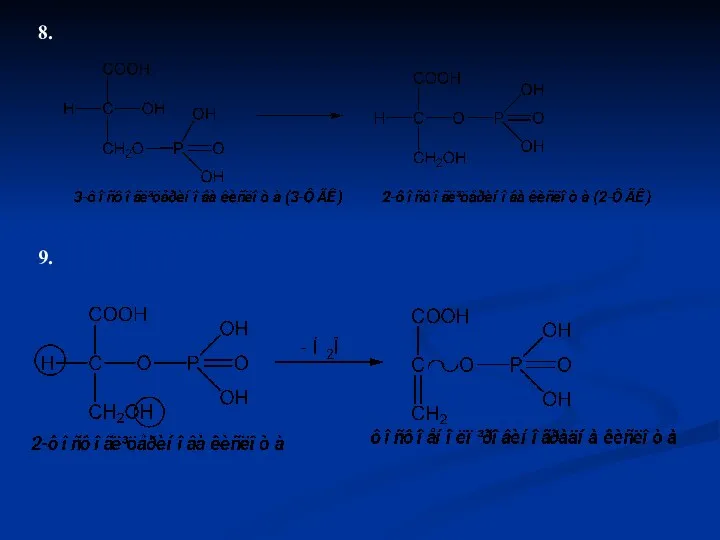
8.
9.
8.
9.
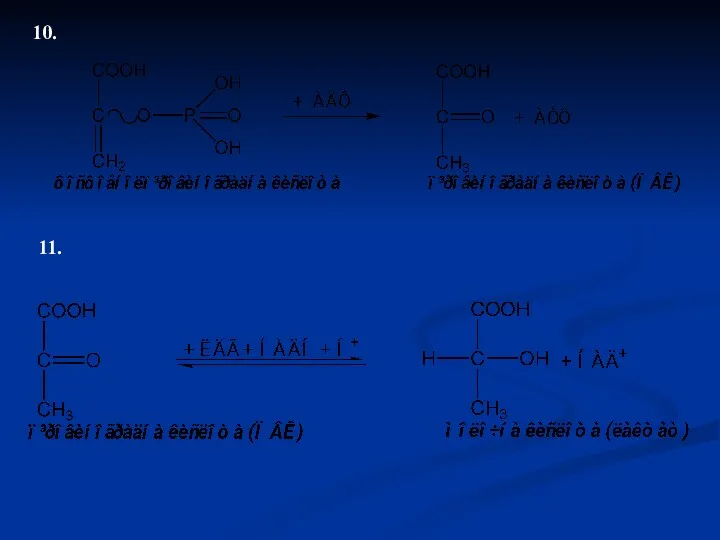
10.
11.
10.
11.

Анаеробний гліколіз
Анаеробним гліколізом називають процес розщеплення глюкози з утворенням в якості
Анаеробний гліколіз
Анаеробним гліколізом називають процес розщеплення глюкози з утворенням в якості
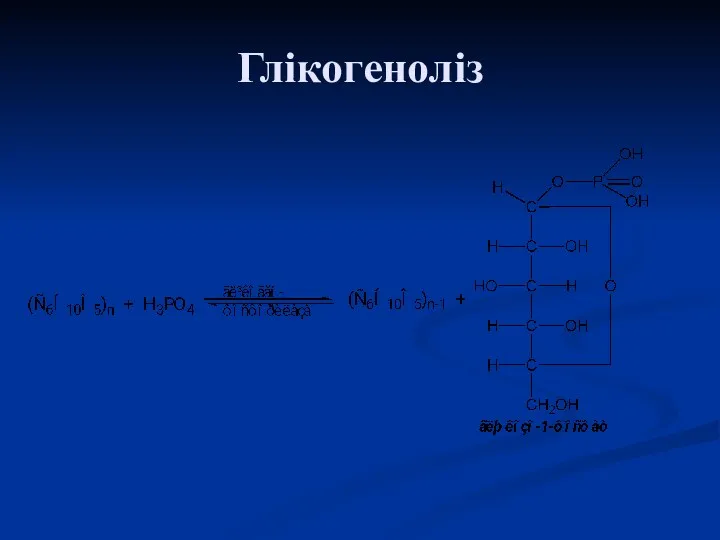
Глікогеноліз
Глікогеноліз
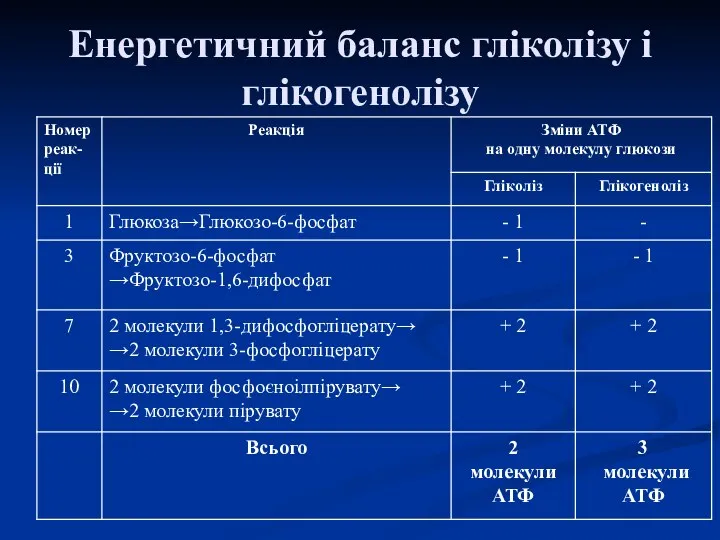
Енергетичний баланс гліколізу і глікогенолізу
Енергетичний баланс гліколізу і глікогенолізу

Катаболізм глюкози
Окиснення глюкози до СО2 і Н2О (аеробний розпад). Аеробний розпад
Катаболізм глюкози
Окиснення глюкози до СО2 і Н2О (аеробний розпад). Аеробний розпад

Етапи аеробного гліколізу
В аеробному гліколізі можна виділити 2 етапи.
1. Підготовчий етап,
Етапи аеробного гліколізу
В аеробному гліколізі можна виділити 2 етапи.
1. Підготовчий етап,

Центральним процесом катаболізму є
цикл лимонної кислоти
Головна функція циклу лимонної кислоти
Центральним процесом катаболізму є
цикл лимонної кислоти
Головна функція циклу лимонної кислоти

Ц.Т.К. – послідовні перетворення ди- і трикарбонових кислот, в ході яких
Ц.Т.К. – послідовні перетворення ди- і трикарбонових кислот, в ході яких
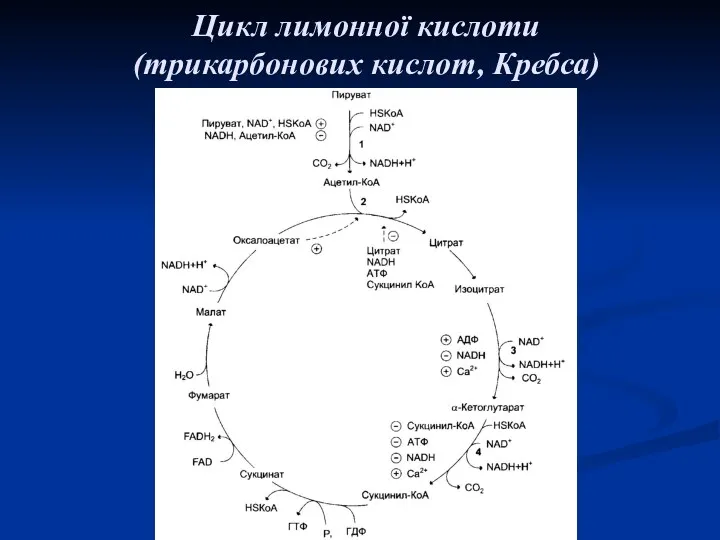
Цикл лимонної кислоти
(трикарбонових кислот, Кребса)
Цикл лимонної кислоти
(трикарбонових кислот, Кребса)
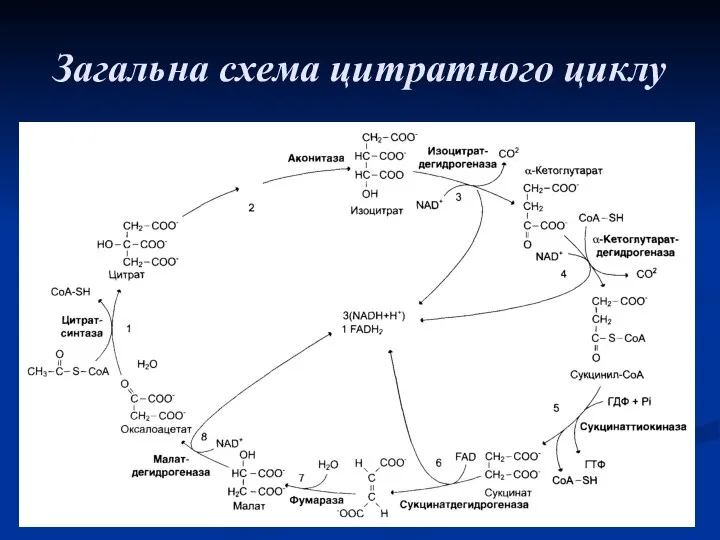
Загальна схема цитратного циклу
Загальна схема цитратного циклу

Цикл Кребса складається з 8 послідовних ферментативних реакцій:
Синтез лимонної кислоти або
Цикл Кребса складається з 8 послідовних ферментативних реакцій:
Синтез лимонної кислоти або
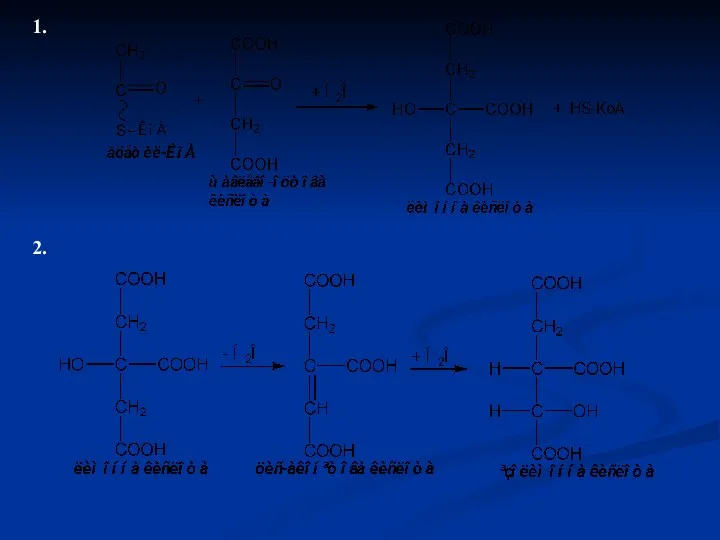
1.
2.
1.
2.

3.
ізолимонна кислота α-кетоглутарова кислота
4.
α-кетоглутарова
кислота
3.
ізолимонна кислота α-кетоглутарова кислота
4.
α-кетоглутарова
кислота
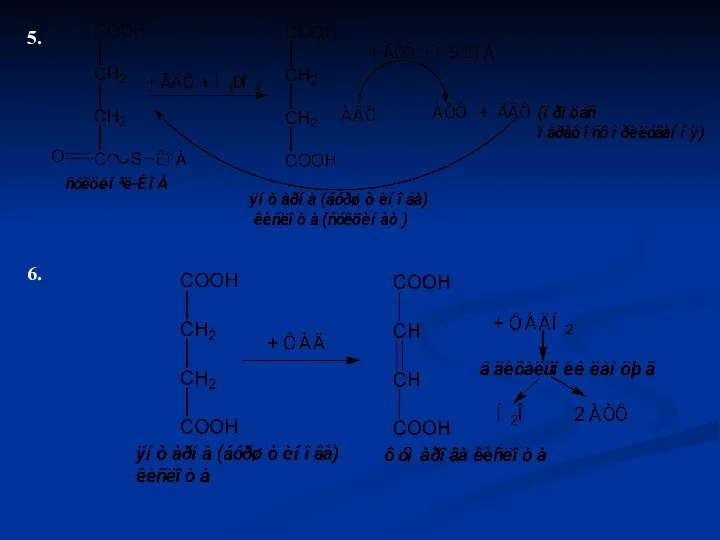
5.
6.
5.
6.
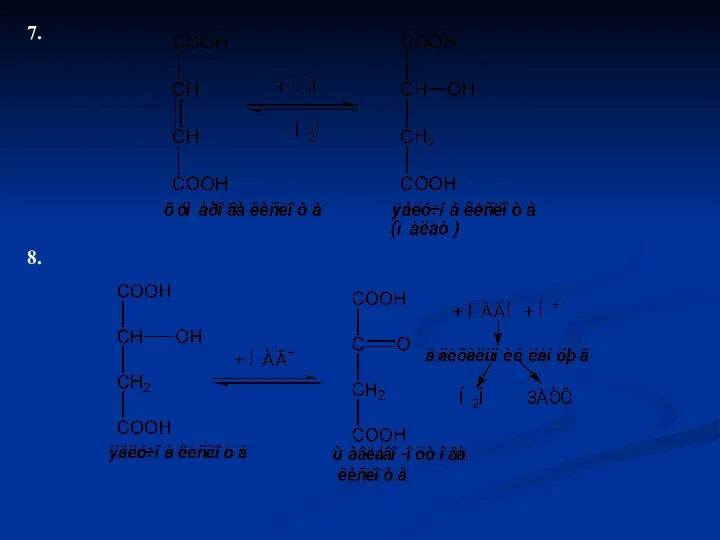
7.
8.
7.
8.

В результаті 1 витку циклу Кребса енергія виділяється у вигляді тепла

Регуляція ЦТК
Алостерична регуляція – здійснюється за рахунок 3 регуляторних ферментів:
цитратсинтази
Регуляція ЦТК
Алостерична регуляція – здійснюється за рахунок 3 регуляторних ферментів:
цитратсинтази

Біологічна роль ЦТК:
Інтегративна – він є метаболічним колектором, що об'єднує шляхи
Біологічна роль ЦТК:
Інтегративна – він є метаболічним колектором, що об'єднує шляхи

Енергетична ефективність
аеробного окиснення глюкози
1М глюкози 38 АТФ
в тому числі:
А)
Енергетична ефективність
аеробного окиснення глюкози
1М глюкози 38 АТФ
в тому числі:
А)

ПЕНТОЗОФОСФАТНИЙ ШЛЯХ ПЕРЕТВОРЕННЯ ГЛЮКОЗИ
Пентозофосфатний шлях, який називається також гексомонофосфатним шунтом, слугує
ПЕНТОЗОФОСФАТНИЙ ШЛЯХ ПЕРЕТВОРЕННЯ ГЛЮКОЗИ
Пентозофосфатний шлях, який називається також гексомонофосфатним шунтом, слугує

Пентозофосфатний шлях забезпечує клітини рибозою для синтезу пуринових та піримідинових нуклеотидів
Пентозофосфатний шлях забезпечує клітини рибозою для синтезу пуринових та піримідинових нуклеотидів

Окисний етап
В окисній частині пентозофосфатного шляху глюкозо-6-фосфат підлягає окисному декарбоксилуванню, в
Окисний етап
В окисній частині пентозофосфатного шляху глюкозо-6-фосфат підлягає окисному декарбоксилуванню, в
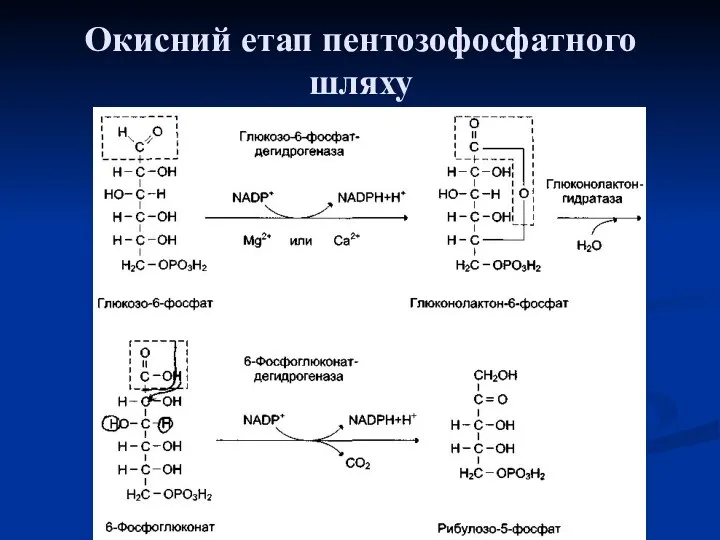
Окисний етап пентозофосфатного шляху
Окисний етап пентозофосфатного шляху

Реакції окисного етапу є основним джерелом НАДФH в клітинах. Гідровані коферменти
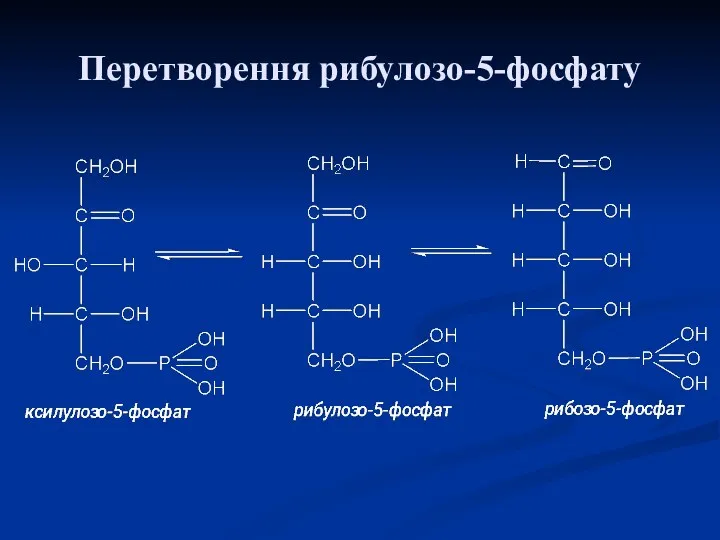
Перетворення рибулозо-5-фосфату
Перетворення рибулозо-5-фосфату

МЕТАБОЛІЗМ ГЛЮКУРОНОВОЇ КИСЛОТИ
Окрім гліколізу і пентозофосфатного шляху, що забезпечують засвоєння
МЕТАБОЛІЗМ ГЛЮКУРОНОВОЇ КИСЛОТИ
Окрім гліколізу і пентозофосфатного шляху, що забезпечують засвоєння

Схема синтезу вітаміну С
Схема синтезу вітаміну С

Біологічне значення глюкуронової кислоти проявляється в тому, що вона здатна зв'язуватися
Біологічне значення глюкуронової кислоти проявляється в тому, що вона здатна зв'язуватися

Синтез глікогену (глікогенез)
Синтез глікогену (глікогенез)
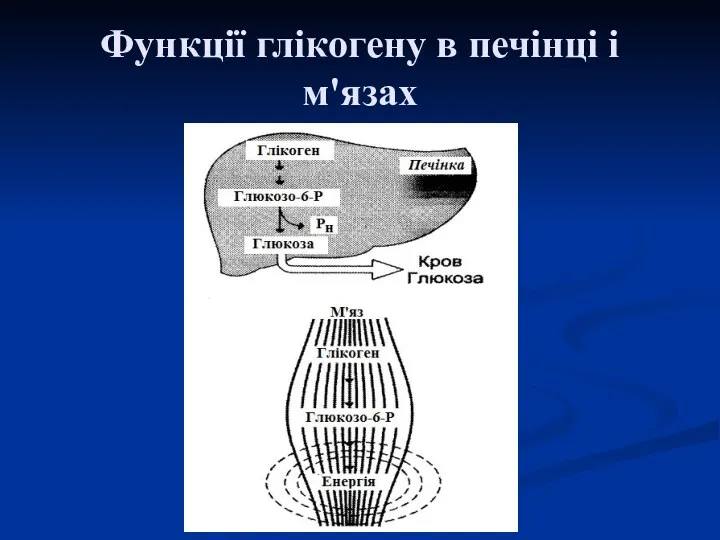
Функції глікогену в печінці і м'язах
Функції глікогену в печінці і м'язах

Регуляція глікогенезу
Активує цей процес інсулін.
Інгібують цей процес:
глюкагон,
адреналін.
Регуляція глікогенезу
Активує цей процес інсулін.
Інгібують цей процес:
глюкагон,
адреналін.

Глюконеогенез
це синтез глюкози з невуглеводних метаболітів (молочної кислоти, карбонових кислот, гліцерину,
Глюконеогенез
це синтез глюкози з невуглеводних метаболітів (молочної кислоти, карбонових кислот, гліцерину,
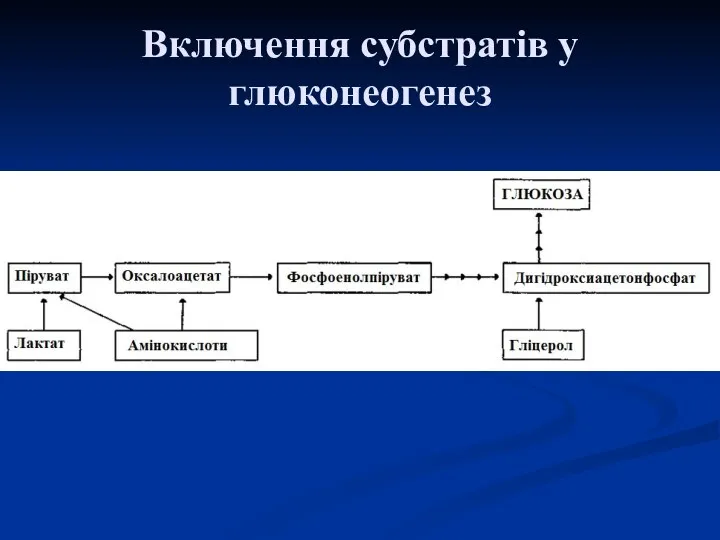
Включення субстратів у глюконеогенез
Включення субстратів у глюконеогенез
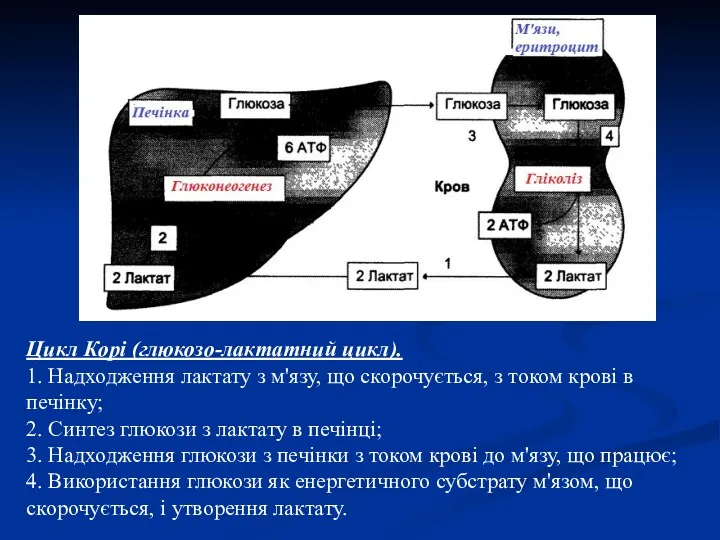
Цикл Корі (глюкозо-лактатний цикл).
1. Надходження лактату з м'язу, що скорочується, з
Цикл Корі (глюкозо-лактатний цикл).
1. Надходження лактату з м'язу, що скорочується, з

Лактат, що утворився в м'язах, які інтенсивно працюють або в клітинах
Лактат, що утворився в м'язах, які інтенсивно працюють або в клітинах

Цикл Корі виконує 2 важливі функції:
1 — забезпечує утилізацію лактату;
2
Цикл Корі виконує 2 важливі функції:
1 — забезпечує утилізацію лактату;
2

Синтез глюкози з амінокислот
В умовах голодування частина білків м'язової тканини розпадається
Синтез глюкози з амінокислот
В умовах голодування частина білків м'язової тканини розпадається

Синтез глюкози з гліцеролу
Гліцерол утворюється в результаті гідролізу триацил-гліцеролів, головним чином
Синтез глюкози з гліцеролу
Гліцерол утворюється в результаті гідролізу триацил-гліцеролів, головним чином

Гормони, регулятори рівня глюкози
а) гіпоглікемічної дії: інсулін
б) гіперглікемічної дії:
Основні: адреналін
Гормони, регулятори рівня глюкози
а) гіпоглікемічної дії: інсулін
б) гіперглікемічної дії:
Основні: адреналін

Регуляція рівня цукру в крові
Центр вуглеводного обміну гіпоталамусу
гіпофіз
АКТГ,ТТГ
Залози внутрішньої секреції
ГОРМОНИ
Регуляція рівня цукру в крові
Центр вуглеводного обміну гіпоталамусу
гіпофіз
АКТГ,ТТГ
Залози внутрішньої секреції
ГОРМОНИ

Біохімічні функції інсуліну
Це головний анаболічний гормон.
Підвищує проникність клітин печінки, нирок, жирової
Біохімічні функції інсуліну
Це головний анаболічний гормон.
Підвищує проникність клітин печінки, нирок, жирової

Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу):
глюкоза + АТФ
Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу):
глюкоза + АТФ
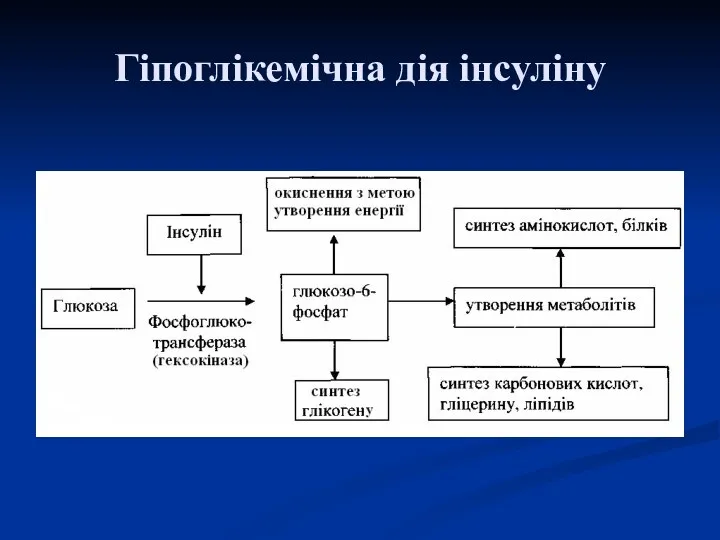
Гіпоглікемічна дія інсуліну
Гіпоглікемічна дія інсуліну

Гіпофункція по інсуліну
Захворювання цукровий діабет.
Основні його ознаки:
1) гіперглікемія;
2) глюкозурія;
3) кетонемія;
4) кетонурія;
5)
Гіпофункція по інсуліну
Захворювання цукровий діабет.
Основні його ознаки:
1) гіперглікемія;
2) глюкозурія;
3) кетонемія;
4) кетонурія;
5)
 Геохимические показатели техногенеза
Геохимические показатели техногенеза Реакция обменного разложения веществ водой - гидролиз
Реакция обменного разложения веществ водой - гидролиз Классы неорганических веществ
Классы неорганических веществ Коррозия металлов
Коррозия металлов Одноосновные карбоновые кислоты. Строение, физические свойства, номенклатура
Одноосновные карбоновые кислоты. Строение, физические свойства, номенклатура Химические понятия
Химические понятия Алкены (этиленовые углеводороды)
Алкены (этиленовые углеводороды) Хімія і здоров'я людини
Хімія і здоров'я людини Фазовые диаграммы и статистическая термодинамика бинарных m-h систем
Фазовые диаграммы и статистическая термодинамика бинарных m-h систем Цинк и его соединения
Цинк и его соединения Complex compounds
Complex compounds Ионное произведение воды. Водородный показатель
Ионное произведение воды. Водородный показатель Спирты
Спирты Щелочные металлы
Щелочные металлы Вода - уникальнейшая структура жизни. Способы ее очистки
Вода - уникальнейшая структура жизни. Способы ее очистки Химические свойства металлов
Химические свойства металлов Соединения железа
Соединения железа Неметаллы. Особенности строения атомов неметаллов
Неметаллы. Особенности строения атомов неметаллов Окислительно-восстановительные реакции
Окислительно-восстановительные реакции Новые катализаторы
Новые катализаторы Реакции ионного обмена
Реакции ионного обмена Полимерные материалы
Полимерные материалы Сполуки неметалічних елементів з Гідрогеном. Особливості водних розчинів цих сполук, їх застосування
Сполуки неметалічних елементів з Гідрогеном. Особливості водних розчинів цих сполук, їх застосування Твердость минералов и их применение
Твердость минералов и их применение Отрицательное влияние азотных минеральных удобрений на почву
Отрицательное влияние азотных минеральных удобрений на почву Теории химической кинетики. Лекция 4
Теории химической кинетики. Лекция 4 альдегиды, свойства, получение
альдегиды, свойства, получение Серная кислота H2SO4
Серная кислота H2SO4