- Цикли трикарбонових кислот

Содержание
- 2. Цикл ЦТК являє собою послідовність восьми реакцій, що протікають в матриксі мітохондрій. Схематично цей процес можна
- 4. Перша реакція циклу ЦТК - це необоротна реакція конденсації ацетил-КоА з оксалоацетатом , каталізується ферментом цитратсинтазою
- 5. Друга реакція це ізомеризація цитрата в ізоцитрат, у процесі якої відбувається перенесення гідроксигрупи до іншого атому
- 6. Третя реакція , подібно першiй – незворотня. У ній відбуваеться окиснювальне де карбоксилювання ізоцитрату: Гидроксигрупа ізоцитрату
- 7. Четверта реакцiя циклу-окиснювальне дикарбоксилювання а-кетоглутарата до високоенергiйного зєднання сукциніл -КоА. Механізм цієї реакції подібний з реакцією
- 8. П'ята реакція є єдиною в циклі реакцією субстратного фосфорилювання, каталізується ферментом сукциніл -КоА - синтетазой. У
- 9. Розщеплення шляхом фосфороліза тіоефірного зв'язку в сукцинил -КоА:
- 10. 2. Активована фосфорильна група сукцинілфосфата переноситься на ГДФ з утворенням ГТФ і сукцинату:
- 11. Сумарне рівняння для двох сполучених реакцій має наступний вигляд:
- 12. У шостій реакції відбувається дегідрирування сукцината до фумарату. Вона каталізується ферментом сукцинатдегідрогеназою, в молекулі якого з
- 13. У ході сьомої реакції здійснюється гідратація фумарату до L -малата. Вона каталізується стереоспецефічним ферментом фумаразою:
- 14. У восьмій, заключній реакції ЦТК відбувається регенерація оксалоацетата. Під дією Н АД + залежної малатдегідрогенази L-
- 15. Сумарне рівняння циклу трикарбонових кислот можна представити в наступному вигляді:
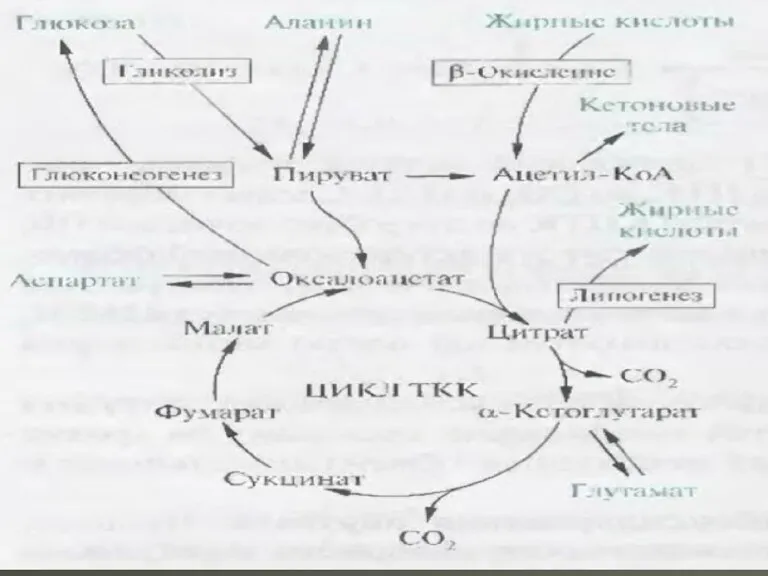
- 16. Далi наведена схема реакцій циклу трикарбонових кислот. Як видно зі схеми стехіометричного рівняння ЦТК , у
- 18. Таким чином, за один оборот циклу ТКК утворюється 12 молекул АТФ, з них 11 макроергів -
- 20. У клітинах печінки, серцевого м'яза і інших функціонує так звана малат - аспартатна човникова система перенесення
- 21. ВИСНОВОК Цикл трикарбонових кислот слід розглядати як універсальний механізм окислення ацетильной групи в аеробних умовах, оскільки
- 23. Скачать презентацию

Цикл ЦТК являє собою послідовність восьми реакцій, що протікають в матриксі
Цикл ЦТК являє собою послідовність восьми реакцій, що протікають в матриксі

Перша реакція циклу ЦТК - це необоротна реакція конденсації ацетил-КоА з
Перша реакція циклу ЦТК - це необоротна реакція конденсації ацетил-КоА з
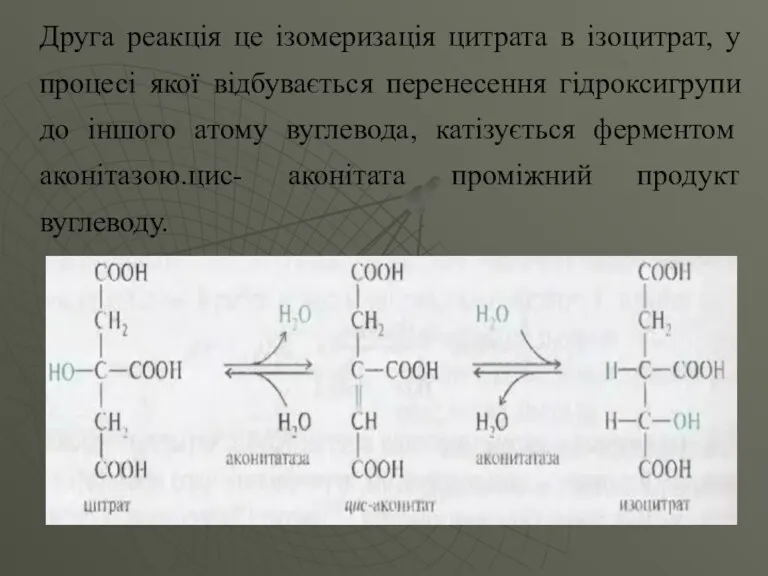
Друга реакція це ізомеризація цитрата в ізоцитрат, у процесі якої відбувається
Друга реакція це ізомеризація цитрата в ізоцитрат, у процесі якої відбувається

Третя реакція , подібно першiй – незворотня. У ній відбуваеться
Третя реакція , подібно першiй – незворотня. У ній відбуваеться

Четверта реакцiя циклу-окиснювальне дикарбоксилювання а-кетоглутарата до високоенергiйного зєднання сукциніл -КоА. Механізм
Четверта реакцiя циклу-окиснювальне дикарбоксилювання а-кетоглутарата до високоенергiйного зєднання сукциніл -КоА. Механізм

П'ята реакція є єдиною в циклі реакцією субстратного фосфорилювання, каталізується
П'ята реакція є єдиною в циклі реакцією субстратного фосфорилювання, каталізується

Розщеплення шляхом фосфороліза тіоефірного зв'язку в сукцинил -КоА:
Розщеплення шляхом фосфороліза тіоефірного зв'язку в сукцинил -КоА:
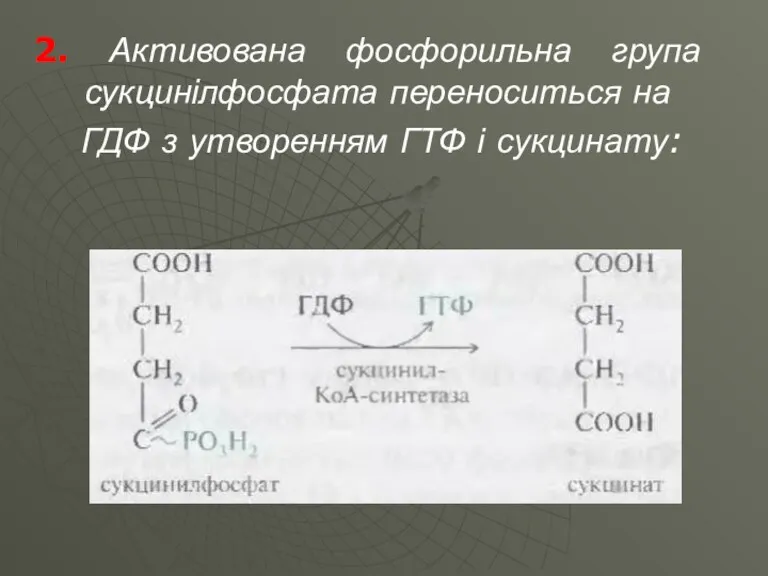
2. Активована фосфорильна група сукцинілфосфата переноситься на
ГДФ з утворенням ГТФ
2. Активована фосфорильна група сукцинілфосфата переноситься на
ГДФ з утворенням ГТФ

Сумарне рівняння для двох сполучених реакцій має наступний вигляд:
Сумарне рівняння для двох сполучених реакцій має наступний вигляд:

У шостій реакції відбувається дегідрирування сукцината до фумарату. Вона каталізується ферментом
У шостій реакції відбувається дегідрирування сукцината до фумарату. Вона каталізується ферментом
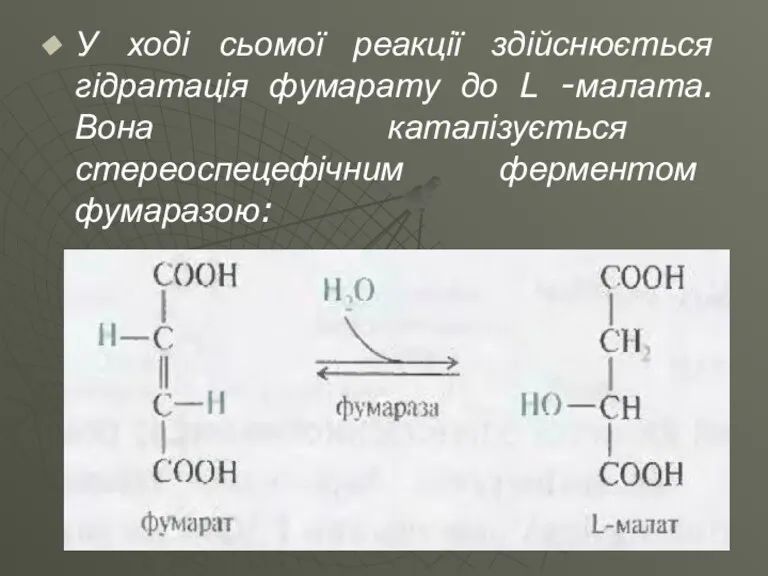
У ході сьомої реакції здійснюється гідратація фумарату до L -малата. Вона
У ході сьомої реакції здійснюється гідратація фумарату до L -малата. Вона

У восьмій, заключній реакції ЦТК відбувається регенерація оксалоацетата. Під дією Н
У восьмій, заключній реакції ЦТК відбувається регенерація оксалоацетата. Під дією Н

Сумарне рівняння циклу трикарбонових кислот можна представити в наступному вигляді:
Сумарне рівняння циклу трикарбонових кислот можна представити в наступному вигляді:

Далi наведена схема реакцій циклу трикарбонових кислот. Як видно зі схеми
Далi наведена схема реакцій циклу трикарбонових кислот. Як видно зі схеми
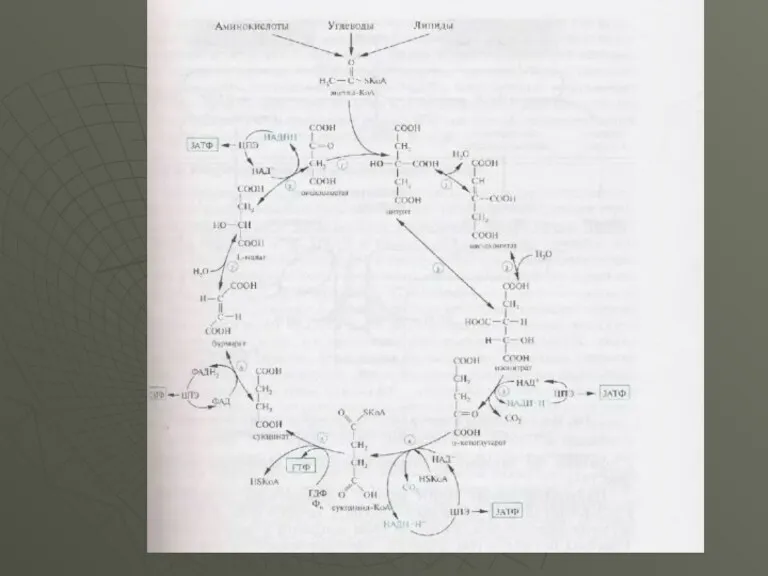

Таким чином, за один оборот циклу ТКК утворюється 12 молекул АТФ,
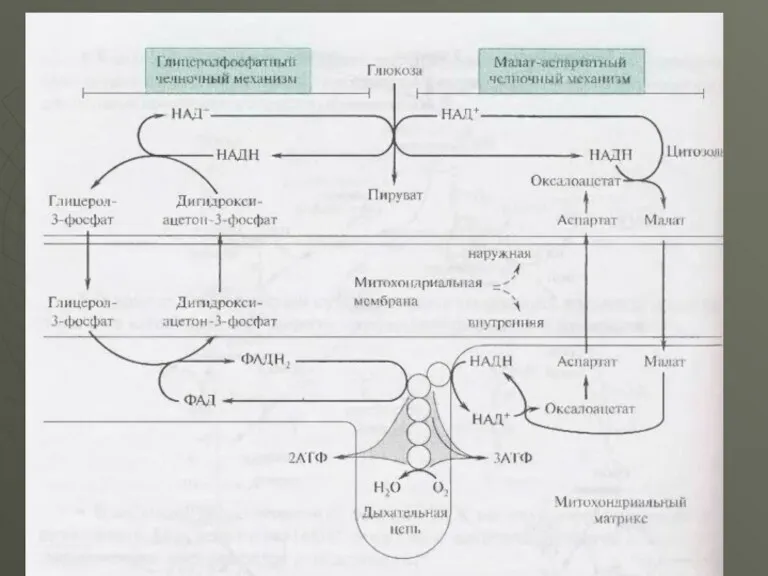

У клітинах печінки, серцевого м'яза і інших функціонує так звана малат
У клітинах печінки, серцевого м'яза і інших функціонує так звана малат

ВИСНОВОК
Цикл трикарбонових кислот слід розглядати як універсальний механізм окислення ацетильной групи
ВИСНОВОК
Цикл трикарбонових кислот слід розглядати як універсальний механізм окислення ацетильной групи
 Элемент V группы фосфор
Элемент V группы фосфор Первичная переработка нефти
Первичная переработка нефти Карбоновые кислоты. Изомерия. Физические, химические свойства. Получение, применение
Карбоновые кислоты. Изомерия. Физические, химические свойства. Получение, применение Железо
Железо Основные классы неорганических соединений
Основные классы неорганических соединений Нуклеопротеины
Нуклеопротеины Скорость химических реакций. Химическое равновесие
Скорость химических реакций. Химическое равновесие Оксиды неметаллов и кислородсодержащие кислоты. 11 класс
Оксиды неметаллов и кислородсодержащие кислоты. 11 класс Фазовые равновесия в системе ограниченно смешивающихся жидкостей
Фазовые равновесия в системе ограниченно смешивающихся жидкостей Реакции ионного обмена
Реакции ионного обмена Спирты. Физические и химические свойства
Спирты. Физические и химические свойства Алкалоиды, производные тропана
Алкалоиды, производные тропана Титриметрический метод анализа
Титриметрический метод анализа Figure
Figure Урок 25
Урок 25 Лекция 8.3. Реакции окисления и восстановления органических соединений
Лекция 8.3. Реакции окисления и восстановления органических соединений Халькогены. Кислород
Халькогены. Кислород Химические свойства кислот в свете теории электролитической диссоциации
Химические свойства кислот в свете теории электролитической диссоциации Формирование ключевых и предметных компетенций учащихся при изучении темы “Металлы”
Формирование ключевых и предметных компетенций учащихся при изучении темы “Металлы” Кристаллические решетки
Кристаллические решетки Создание слоев пористого кремния на p-Si
Создание слоев пористого кремния на p-Si Ауылшаруашылық дақылдарының тұқымдарын фунгецидтермен улау және химиялық қорғау тәсілдерінің биологиялық
Ауылшаруашылық дақылдарының тұқымдарын фунгецидтермен улау және химиялық қорғау тәсілдерінің биологиялық Тепловой эффект химических реакций. 8 класс
Тепловой эффект химических реакций. 8 класс Химическая связь
Химическая связь Chemistry th 11 grade The end of second Term
Chemistry th 11 grade The end of second Term Методика изучения современной теории строения как фундамента курса органической химии в старшей школе
Методика изучения современной теории строения как фундамента курса органической химии в старшей школе Производные пиримидина
Производные пиримидина Синтез олиголактозаминов – лигандов галектинов
Синтез олиголактозаминов – лигандов галектинов