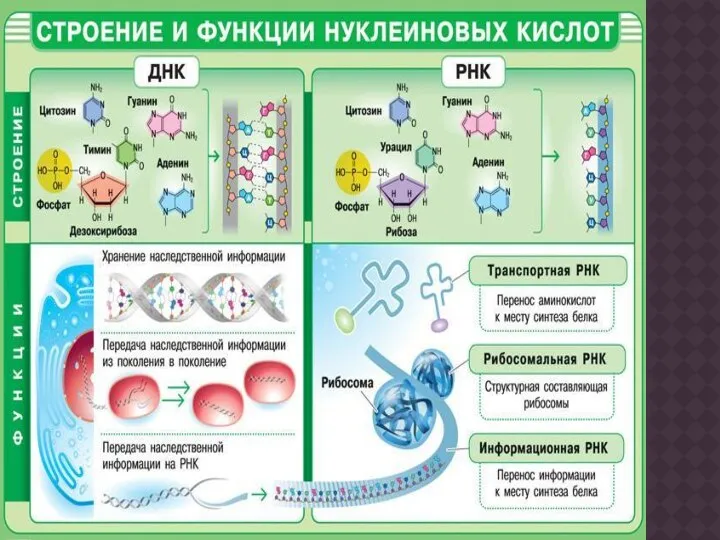
- Нуклеиновые кислоты, их структурные компоненты

Содержание
- 3. Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
- 4. НУКЛЕИНОВЫЕ ОСНОВАНИЯ Пиримидиновые основания в составе полинуклеотидов представлены лактамной формой, что обусловлено возможностью образования водородных связей
- 5. Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти гетероциклические соединения способны к лактим-лактамной таутомерии и
- 6. РЕДКИЕ (МИНОРНЫЕ) КОМПОНЕНТЫ НУКЛЕИНОВЫХ КИСЛОТ В ДНК встречаются метилированные основания: 5-метилцитозин, 6-N- метиладенин и др. В
- 7. НУКЛЕОЗИДЫ Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой.. D-рибоза и 2-дезокси-D-рибоза в состав природных
- 8. СТРУКТУРА НУКЛЕОЗИДОВ
- 9. В состав некоторых РНК входят необычные нуклеозиды. Например, инозин, который можно рассматривать как продукт дезаминирования аденозина,
- 10. НУКЛЕОТИДЫ Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил при С-5' или С-3' в
- 11. СТРУКТУРА НЕКОТОРЫХ НУКЛЕОТИДОВ
- 12. Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества энергии: 30 кДж/моль и более.
- 13. СТРОЕНИЕ ПОЛИНУКЛЕОТИДОВ Многообразие молекул ДНК и РНК определяется их первичной структурой – последовательностью нуклеотидных остатков в
- 14. ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ Первичная структура нуклеиновых кислот – нуклеотидный состав и нуклеотидная последовательность, т.е. порядок
- 15. Нуклеотидный состав ДНК различного происхождения находится в соответствии с правилами Э.Чаргаффа: 1) Все ДНК независимо от
- 16. ВТОРИЧНАЯ СТРУКТУРА ДНК Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи. Водородные связи образуются между
- 17. В сдвоенной молекуле ДНК суммарно число групп А равно числу групп Т, а число групп G
- 18. ВТОРИЧНАЯ СТРУКТУРА РНК Молекула РНК построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные
- 19. Основная роль РНК - непосредственное участие в биосинтезе белка. Известны три вида клеточных РНК, которые отличаются
- 20. ТРЕТИЧНАЯ СТРУКТУРА ДНК Третичная структура ДНК эукариот формируется путем взаимодействия с ядерными белками и на определенном
- 21. ХИМИЧЕСКИЕ СВОЙСТВА НУКЛЕИНОВЫХ КИСЛОТ ДНК и РНК имеют много общих химических и физических свойств. Так, нуклеиновые
- 22. Гидролиз в щелочной среде В щелочной среде РНК легко гидролизуются до нуклеотидов, которые в свою очередь,
- 24. Скачать презентацию
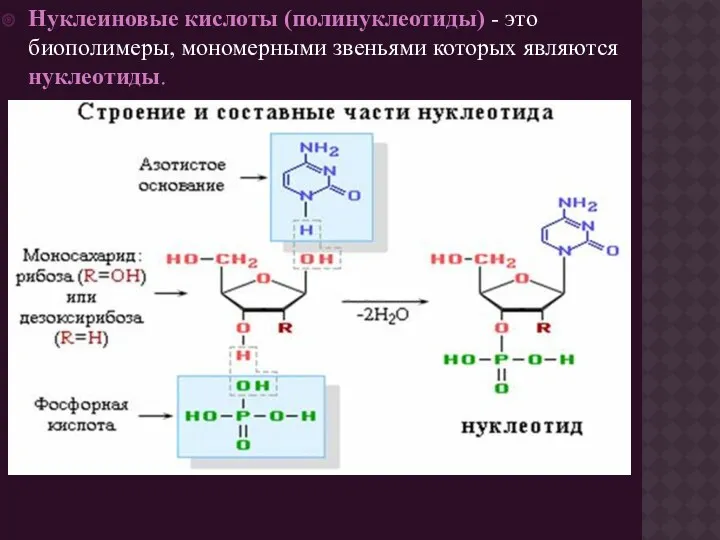
Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
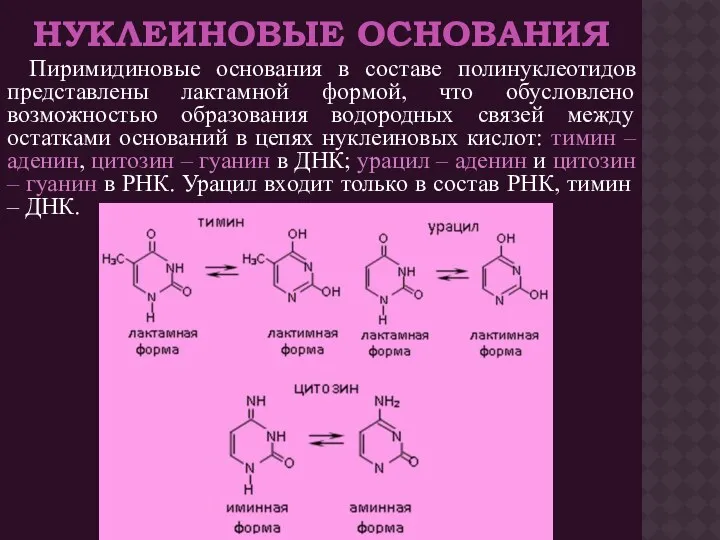
НУКЛЕИНОВЫЕ ОСНОВАНИЯ
Пиримидиновые основания в составе полинуклеотидов представлены лактамной формой, что
НУКЛЕИНОВЫЕ ОСНОВАНИЯ
Пиримидиновые основания в составе полинуклеотидов представлены лактамной формой, что
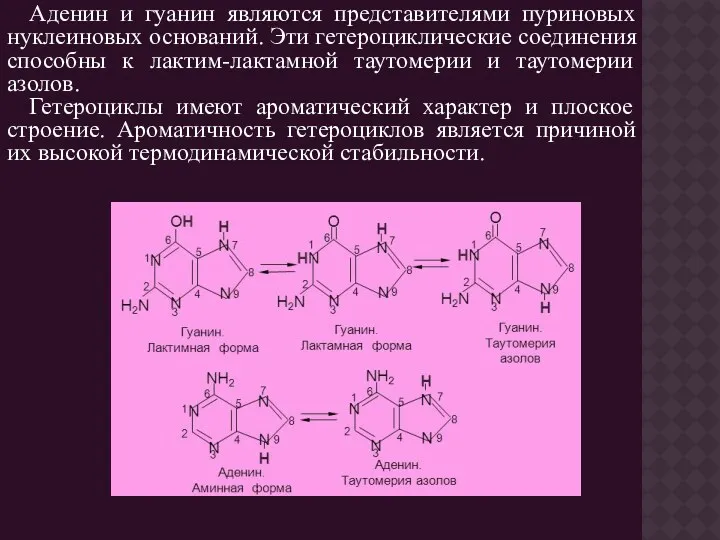
Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти гетероциклические соединения
Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти гетероциклические соединения

РЕДКИЕ (МИНОРНЫЕ) КОМПОНЕНТЫ НУКЛЕИНОВЫХ КИСЛОТ
В ДНК встречаются
метилированные
основания: 5-метилцитозин,
6-N-
РЕДКИЕ (МИНОРНЫЕ) КОМПОНЕНТЫ НУКЛЕИНОВЫХ КИСЛОТ
В ДНК встречаются
метилированные
основания: 5-метилцитозин,
6-N-

НУКЛЕОЗИДЫ
Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой..
D-рибоза и
НУКЛЕОЗИДЫ
Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой..
D-рибоза и

СТРУКТУРА НУКЛЕОЗИДОВ
СТРУКТУРА НУКЛЕОЗИДОВ
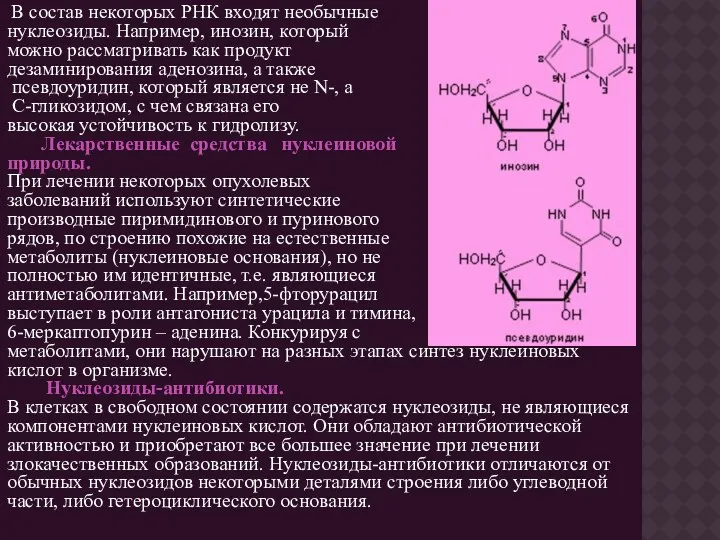
В состав некоторых РНК входят необычные
нуклеозиды. Например, инозин, который
В состав некоторых РНК входят необычные
нуклеозиды. Например, инозин, который

НУКЛЕОТИДЫ
Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил
НУКЛЕОТИДЫ
Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил
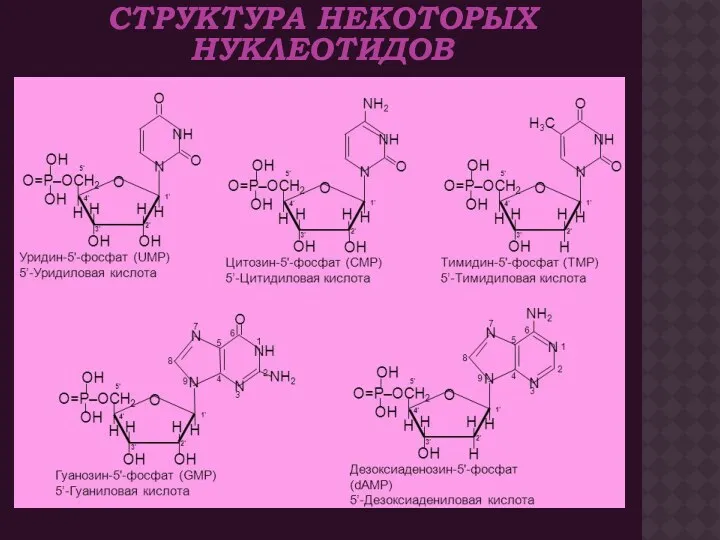
СТРУКТУРА НЕКОТОРЫХ НУКЛЕОТИДОВ
СТРУКТУРА НЕКОТОРЫХ НУКЛЕОТИДОВ
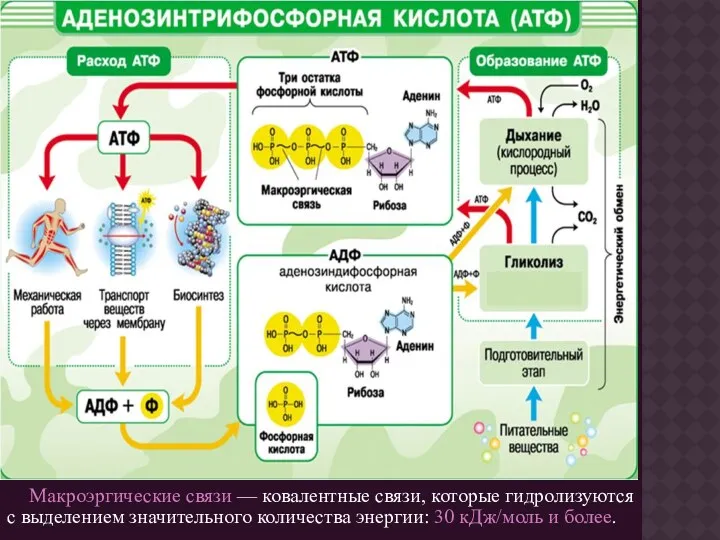
Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества
Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества
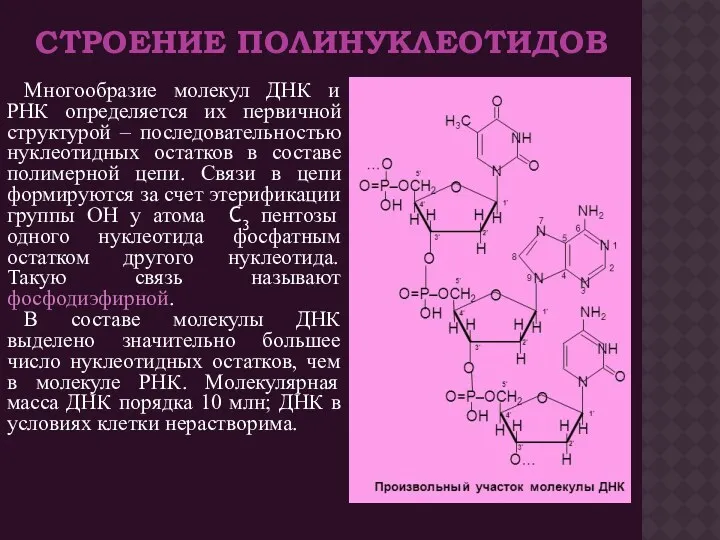
СТРОЕНИЕ ПОЛИНУКЛЕОТИДОВ
Многообразие молекул ДНК и РНК определяется их первичной структурой
СТРОЕНИЕ ПОЛИНУКЛЕОТИДОВ
Многообразие молекул ДНК и РНК определяется их первичной структурой
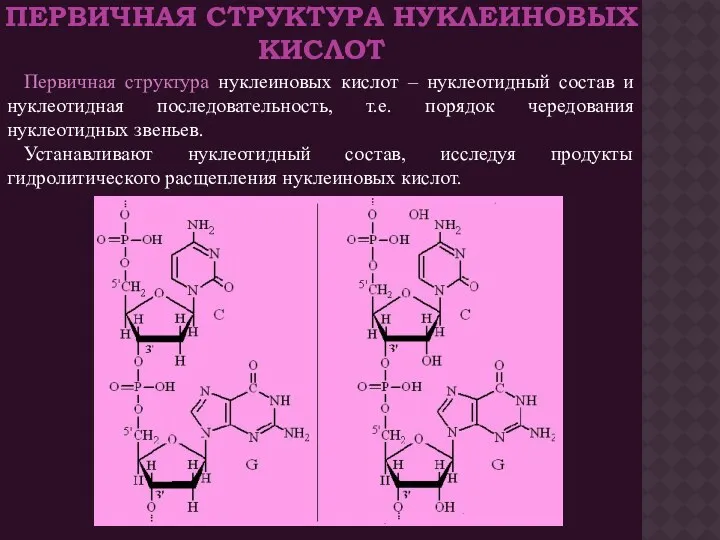
ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Первичная структура нуклеиновых кислот – нуклеотидный состав и
ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Первичная структура нуклеиновых кислот – нуклеотидный состав и

Нуклеотидный состав ДНК различного происхождения находится в соответствии с правилами Э.Чаргаффа:
1)
Нуклеотидный состав ДНК различного происхождения находится в соответствии с правилами Э.Чаргаффа:
1)
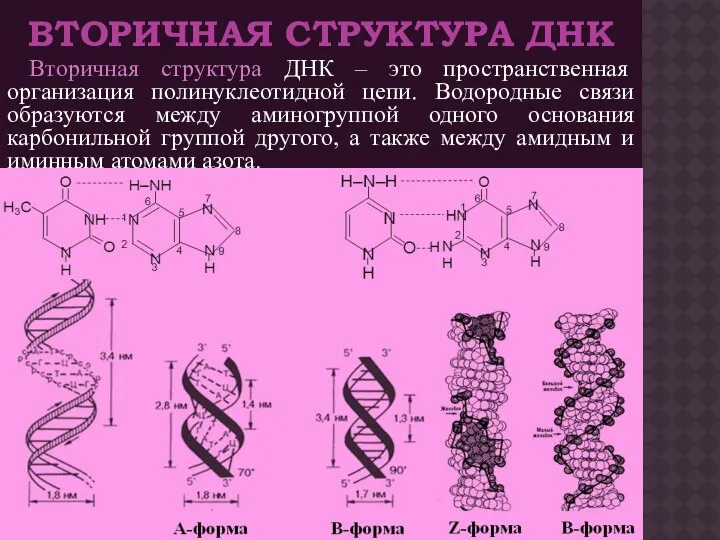
ВТОРИЧНАЯ СТРУКТУРА ДНК
Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи.
ВТОРИЧНАЯ СТРУКТУРА ДНК
Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи.

В сдвоенной молекуле ДНК суммарно число групп А равно числу групп
В сдвоенной молекуле ДНК суммарно число групп А равно числу групп

ВТОРИЧНАЯ СТРУКТУРА РНК
Молекула РНК построена из одной
полинуклеотидной цепи. Отдельные
участки
ВТОРИЧНАЯ СТРУКТУРА РНК
Молекула РНК построена из одной
полинуклеотидной цепи. Отдельные
участки

Основная роль РНК - непосредственное участие в биосинтезе белка. Известны три
Основная роль РНК - непосредственное участие в биосинтезе белка. Известны три
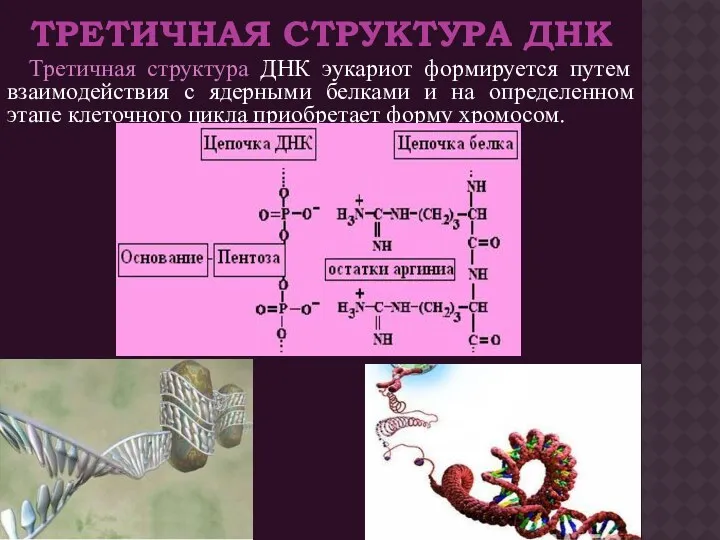
ТРЕТИЧНАЯ СТРУКТУРА ДНК
Третичная структура ДНК эукариот формируется путем взаимодействия с ядерными
ТРЕТИЧНАЯ СТРУКТУРА ДНК
Третичная структура ДНК эукариот формируется путем взаимодействия с ядерными

ХИМИЧЕСКИЕ СВОЙСТВА НУКЛЕИНОВЫХ КИСЛОТ
ДНК и РНК имеют много общих химических и
ХИМИЧЕСКИЕ СВОЙСТВА НУКЛЕИНОВЫХ КИСЛОТ
ДНК и РНК имеют много общих химических и

Гидролиз в щелочной среде
В щелочной среде РНК легко гидролизуются до нуклеотидов,
Гидролиз в щелочной среде
В щелочной среде РНК легко гидролизуются до нуклеотидов,
 История развития и предмет исследования супрамолекулярной химии. Классификация супрамолекулярных соединений
История развития и предмет исследования супрамолекулярной химии. Классификация супрамолекулярных соединений Химия в решении сырьевой проблемы
Химия в решении сырьевой проблемы Основные химические понятия и законы
Основные химические понятия и законы Энергетика химических реакций.(Лекция 8,9)
Энергетика химических реакций.(Лекция 8,9) Немного формул. Aminoacids
Немного формул. Aminoacids Отчет по исследовательской работе Образование АСПО
Отчет по исследовательской работе Образование АСПО Крохмаль. Структурна формула крохмалю
Крохмаль. Структурна формула крохмалю 20230330_metodicheskaya_razrabotka_ovr_v_organicheskoy_himii
20230330_metodicheskaya_razrabotka_ovr_v_organicheskoy_himii Химияның негізгі стехиометриялық заңдары
Химияның негізгі стехиометриялық заңдары Щелочноземельные металлы
Щелочноземельные металлы Кремний. Физические свойства кремния
Кремний. Физические свойства кремния Спирты. Классификация
Спирты. Классификация Методика изучения современной теории строения органических веществ
Методика изучения современной теории строения органических веществ Бериллий, магний и щелочноземельные металлы. Главная подгруппа II группы
Бериллий, магний и щелочноземельные металлы. Главная подгруппа II группы химиияяяяяяяяяяяяяяяяяяяяяяяяяяяяяяяяя
химиияяяяяяяяяяяяяяяяяяяяяяяяяяяяяяяяя Подготовка к ГИА. А2. Периодический закон и Периодическая система химических элементов Д.И. Менделеева
Подготовка к ГИА. А2. Периодический закон и Периодическая система химических элементов Д.И. Менделеева Бутадиеновые каучуки
Бутадиеновые каучуки Совместимость электродных материалов в новой электрохимической системе Li4Ti5O12/Li3V2(PO4)3 с традиционным электролитом для
Совместимость электродных материалов в новой электрохимической системе Li4Ti5O12/Li3V2(PO4)3 с традиционным электролитом для Вода: фізичні та хімічні властивості. Поширеність в природі
Вода: фізичні та хімічні властивості. Поширеність в природі Кислоты: названия, получение, свойства, применение
Кислоты: названия, получение, свойства, применение Оксиды в минералогии
Оксиды в минералогии Аналитическая химия. Физико-химические методы анализа
Аналитическая химия. Физико-химические методы анализа Химические превращения веществ
Химические превращения веществ Кремний и его соединения. Нахождение в природе
Кремний и его соединения. Нахождение в природе Международный Год периодической таблицы химических элементов
Международный Год периодической таблицы химических элементов Anionic Polymerization
Anionic Polymerization Щелочи
Щелочи Оксиды неметаллов и кислородсодержащие кислоты. 11 класс
Оксиды неметаллов и кислородсодержащие кислоты. 11 класс